
Abstract
Evidence of human impact on the vegetation obtained from pollen diagrams at sites in the process of neolithisation is often difficult to detect. Apart from aspects like site function and occupation intensity, methodological aspects play a considerable role. In the Rhine-Meuse delta in the Netherlands, neolithisation is documented at the Final Mesolithic sites Hardinxveld-Giessendam Polderweg and Hardinxveld-Giessendam De Bruin and the local Early Neolithic sites of Brandwijk-Kerkhof and the Hazendonk, covering the period of 5500–2500 cal. b.c. The off-site core from Hardinxveld-Giessendam De Bruin supports earlier results that human influence on the vegetation at Hardinxveld is restricted and difficult to distinguish from natural vegetation disturbance. Human impact is more easily recognisable in the diagrams of Neolithic phases at Brandwijk-Kerkhof and the Hazendonk that show evidence of both domestic animals and cereals. Continuing neolithisation and also research methodology, including the location of the pollen cores, may play a role in this. It is tested whether the use of a new pollen sum excluding extra-regional taxa increases the visibility of human impact.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Neolithisation (the process of the onset of Neolithic culture) can be studied by analysing human impact on the vegetation as registered in pollen diagrams. In these, neolithisation is often characterised by a decrease of arboreal pollen, an increase of pioneers and/or secondary vegetation and an increase of anthropogenic indicators. These include amongst others taxa that depend on people for their distribution and persist only by direct or indirect human influence on the environment (anthropochores), taxa that were originally part of the natural vegetation and became arable weeds after the introduction of crop cultivation (apophytes), and microcharcoal. Together these changes point to the creation of open patches, trampling, increased disturbance and eutrophication, resulting from increasing sedentism, clearance of woodland and agricultural activities (Behre 1981; Fægri and Iversen 1989, p. 179; Kalis and Meurers-Balke 1998; Moore et al. 1991, p. 185). Non-pollen palynomorphs (NPPs) may additionally indicate aspects such as the degree of eutrophy or the presence of decomposed plant material (cellulose, wood and/or herbivore dung), redeposited sand or carbonised plant material (Innes et al. 2006; Van Geel and Aptroot 2006; Van Geel et al. 2003).
Human impact before the introduction of agriculture is often difficult to gauge due to the marked human mobility and modest densities of Palaeolithic and Mesolithic people (Bos and Urz 2003; Bos et al. 2005; Day and Mellars 1998, pp. 112, 209; Hörnberg et al. 2006; Regnell 2012). Even after the introduction of agriculture, the detection and understanding of human impact in pollen diagrams remains a challenge, depending on the role and scale of agriculture, human mobility, human population densities, site function and seasonality. Furthermore, small-scaled, local disturbances of the vegetation would have created only small woodland clearings from which pollen would have dispersed only on a restricted scale, and the surrounding vegetation may have masked the signals of human activity (Binney et al. 2005; Bunting et al. 2005; Birks and Birks 1980, p. 179; Brown 1999; Fægri and Iversen 1989, p. 147; Jackson 1994; Sugita 1994; Waller et al. 2005). Therefore, human impact at Palaeolithic, Mesolithic and Early Neolithic sites can best be studied by high-resolution studies and the analysis of multiple cores including core transects and on- and off-site cores (Behre and Kučan 1986; Bos and Janssen 1996; Bos and Urz 2003; Joly and Visset 2009; Kuneš et al. 2007; Out 2008a, b; Poska et al. 2004).
An aspect also affecting the detection of human impact is the pollen sum, of which the composition depends on the vegetation and the research question (Birks and Birks 1980, p. 167; Jackson 1994). In order to focus on regional developments or human impact, taxa often omitted from the sum are local taxa (Janssen 1970, p. 195; Moore et al. 1991, p. 171) such as aquatics or wetland taxa (Waller et al. 2005, p. 200; Van der Wiel 1982), or local, overrepresented trees (Alnus or Betula) and dry land herbs (Janssen 1959; Van Zeist 1967). It can be argued that for an optimal understanding of extra-local and regional human impact on the vegetation, extra-regional taxa should also be excluded from the pollen sum (cf. Fægri and Iversen 1989, p. 142; Janssen 1970, p. 190). Such exclusion of extra-regional taxa is only acceptable when clear indications exist of their very limited presence in the local, extra-local and if possible regional vegetation.
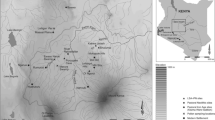
The case studies presented here investigate human impact during neolithisation at inland river dune sites in the Rhine-Meuse delta in the Netherlands (Fig. 1), with focus on an off-site core from Hardinxveld-Giessendam De Bruin. The wetland sites in the Rhine-Meuse delta illustrate the neolithisation process in the Netherlands after the arrival of the Linear Pottery Culture (LBK) in the southern loess area at 5300 cal. b.c. (Raemaekers 1999; Louwe Kooijmans 2007; cal b.c. will referred to as b.c. from now on). The Final Mesolithic sites of Hardinxveld-Giessendam Polderweg and De Bruin record the introduction of Swifterbant pottery at 4900 b.c. and the earliest known presence of domestic animals at 4700–4500 b.c. (Louwe Kooijmans 2003, 2007; though see Rowley-Conwy 2013). At Brandwijk-Kerkhof, two minor ceramic phases with few remains of domestic animals and no evidence of crops between 4600 and 4370 b.c. are followed by ceramic phases with evidence of domestic animals and cereals between 4220 and 3820 b.c. (Raemaekers 1999; Out 2008a, 2009, App. 2). At the Hazendonk, repetitively occupied between 4000 and 2500 b.c., all main phases yielded evidence of pottery, domestic animals and cereal grains (Raemaekers 1999; Zeiler 1997; Out 2010a). All sites show evidence of contacts with or influence from societies in the southern Pleistocene sand area of the Netherlands (Louwe Kooijmans 2007; Vanmontfort 2008). Insight in the relationship with this region is however limited due to the poor preservation of organic remains in the Pleistocene sandy soils. Other contemporary sites in the river area have been attested Guiran and Brinkkemper 2007; De Koning and Van Dasselaar 2008; Schiltmans 2010; Van der Linden 2010; Verbruggen 1992; Zijl et al. 2011), but excavated sites covering the Mesolithic/Neolithic transition with well-dated, extensive botanical data sets are rare.

Palaeogeographical map of The Netherlands at 5500 cal. b.c. (Vos et al. 2011). 1 Hardinxveld-Giessendam Polderweg; 2 Hardinxveld-Giessendam De Bruin; 3 the Hazendonk; 4 Brandwijk-Kerkhof
This paper reconstructs the development of the vegetation and human impact as reflected by the off-site pollen diagram from Hardinxveld-Giessendam De Bruin, focussing on the period corresponding with the occupation and submergence of the site. The results of this off-site core are compared with earlier published diagrams of Hardinxveld-Giessendam Polderweg, Hardinxveld-Giessendam De Bruin, Brandwijk-Kerkhof and the Hazendonk to evaluate the evidence for human impact during neolithisation. Additionally, it is considered whether the exclusion of extra-regional taxa from the pollen sum provides clearer signals of human impact in the area of the Rhine-Meuse delta. This is investigated by comparing the main diagrams of the mentioned sites based on the traditional pollen sum with those based on a modified pollen sum.
Hardinxveld-Giessendam Polderweg and De Bruin
Archaeological background and palaeogeography
The dune sites of Polderweg and De Bruin, located 1 km from each other, revealed colluviums, refuse layers, hearths, postholes, pits and various types of organic remains. The sites were repetitively occupied between 5500 and 4450 b.c., Polderweg being inhabited mainly between 5500 and 5000 b.c. and De Bruin mainly afterwards. At both sites, three occupation phases were distinguished (Table 1). The subsistence was based on a broad-spectrum economy combining gathering, fishing, fowling and hunting, with wild boar, beaver, otter and pike as primary animal resources. Domestic animals other than dogs are represented only in the latest phase of De Bruin by limbs of cow and sheep/goat and teeth of pigs that, together with other parts of pig/wild boar, may represent complete pigs. An active search for cereals at De Bruin did not provide evidence of crop plants. Polderweg is interpreted as a winter base camp for households, while De Bruin is interpreted as such a camp changing into a seasonal extraction camp (Louwe Kooijmans 2001a, b, 2003).
The Hardinxveld sites are situated on inland dunes which were deposited during the Younger Dryas and Pre-boreal (Fig. 2). The eustatic rise of the sea level caused marine and fluvial aggradation with deposition of peat and clay; see Van de Plassche et al. (2010) for a local ground water table curve. At both sites, refuse layers representing subsequent occupation phases wedge out into the surrounding sediments at different levels. During occupation, an anastomosing river system was active in the region. The landscape in the central river area was dominated by shallow eutrophic lakes, alder carr and river channels (Hijma and Cohen 2011; Mol 2003; Van der Woude 1983). Near Polderweg and De Bruin, a crevasse channel was active at c. 5500–5100 b.c. and c. 4700–4450 b.c., allowing the deposition of loam and clay near De Bruin during the period 4700–4450 b.c. At the onset of occupation, Polderweg and De Bruin had surface areas of c. 4500 and 44,200 m2 respectively, both rising c. 4 m above the surrounding landscape. Two other dunes existed nearby until they were submerged at 5000 b.c.; Polderweg followed soon afterwards. During the last occupation phase, the surface of De Bruin, restricted to two islands, was c. 950 m2 and protruding c. 0.8 m (Fig. 2). The dunes are covered by the clayey deposits of the anastomosing river Schaik system (Mol 2001a, b, 2003).

Earlier palynological studies at Hardinxveld-Giessendam
At Polderweg, the pollen investigation was based on a core located north of the site, partially corresponding to all main occupation phases (Fig. 2b; Bakels and Van Beurden 2001). The distance between the core and the refuse layer, located at the other side of the dune and representing the main area of human activity, was c. 40 m during all phases. The distance to the dune edge was 4 m during Phase 1, 15 m during Phase 1/2 and 26 m during Phase 2. The published analysis of De Bruin (ESM 1; Bakels et al. 2001) concerns two sample series from the excavation trench, covering particularly the two later occupation phases. The distance to the refuse layer was less than 4 m for all phases. The distance to the dune edge was 0 m during Phase 1 and a maximum of 10 m during Phase 3 (Fig. 2c). At both sites, the choice for sample locations at both sites was restricted by the frequent occurrence of wood in the subsoil.
Besides palynomorphs, the botanical analyses at Polderweg and De Bruin covered macrofossils, wood and charcoal from the excavation trenches (Bakels and Van Beurden 2001; Bakels et al. 2001; Louwe Kooijmans et al. 2001a, b). The integrated approach allowed a detailed reconstruction of the vegetation. At both sites, the dry land vegetation consisted of deciduous lime-oak woodland changing down-slope and with time into alder carr and vegetation of marshes and open water (Bakels and Van Beurden 2001; Bakels et al. 2001). A more detailed description of the vegetation is given in the Discussion.
The available evidence of human impact on the vegetation at Hardinxveld is limited. The AP/NAP ratio is quite stable, thus not providing evidence of large-scale woodland clearance, and it is not possible to recognise individual occupation phases in the pollen diagrams. Anthropogenic indicators such as Chenopodiaceae, Artemisia and Urtica dioica are few, do not include crop plants or show major changes through time. The secondary shrub vegetation with Viburnum opulus, Rhamnus cathartica, Cornus sanguinea-type and Sambucus nigra-type shows a gradual increase through time, perhaps related to human impact (Bakels et al. 2001, p. 379). While this may have been the case, the changes may alternatively have been caused by non-human factors such as tree falls (Louwe Kooijmans 2001a, p. 89), animal activity (foraging by beavers or deer) or the gradual rising water table. Only at De Bruin, Phase 2 corresponds with a decrease of Quercus, a peak of Corylus, Alnus fluctuations, peaks of the wetland taxa Poaceae, Humulus lupulus, Urtica dioica-type, Lythrum salicaria and a small Chenopodiaceae peak (at −7.40 m NAP; NAP = Dutch Ordnance Datum). These changes indicate the increased presence of light, nutrients and disturbance and are probably related to local human activity (cf. Bakels et al. 2001, p. 378). Moreover, a concentration of Acipenser sturio (sturgeon) remains, probably fish cleaning waste, was found near the sample location (Beerenhout 2001, p. 316). A structure interpreted as a landing place for canoes was located only a few metres away. It therefore appears that these changes during Phase 2 do indeed reflect human impact (Bakels et al. 2001, p. 378; Out 2009, App. 1, p. 34).
Materials and methods
Analysis of the off-site core Hardinxveld-Giessendam De Bruin
The off-site core at De Bruin was sampled south of the dune away from the excavation trench and the refuse layers (coordinates: N51°50′25.7″, E4°48′33.2″). The sampling location was chosen for its proximity to the dune, the presence of peat that could be linked stratigraphically to the site’s occupation, and the absence of wood remains that prevented sampling at most alternative locations. The distance to the dune was 0, 30 and 35 m during Phases 1, 2 and 3 respectively, while the distance to the crevasse channel during Phase 3 was c. 60 m (Fig. 2c). The sediment of the core consisted of amorphous peat and detritus-gyttja, in its lower part mixed with dune sand and higher up mixed with clay deposited during the Middle Ages. The uppermost part of the core, consisting of clay, was not investigated. The sample interval was 20 cm in the lower half of the core and 40 cm in the upper half.
After short boiling with 10 % KOH to decay humic acids, the samples with a volume of 1 cm3 were sieved on a 1 mm mesh to retrieve macrofossils. The standard pollen analysis method according to Fægri and Iversen (1989) was followed. Acetolysis with 10 % sulphuric acid (H2SO4) and 90 % acetic anhydride (C4H6O3) was applied to dissolve cellulose. Specific gravity separation with bromoform/alcohol (ρ = 2.0 g/ml) permitted separation of the palynomorphs from the sediment. One Lycopodium tablet was added to each sample. In order to study human impact on the dry land vegetation, and to make the diagram comparable with the earlier Hardinxveld diagrams, the analysis was based on an upland pollen sum of a minimum of 300 pollen grains (dry land trees, shrubs, herbs, spore plants and crop plants). Verhoeven (2003) analysed the pollen and NPPs and W.J. Kuijper identified the macrofossils. Pollen identification was based on literature (Fægri and Iversen 1989; Moore et al. 1991; Punt et al. 1976–1995; Reille 1992) and the reference collection of the Faculty of Archaeology, Leiden University, using the nomenclature of Fægri and Iversen (1989). The identification of NPPs was based on literature by Van Geel and colleagues (see references below). Plant names are according to van der Meijden (1996). Seeds of the Veronica beccabunga-type represent V. beccabunga, V. anagallis-aquatica and/or V. catenata. Data were visualised with Tilia (Grimm 1991–2011).
Composition of the new pollen sum
The pollen sum proposed in addition to the standard pollen sum excludes the extra-regional taxa, as these trees mask vegetation changes at the dune, especially when they form a substantial part of the total assemblage. For the studied sites in the central-western part of the Rhine-Meuse delta dating between 5500 and 3500 b.c., the extra-regional taxa are Abies, Carpinus, Picea and Pinus. The extra-regional origin of Abies and Picea in the Dutch river area due to fluvial transport from the hinterland during the Atlantic has already been recognised by various authors (Van der Woude 1983, p. 20; Zagwijn 1966). Abies, Carpinus and Picea are indeed found only in very low percentages at Hardinxveld. The fluvial influx implies that a part of the other pollen assemblage also represents extra-regional influx (Hartman 1968; Zagwijn 1966). This will in the first place concern anemophilous pollen produced in large amounts, and aquaphilous pollen (transported by water).
The local presence of Pinus in the river area during the middle Holocene is not completely clear (De Jong and Zagwijn 1983). A possible extra-regional origin of Pinus pollen has been suggested before (Florschütz and Jonker 1939). The absence of Pinus from the region of Hardinxveld and presumably from the wider Dutch delta at the time of the studied occupation is supported by the absence of macroremains and by the dates of rare Pinus wood and charcoal that reveal the presence of reworked Pre-boreal or Boreal material (Out 2010b). Outside the delta, Pinus may have been present at that time due to differences in soil and climate. In the pollen diagrams of Hardinxveld, Pinus mostly reaches 10–20 % when included in the pollen sum, with rare higher frequencies. Instead of pointing to local presence, the higher values can be explained by fluvial transport from the hinterland and in the case of sandy sediment, by selective pollen preservation.
Although all data point to the absence of Fagus from the area during the period considered here, Fagus is not excluded from the new sum since the precise date of its arrival in the region is unknown and may overlap with occupation at the studied sites.
For the off-site diagram of De Bruin the application of the new sum is invalid for the upper part of the core because in the Middle Ages at least some of the excluded taxa were part of the regional vegetation. However, for this paper only the lower part of the diagram is relevant.
Results of the off-site core Hardinxveld-Giessendam De Bruin
Dates
Table 2 provides the AMS dates from botanical macro remains from the off-site core De Bruin, as provided by the Centre for Isotope Research in Groningen, the Netherlands. The calibration is based on the IntCal09 curve of Oxcal version 4.1 (Bronk Ramsey 2009). The core covers at least part of the late Atlantic and Sub-boreal. Date 1 (4686–4369 b.c.) corresponds with the period during or after occupation Phase 3 (4700–4450 b.c.). The reconstruction of the ground water level indicates that peat accumulation at the location of the core started at c. 5200 b.c. (end Phase 1; Louwe Kooijmans 2001b, p. 75), providing a terminus post quem for the lowest sample of the core. Sampling the deepest peat sediments was however hindered by the presence of wood. Date 2 (4350–4052 b.c.) most probably postdates the third and last occupation phase (Table 1). Since the height of the dune at the end of occupation was c. 0.5 m, the submerging of the dune is expected to correspond more or less with this date. At this time, the site was probably already completely submerged (Cohen 2005; Van de Plassche et al. 2010). Pollen of Carpinus, Fagus, Juglans, Fagopyrum and Centaurea cyanus date the upper part of the diagram to the late Sub-atlantic.
Off-site core Hardinxveld-Giessendam De Bruin
Figure 3 shows the pollen, NPP’s and macroremains from the off-site core of De Bruin (ESM 2 provides the raw data). The diagram is divided into four zones. Zone I (7.27–5.97 m NAP) is characterised by high and stable Quercus values. Further important dry land taxa are Tilia, Ulmus, Fraxinus, Corylus, Cornus sanguinea-type, Rhamnus cathartica and Viburnum opulus, pointing to the presence of mixed deciduous woodland with perhaps some open patches. High values of Pinus and Tilia in the lowest sample are likely to be the result of selective corrosion, which may also explain the initial low value of Quercus. Corylus shows a peak in the lower half of the zone, while Fraxinus shows a peak in the upper half of the zone. Dry land herbs and pteridophytes comprise Chenopodiaceae, Artemisia, Plantago lanceolata, P. major/media and Pteridium aquilinum, presumably pointing to human impact (discussed below). A Viscum album pollen grain found in one sample points to the climate optimum of the Atlantic (Zagwijn 1994). This agrees with macroremain finds of another Atlantic indicator at the excavation, Trapa natans (Bakels et al. 2001, p. 399), and the 14C age of the sediment.


Off-site core Hardinxveld-Giessendam De Bruin, pollen, non-pollen palynomorphs and macrofossil diagram. When the x axis does not provide a value, it corresponds to 1 %. Exaggeration, if applied, is fivefold. Macrofossils are indicated by bars. + = 1–5, in the case of sand: present. Dotted line: end of the third (latest) occupation phase based on the interpolation of 14C dates (see also Fig. 4)
At the start of the zone, the wetland taxa Alnus and Cyperaceae show a major peak. This indicates the local presence of alder carr, also supported by the presence of Asteraceae tubuliflorae-type, Galium-type, Mentha-type, Urtica dioica-type, monoletae psilatae fern spores and Type 114 (scalariform perforation plates probably representing decaying Alnus wood), Type 25 (cf. Clasterosporium caricinum), pointing to the local occurrence of periodically flooded Carex and Type 128, pointing to shallow, stagnant to slowly moving, eutrophic fresh water (Pals et al. 1980). The shrubs Cornus sanguinea, Rhamnus cathartica and Viburnum opulus may have been part of the carr. After the initial high values, Alnus pollen values fall back and the Cyperaceae decrease. Alnus macrofossils are present in various samples. The slightly increased curves of Humulus lupulus and Filipendula suggest more open conditions. The appearance of Nuphar lutea-type, Nymphaea alba-type and hair cells and trichosclereids of Nymphaeaceae (Types 127 and 129) indicate the water depth increasing to 2 m at the sample location. Marsh taxa such as Sparganium erectum and Apiaceae gradually increase slightly towards the end of the zone. Together, these changes suggest paludification of the location, therefore a shift of the alder carr to the higher slopes of the dune and the development of marsh vegetation, possibly with mats of floating plants.
The NPPs show high values of Gaeumannomyces (Type 126), indicative of local occurrence of Carex, Diporotheca rizophilla (Type 143), indicative of eutrophic to mesotrophic conditions and possibly of Thelypteris, Gelasinospora (Type 1, included in the genus Neurospora), indicative of dry conditions and often, but not exclusively, occurring on charcoal substrates, and Type 361, indicative of redeposited sand (Van Geel and Aptroot 2006; Van Geel et al. 1981). At the Hazendonk, Type 361 corresponds with human occupation, while also occurring independently of occupation (Out 2010a). The potential indicator of human impact Ustulina deusta (Type 44, Kretzschmaria deusta), a parasite of stumps and dead roots of deciduous trees causing soft-rot (Van Geel and Aptroot 2006, p. 324; Innes et al. 2006) is present in all zones and does not seem to be related to occupation here.
Zone II (5.97–4.12 m NAP) is divided into Subzones IIa and IIb at −5.37 m. The subzone is characterised by a fall in Quercus in Subzone IIa, corresponding to an increase of Fraxinus and a peak of Viburnum opulus, followed in Subzone IIb by a decrease of the latter and a gradual decrease of Corylus. The relative abundance of Quercus rises in Subzone IIb, but the relative abundance of the Lycopodium marker indicates a decline of the absolute pollen concentration. Viburnum opulus, important in Subzone IIa, favours moist locations with rather eutrophic, somewhat mineral, humic to peaty soils; conditions that are found on sandy river clay and dune sand. The shrub is found in the moister parts of dry land vegetation like ash-elm woodland and in the dryer zones of wetland vegetation such as alder carr (Weeda et al. 1988, p. 270). The decrease of Quercus, the peak of Viburnum, an observed peak of Apiaceae and a decrease in Cyperaceae in Zone IIa may result from human impact, as similar changes have been observed during occupation on other river dunes (Out 2009, pp. 54, 271, App. 3, p. 172). The relationship with human activity is however tenuous.
Subzone IIb is a wetter phase with fluctuating water levels. As explained, date 2 at the beginning of Subzone IIb, approximately corresponds to the drowning of the dune. Fluctuations of the presumably non-local taxa Pinus and Betula and a restricted but continuous Picea curve in the upper part of the zone probably reflect the increasing influence of river water, as pollen from conifers primarily originates from the hinterland at this time (Van der Wiel 1982, p. 72; Van der Woude 1983, p. 20). Pollen influx from river water in this zone is confirmed by an increase in the wetland herbs and unidentified pollen. Fraxinus and Ulmus show small peaks, possibly representing regional alluvial woodland vegetation. Small peaks of Artemisia and Plantago lanceolata may result from human activity but probably indicate natural disturbance or represent pollen transported by the river water here, since these taxa are part of the natural vegetation along levees (Groenman-van Waateringe 1968).
Subzone IIb is further characterised by increased values of the wetland taxa Salix, Sparganium emersum-type, S. erectum, Alisma-type, Urtica dioica-type and Cyperaceae at the beginning and the end of the subzone. Together with Alnus macrofossils in the first half of the subzone, a small Alnus pollen peak at the end of the subzone and increased values of Filipendula, Lythrum salicaria and to a lesser degree Typha latifolia point to eutrophic marshes with perhaps some mesotrophic patches. The algae Spirogyra and Mougeotia suggest the presence of shallow, stagnant, mesotrophic to eutrophic water, while the onset of the continuous curve of Type 128 points to shallow, mesotrophic to eutrophic, stagnant or slowly moving water (Pals et al. 1980). In the middle of the subzone, water levels may have been slightly deeper or floods may have occurred, as indicated by lower values of some of the above-mentioned marsh taxa and the presence of macrofossils of Nuphar lutea and Nymphaea alba.
Zone III (4.12–2.67 m NAP) is characterised by the regular occurrence of Fagus (<1.5 %) and low values of upland shrubs and herbs. Quercus shows slightly increased values, Tilia slightly decreased values. The continuous presence of alder carr is indicated by the wetland taxa, mostly stable and similar to those of the previous phase, with exception of peaks of Urtica dioica-type, monoletae psilatae fern spores and scalariform perforation plates at the end of the zone.
Zone IV (2.67–1.87 m NAP), dating to the Sub-atlantic, is characterised by the appearance of Carpinus and a peak of crop plants and arable weeds (c. 20 %). The sudden changes at −2.67 m NAP suggest a hiatus. The dry land vegetation shows an increase of Pinus, a decrease of Quercus and Fraxinus and small peaks of the dry land shrubs Sambucus nigra-type and Ligustrum, together pointing to large-scale regional woodland clearance. The zone yields pollen of Juglans, known in the Netherlands from the Roman period onwards and cultivated at least since the medieval period (Pals 1997). The crops Cerealia-type and Fagopyrum are present. Additionally, one observes the presence or peaks of the potential arable weeds Artemisia, Chenopodiaceae, Plantago lanceolata, Polygonum sect. persicaria, P. aviculare, Plantago major/media, Centaurea cyanus, C. jacea-type and Spergula arvensis. Centaurea cyanus pollen and fruits of Fagopyrum are well known in the Netherlands since the 14th century a.d. (Janssen 1974; Van Haaster 1997). Poaceae show a major peak, as well as the ecologically indeterminate Apiaceae, Asteraceae tubuliflorae-type, Asteraceae liguliflorae-type, Brassicaceae, Galium-type and Ranunculus acris-type. These changes indicate a substantial increase of agricultural practices. The NPPs show high values, for example of Gaeumannomyces (Type 126, discussed above), cf. Arcella discoides (Type 352), a testate amoeba with wide ecological amplitude, and Type 361, indicative of redeposited sand. The wetland taxa remain relatively stable, suggesting the continued presence of alder carr and eutrophic marshes with local mesotrophic patches.
Discussion
Vegetation at Hardinxveld-Giessendam De Bruin
In general, the new pollen diagram of Hardinxveld-Giessendam De Bruin indicates similar vegetation for the Atlantic and Sub-boreal as do the published diagrams of Polderweg and De Bruin. On the higher parts of the dune, deciduous woodland was present, dominated by Tilia, Quercus, Corylus avellana and perhaps Fraxinus excelsior, Ulmus and Humulus lupulus. This woodland was moist, dense and of considerable age. The occurrence of Polypodium vulgare confirms the presence of shady parts. On the lower parts of the slope, Tilia and Quercus were scarcer, and Fraxinus excelsior and Ulmus more common. The dry land and wetland vegetation elements will not have been strictly separated as suggested by the distinction of groups in the pollen diagram (Binney et al. 2005). On those parts of the slope under the influence of ground water, alder carr dominated by Alnus glutinosa occurred, presumably also including shrubs such as Rhamnus cathartica, Rhamnus frangula (Frangula alnus), Viburnum opulus and Cornus sanguinea. The alder carr was generally dense and old, but probably less stable than the dry land vegetation due to local changes in the wetland environment. Around the dune, marshes occurred, characterised by forb vegetation, marsh vegetation and water plants. These wetlands were eutrophic with only minor mesotrophic patches. The various marsh plants such as Cyperaceae and Filipendula and NPPs point to the presence of shallow pools with a depth of 0.5 m, possibly drying out during summers, while the regular occurrence of Nymphaeaceae in all zones except Zone 3 points to water depths up to 1.5–2 m. As a result of the rise of the mean water level and the drowning of the dune, the various vegetation belts moved upwards along the slope through time.
Comparison of the off-site core results with the data from the macrofossils, wood and charcoal from the excavation (Bakels and Van Beurden 2001; Bakels et al. 2001; Louwe Kooijmans et al. 2001a, b) confirms the local presence of Tilia, Quercus, Corylus avellana, Fraxinus excelsior, Ulmus, Rhamnus cathartica, Cornus sanguinea, Alnus glutinosa, Salix, Rhamnus frangula and Viburnum opulus. All these taxa were identified as uncarbonised macrofossils, except for Ulmus and Salix, which rarely remain preserved in such a state. Taxa that were only attested irregularly and in small percentages in the unworked or worked wood, charcoal and macrofossils, thus probably not presenting local vegetation, are Acer campestre, Betula and Viscum album. Taxa that have been attested in the wood and/or charcoal but not in the pollen assemblage are Euonymus, Pomoideae (subfamily of the Rosaceae including Malus, Pyrus, Sorbus and Crataegus) and Prunus. Of these, Crataegus monogyna, Malus sylvestris and Prunus padus were also attested in the macrofossil assemblage; the first two both uncarbonised and carbonised. The local presence of Euonymus, Pomoideae and Prunus however remains unclear; these entomophilous taxa are usually underrepresented in the pollen rain and the macrofossils and wood may have been collected elsewhere (Out 2009).
Human impact at Hardinxveld-Giessendam De Bruin
Human impact related to the occupation by people of the Final Mesolithic Swifterbant Culture is expected in Zone I and perhaps in a part of Subzone IIa of the off-site core. Together these zones correspond with occupation Phases 2 and 3. Archaeological indicators related to occupation during Phases 2 and 3 were registered not only in and near the excavation trench but also south of the dune at c. 23 m distance from the sampling location (Fig. 2). Evidence of human activity is however very scarce: the non-arboreal pollen curve does not show major fluctuations, suggesting that any small-scale clearings were soon overgrown again. Possible indications of human impact in Zone I, presumably corresponding with the end of Phase 2 or with Phase 3 as indicated by date 1 (4686–4369 b.c., a terminus ante quem), include a peak of Corylus, indicating more open vegetation, and the presence of Chenopodiaceae, Artemisia, Plantago lanceolata, P. major/media and Pteridium aquilinum, pointing to disturbance, trampling and burning of vegetation, and NPPs indicative of redeposited sand, charcoal and decaying plant material. The earlier published diagram of De Bruin (ESM 1) also shows a peak of Corylus and the presence of human indicators including Chenopodiaceae and Artemisia during Phases 2 and 3, indicating that similar changes occurred on the eastern side of the dune. As explained in the introduction, the Corylus peak during the middle of Phase 2 (−7.40 m NAP) is clearly related to human impact. This supports the hypothesis that the Corylus peak observed in the off-site core is also indicative of human impact. In Subzone IIa, the decrease of Quercus, the peak of Viburnum, the presence of Artemisia, Plantago lanceolata and Polygonum sect. persicaria and changes in the wetland herb vegetation may also result from human impact. These changes can however also be explained by tree falls caused by the rising water level, activities of animals such as beavers and deer, and river activity such as high water levels affecting the vegetation. River influence is indicated by the almost continuous curve of Chenopodiaceae in both zones, presumably representing drift litter and riverbank vegetation (Cappers 1993; Weeda et al. 1988, p. 157) and the presence of pollen grains of Picea in Subzone IIa (discussed above). Indeed, several of these taxa also occur in the diagram after the drowning of the dune when occupation was no longer possible.
Concluding, the evidence of human impact at the Final Mesolithic site of Hardinxveld-Giessendam De Bruin observed in the pollen diagrams is weak and it remains difficult to separate human impact from other disturbances (cf. Brown 1997; Day and Mellars 1998, p. 112; Timpany 2005). This can be related to the mostly non-agricultural character of the site (see Introduction) and its presumed function as a seasonal extraction camp during Phase 3 (Louwe Kooijmans 2001b, p. 534). Similarly, the pollen diagram from Polderweg did not provide clear evidence of human impact during any of the occupation phases either. This is despite the fact that this core was located northeast of the dune and might be expected to register the pollen rain of the dune better than the off-site core at De Bruin since the prevailing wind in the Netherlands is usually coming from a southwesterly direction. Apart from occupation-related aspects, the restricted evidence of human impact at the off-site core of De Bruin can also be explained by the great distance to the nearest dune patch (c. 30 and 35 m during Phases 2 and 3), the distance to the main part of the find layer (c. 40 m during all phases), and the presence of alder carr hampering the deposition of the dune vegetation pollen rain at the sample location. This hypothesis is also supported by the Polderweg core, located at a maximum of 26 m from the dune edge and during all phases c. 40 m from the refuse layer. Even at the agricultural site of the Hazendonk the evidence of human impact on the vegetation south of the dune is weak and decreases over a distance of a few metres (Out 2008b, 2010a; Van der Wiel 1982, Van der Woude 1983).
Evidence of neolithisation in pollen diagrams from Dutch wetland sites?
Figure 4a presents the main diagrams of Polderweg, De Bruin, Brandwijk-Kerkhof and the Hazendonk. Their comparison shows that the evidence of human impact is restricted at Hardinxveld-Giessendam Polderweg and De Bruin (as discussed above) and expressed more strongly during the Early Neolithic phases at Brandwijk-Kerkhof and the Hazendonk which yielded evidence of crop plants (Brandwijk-Kerkhof: layers 50 and 60; Hazendonk: including Phases Haz 1, 2 and 3). At Brandwijk-Kerkhof, occupation related to layers 50 and/or 60 can be detected in both cores D and B and is characterised by a decrease of arboreal pollen and an increase of upland shrubs, herbs and spore plants. At the Hazendonk, the major occupation phases are marked by similar changes. The complete diagrams (Out 2008a, 2009) show decreases particularly of Tilia, Quercus and Alnus glutinosa, coinciding with peaks of various shrubs and both dry land and wetland herbs including Cerealia-type. The recovery of the vegetation after occupation, the recognition of archaeological layers in the cores and the presence of Cerealia-type pollen allows the linking of these signals to human activity, thus ruling out non-human disturbance factors. However, the increased evidence of human impact and the presence of Cerealia pollen are not automatically related to local farming, since the cereal pollen may also result from the processing of crop products imported from more southern producer sites (Out 2008c).

Main pollen diagrams of Hardinxveld-Giessendam Polderweg, Hardinxveld-Giessendam De Bruin on-site and off-site, Brandwijk-Kerkhof and the Hazendonk, based on a, an upland pollen sum including extra-regional taxa; b, an upland pollen sum excluding the extra-regional taxa Abies, Carpinus, Pinus and Picea. The sequences are lined up according to the age of the deposits. Dotted lines De Bruin off-site: end of the second and third occupation phase based on the interpolation of 14C dates
The difference in the visibility of human impact between the Final Mesolithic and Early Neolithic sites and phases may be related to the degree of neolithisation of these communities and the introduction of (live) domestic animals and cereals. Another possible cause is the research methodology. The cores of Brandwijk-Kerkhof and the Hazendonk were analysed in more detail by application of smaller sample intervals and were taken at the border of sites where find layers wedge out into the peat. In contrast, the cores at Hardinxveld-Giessendam were sampled either on the site or quite far away. It is expected that more detailed sampling of comparable Final Mesolithic sites at their margins, where find layers wedge out into the peat, will enable a clearer distinction of the development before, during and after occupation.
The restricted evidence of Neolithic impact at the Dutch wetland sites considered here is not exceptional (see Introduction and Van der Linden 2010). Comparable wetland sites show both corresponding and other results concerning the relation between neolithisation and human impact. Varying factors such as site function and the research methodology probably explain the absence of a general pattern. Eight core samples from Alblasserdam-Nieuw Kinderdijk, representing the Mesolithic/Neolithic transition, showed very poor evidence of human impact, if at all, that remained stable through time (De Koning and Van Dasselaar 2008). At a site in the Noordoostpolder, human impact indicators including cereal pollen and apophytes increased after the introduction of agriculture (Weijdema et al. 2011). On the other hand, a pollen diagram from the Mesolithic site of Rotterdam-Randstad allowed the discerning of human impact on the vegetation despite the non-agricultural function of the site (Guiran and Brinkkemper 2007).
Application of a new pollen sum excluding extra-regional taxa
To obtain an optimal understanding of human impact on the vegetation near to archaeological sites, this paper investigates the application of a pollen sum that excludes extra-regional taxa, applied to De Bruin and comparable sites. It is expected that the main diagrams based on the new pollen sum should give more direct information on the development of the vegetation of the inland dunes than those based on the traditional pollen sum, since fluctuations of extra-regional pollen are excluded. Since the composition of the regional vegetation changes through the Holocene and differs between regions, the new pollen sum is applicable during a restricted period and for a restricted area only. The new pollen sum is therefore proposed to be used in addition to the usual pollen sum. It can furthermore be adapted in other studies according to information on the presence/absence of potentially extra-regional taxa.
Figure 4 shows the main diagrams of Polderweg, De Bruin, Brandwijk-Kerkhof and the Hazendonk, with one version (a) based on the usual upland pollen sum including dry land trees, shrubs, herbs, spore plant and crops and another version (b) based on the same upland pollen sum but excluding the extra-regional taxa (see also “Materials and methods” section). Compared with the diagrams based on the original sum, the diagrams based on the pollen sum excluding extra-regional taxa occasionally show slightly more stable curves (for example De Bruin off-site core: −4.87 m NAP), a small decrease of the dry land arboreal pollen and a small increase of the upland shrubs (for example Brandwijk core D, −5.81 m NAP) and a small increase of the clustered group of herbs, ferns and crop plants (for example Brandwijk core B, −5.00 m NAP). A diagram that shows a greater effect of the new sum is the on-site diagram from De Bruin. While the values of upland herbs and spore plants are negligible when using the original sum (0–3 %) and therefore poorly visible in the main diagram, these taxa are clearly better represented in the diagram based on the new sum, demonstrating the usefulness of the new pollen sum.
Although it is possible to observe differences between the presented main diagrams based on the two pollen sums, the differences are restricted. This may however differ in other case studies. One explanation for the restricted differences is the number of pollen included in the pollen sum. In case of core B and to a lesser degree core D from Brandwijk, the number of pollen included in the new sum is rather low because earlier collected data were recalculated. When the new pollen sum would have been applied during the counting of 300 grains of the new sum, this would presumably have resulted in better information about the local and extra-local vegetation. Indeed, the diagrams of the Hazendonk, in contrast to more recently collected data-sets based on a pollen sum regularly including 350–450 upland pollen grains, have demonstrated an unusually large variety of pollen types and provide a very detailed picture of vegetation development and human impact (investigation Louwe-Kooijmans; Out 2009, App. 3).
Another explanation is the character of the sediment influencing the importance of extra-regional pollen. Extra-regional pollen transported by river water is particularly expected in clayey sediments. Clay has been observed in the upper part of Core 3 from the Hazendonk, which indeed shows subtle differences between the two main diagrams between −4.15 and −3.95 m NAP. In the upper part of the on-site sample series from De Bruin, clayey peat was observed, but this unexpectedly does not result in greater changes than in the remaining part of the diagram. In the other cores, clay was not noticed during analysis, which explains the absence of larger differences. Besides, overrepresentation of extra-regional gymnosperm pollen with sacci also occurs in sandy sediment, resulting in selective preservation. Sand is present in the lower parts of some cores where the transition from the dune sand to peat is sampled. Indeed, at these levels the diagrams of Polderweg, De Bruin off-site, and the Hazendonk Square 57, calculated on the new pollen sum, show subtle differences. In future studies, for example in studies in the Dutch river delta where influx of extra-regional pollen in river water is expected, it may be useful to count sacci of gymnosperm pollen, to get an understanding of the importance of extra-regional pollen and the relevance of applying the proposed new pollen sum.
Conclusions
The data on the vegetation development and human impact obtained from the off-site core at the Final Mesolithic site of De Bruin correspond well with earlier results. The dune vegetation consisted of deciduous woodland on the higher slopes combined with alder carr and mostly eutrophic freshwater marsh vegetation on the lower slopes and in the surrounding wetlands. Although non-human disturbance cannot be ruled out, an increase of Corylus and the presence of Chenopodiaceae, Artemisia, Plantago lanceolata, P. major/media and Pteridium aquilinum occurring shortly before 4690–4360 b.c. and probably related to occupation Phase 3 (4700–4450 b.c.) off-site is possibly related to human activities. Comparable Corylus peaks occur during Phases 1, 2 and 3 in the on-site sequence, of which at least the second peak during Phase 2 is linked to human activities. The sites of Hardinxveld do not provide evidence of Mesolithic arable farming.
Despite the ongoing neolithisation, reflected in the archaeological finds, the pollen diagrams of Hardinxveld do not clearly show increasing or changing levels of human influence on the vegetation through time, independent of site function and occupation intensity, indicating stable plant exploitation strategies. At the agricultural sites of Brandwijk-Kerkhof and the Hazendonk, the phases yielding cereals show clearer evidence of human impact. This increased visibility may be related to the introduction of domestic animals and crop plants, but also to the research methodology. Detailed archaeological and archaeobotanical analyses of comparable sites in the region, and particularly those covering the period 4500–4000 b.c., may provide better understanding of the neolithisation process and how people affected their environment.
Recommendations for future research at comparable sites in the Rhine-Meuse river delta and contemporary sites in similar delta environments concern the research methodology. The first advice concerns the sample location with respect to dunes or dry land terrain. The expected activity zones should be within a range of 25 m from the pollen sample location, for both Mesolithic and Neolithic sites. Sampling along transects where find layers wedge out into the peat will make it easier to relate the vegetation development to occupation. It may be relevant to take the prevailing wind direction into consideration. Secondly, calculation of pollen percentages on a standard pollen sum and an upland sum excluding extra-regional taxa is recommended. It is expected that the removal of the extra-regional taxa will result in the counting of more pollen of the local and extra-local vegetation; this increases the chances to encounter taxa of rare and entomophilous shrubs and herbs including anthropogenic indicators and provide better insights into how people affected the vegetation.
References
Bakels CC, Van Beurden LM (2001) Archeobotanie. In: Louwe Kooijmans LP (ed) Archeologie in de Betuweroute. Hardinxveld-Giessendam Polderweg. Rapp Archeol Monumentenzorg, vol 83, Amersfoort, pp 325–378
Bakels CC, Van Beurden LM, Vernimmen TJJ (2001) Archeobotanie. In: Louwe Kooijmans LP (ed) Archeologie in de Betuweroute. Hardinxveld-Giessendam De Bruin. Rapp Archeol Monumentenzorg, vol 88, Amersfoort, pp 369–434
Beerenhout B (2001) Vissen. In: Louwe Kooijmans LP (ed) Archeologie in de Betuweroute. Hardinxveld-Giessendam De Bruin. Rapp Archeol Monumentenzorg, vol 88, Amersfoort, pp 299–326
Behre K-E (1981) The interpretation of anthropogenic indicators in pollen diagrams. Pollen Spores 23:225–245
Behre K-E, Kučan D (1986) Die Reflektion archäologisch bekannter Siedlungen in Pollendiagrammen verschiedener Entfernung – Beispiele aus der Siedlungskammer Flögeln, Nordwestdeutschland. In: Behre K-E (ed) Anthropogenic indicators in pollen diagrams. Balkema, Rotterdam, pp 95–115
Binney HA, Waller MP, Bunting MJ, Armitage RA (2005) The interpretation of fen carr pollen diagrams: the representation of the dry land vegetation. Rev Palaeobot Palynol 134:197–218
Birks HJB, Birks HH (1980) Quaternary palaeoecology. Arnold, London
Bos JAA, Janssen CR (1996) Local impact of Palaeolithic man on the environment during the end of the last glacial in the Netherlands. J Archaeol Sci 23:731–739
Bos JAA, Urz R (2003) Late Glacial and early Holocene environment in the middle Lahn river valley (Hessen, central-west Germany) and the local impact of early Mesolithic people—pollen and macrofossil evidence. Veget Hist Archaeobot 12:19–36
Bos JAA, Van Geel B, Groenewoudt BJ, Laurier RCGM (2005) Early Holocene environmental change, the presence and disappearance of early Mesolithic habitation near Zutphen (the Netherlands). Veget Hist Archaeobot 15:27–43
Bronk Ramsey C (2009) Bayesian analysis of radiocarbon dates. Radiocarbon 51:337–360
Brown T (1997) Clearances and clearings: deforestation in Mesolithic/Neolithic Britain. Oxf J Archaeol 16:133–146
Brown AG (1999) Characterising Prehistoric lowland environments using local pollen assemblages. Quat Proc 7:585–594
Bunting MJ, Armitage R, Binney HA, Waller M (2005) Estimates of ‘relative pollen productivity’ and ‘relevant source area of pollen’ for major tree taxa in two Norfolk (UK) woodlands. Holocene 15:459–465
Cappers RTJ (1993) Seed dispersal by water: a contribution to the interpretation of seed assemblages. Veget Hist Archaeobot 2:173–186
Cohen K (2005) A model for groundwater rise. www.geo.uu.nl/paleogeography. Accessed 1 Sept 2012
Day P, Mellars P (1998) Star carr in context. Oxbow Books, Oxford
De Jong J, Zagwijn WH (1983) De vegetatiegeschiedenis van, s-Gravenhage en omgeving. Meded Rijks Geol Dienst 37:44–62
De Koning MWA, Van Dasselaar M (2008) Archeologisch onderzoek plan Nieuw Kinderdijk te Alblasserdam. Archeomedia rapport A06-189-R. Archeomedia, Capelle aan den IJssel
Fægri K, Iversen J (1989) Textbook of pollen analysis, 4th edn. (by Fægri K, Kaland PE and Krzywinski K). Wiley, Chichester
Florschütz F, Jonker FP (1939) A botanical analysis of a Late-Pleistocene and Holocene profile in the Rhine delta. Rec Trav Bot Neerl 36:686–696
Grimm E (1991–2011) TILIA v1.7.16. Illinois State Museum, Springfield
Groenman-van Waateringe W (1968) The elm decline and the first appearance of Plantago major. Vegetatio 15:292–296
Guiran AJ, Brinkkemper O (2007) Archeologisch onderzoek 1 Emplacement Central Station, Rotterdam. BOORrapporten 318. Bureau Oudheidkundig Onderzoek, Rotterdam (in cooperation with Brinkhuizen DC, Klink A Rotterdam-Randstadrail)
Hartman AA (1968) A study on pollen dispersal and sedimentation in the western part of the Netherlands. Acta Bot Neerl 17:506–549
Hijma MP, Cohen KM (2011) Holocene transgression of the Rhine river mouth area, the Netherlands/Southern North Sea: paleogeography and sequence stratigraphy. Sedimentology 58:1,453–1,485
Hörnberg G, Bohlin E, Hellberg E, Bergman I, Zackrisson O, Olofsson A, Wallin J-E, Påsse T (2006) Effects of Mesolithic hunter-gatherers on local vegetation in a non-uniform glacio-isostatic land uplift area, northern Sweden. Veget Hist Archaeobot 15:13–26
Innes J, Blackford J, Chambers F (2006) Kretzschmaria deusta and the Northwest European Mid-Holocene Ulmus decline at Moel Y Gerddi, North Wales, United Kingdom. Palynol 30:121–132
Jackson ST (1994) Pollen and spores in Quaternary lake sediments as sensors of vegetation composition: theoretical models and empirical evidence. In: Traverse A (ed) Sedimentation of organic particles. Cambridge University Press, Cambridge, pp 253–286
Janssen CR (1959) Alnus as a disturbing factor in pollen diagrams. Acta Bot Neerl 8:55–58
Janssen CR (1970) Problems in the recognition of plant communities in pollen diagrams. Vegetatio 20:187–198
Janssen CR (1974) Verkenningen in de palynologie. Oosthoek, Scheltema and Holkema
Joly C, Visset L (2009) Evolution of vegetation landscapes since the Late Mesolithic on the French West Atlantic coast. Rev Palaeobot Palynol 154:17–124
Kalis AJ, Meurers-Balke J (1998) Die “Landnam”-Modelle von Iversen und Troels-Smith zur Neolithisierung des westlichen Ostseegebietes – ein Versuch ihrer Aktualisierung. Prähist Z 73:1–24
Kuneš P, Pokorný P, Šída P (2007) Detection of the impact of early Holocene hunter-gatherers on vegetation in the Czech Republic, using multivariate analysis of pollen data. Veget Hist Archaeobot 17:269–287
Louwe Kooijmans LP (ed) (2001a) Archeologie in de Betuweroute. Hardinxveld-Giessendam Polderweg. Rapp Archeol Monumentenzorg, vol 83. ROB, Amersfoort
Louwe Kooijmans LP (ed) (2001b) Archeologie in de Betuweroute. Hardinxveld-Giessendam De Bruin. Rapp Archeol Monumentenzorg, vol 88. ROB, Amersfoort
Louwe Kooijmans LP (2003) The Hardinxveld sites in the Rhine/Meuse delta, the Netherlands, 5500–4500 cal b.c.. In: Larsson L, Kindgren H, Knutsson K, Leoffler D, Åkerlund A (eds) Mesolithic on the move. Oxbow, Oxford, pp 608–624
Louwe Kooijmans LP (2007) The gradual transition to farming in the Lower Rhine Basin. In: Whittle A, Cummings V (eds) Going over: the Mesolithic–Neolithic transition in north-west Europe. Oxford University Press, Oxford, pp 287–309
Louwe Kooijmans LP, Mol J (2001) Stratigrafie, chronologie en fasering. In: Louwe Kooijmans LP (ed) Archeologie in de Betuweroute. Hardinxveld-Giessendam Polderweg. Rapp Archeol Monumentenzorg, vol 83, Amersfoort, pp 55–72
Louwe Kooijmans LP, Hänninen K, Vermeeren CE (2001a) Artefacten van hout. In: Louwe Kooijmans LP (ed) Archeologie in de Betuweroute. Hardinxveld-Giessendam De Bruin. Rapp Archeol Monumentenzorg, vol 88, Amersfoort, pp 435–478
Louwe Kooijmans LP, Vermeeren CE, Van Waveren AMI (2001b) Hout en artefacten van vezels. In: Louwe Kooijmans LP (ed) Archeologie in de Betuweroute. Hardinxveld-Giessendam Polderweg. Rapp Archeol Monumentenzorg, vol 83, Amersfoort, pp 379–418
Mol J (2001a) Landschapsgenese en paleogeografie. In: Louwe Kooijmans LP (ed) Archeologie in de Betuweroute. Hardinxveld-Giessendam Polderweg. Rapp Archeol Monumentenzorg, vol 83, Amersfoort, pp 35–54
Mol J (2001b) Landschapsgenese en paleogeografie. In: Louwe Kooijmans LP (ed) Archeologie in de Betuweroute. Hardinxveld-Giessendam De Bruin. Rapp Archeol Monumentenzorg, vol 88, Amersfoort, pp 33–56
Mol J (2003) Landscape evolution and site formation of two Mesolithic sites in the lower Rhine-Meuse delta (Hardinxveld, the Netherlands). In: Howards AJ, Macklin MG, Passmore DG (eds) Alluvial archaeology in Europe. Balkema, Lisse, pp 147–161
Mol J, Louwe Kooijmans LP (2001) Stratigrafie, chronologie en fasering. In: Louwe Kooijmans LP (ed) Archeologie in de Betuweroute. Hardinxveld-Giessendam De Bruin. Rapp Archeol Monumentenzorg, vol 88, Amersfoort, pp 57–74
Mol J, Van Zijverden W (2007) Prehistorische woonplaatsen in de dynamische landschappen van de Rijn-Maasdelta. In: Jansen R, Louwe Kooijmans LP (eds) Tien jaar Archol. Van contract tot wetenschap. Archol BV, Leiden, pp 89–101
Moore PD, Webb JA, Collinson ME (1991) Pollen analysis. Blackwell, London
Out WA (2008a) Neolithisation at the site Brandwijk-Kerkhof, the Netherlands: natural vegetation, human impact and plant food subsistence. Veget Hist Archaeobot 17:25–39
Out WA (2008b) The scale of human impact at the Hazendonk, the Netherlands, during the Late Neolithic. Analecta Praehist Leiden 40:153–165
Out WA (2008c) Growing habits? Delayed introduction of crop cultivation at marginal Neolithic wetland sites. Veget Hist Archaeobot 17(Supplement 1):131–138
Out WA (2009) Sowing the seed? Human impact and plant subsistence in Dutch wetlands during the Late Mesolithic and Early and Middle Neolithic (5500–3400 b.c.). Archaeological Studies Leiden University, vol 18. University Press, Leiden
Out WA (2010a) Integrated archaeobotanical analysis: human impact at the Dutch Neolithic wetland site the Hazendonk. J Archaeol Sci 37:1521–1531
Out WA (2010b) Firewood collection strategies at Dutch wetland sites in the process of neolithisation. Holocene 20:191–204
Pals JP (1997) De introductie van cultuurgewassen in de Romeinse tijd. In: Zeven AC (ed) De introductie van onze cultuurplanten en hun begeleiders, van het Neolithicum tot 1500 a.d. Vereniging voor Landbouwgeschiedenis, Wageningen, pp 25–52
Pals JP, Van Geel B, Delfos A (1980) Paleoecological studies in the Klokkeweel bog near Hoogkarspel (prov. of Noord-Holland). Rev Palaeobot Palynol 30:371–418
Poska A, Saarse L, Veski S (2004) Reflections of pre- and early-agrarian human impact in the pollen diagrams of Estonia. Palaeogeogr Palaeoclimatol Palaeoecol 209:37–50
Punt W, Blackmore S, Clarke GCS, Hoen PP (1976–1995) The Northwest European pollen flora, vol I (1976); vol 2 (1980), vol 3 (1981), vol 4 (1984), vol 5 (1988), vol 6 (1991), vol 7 (1995). Elsevier, Amsterdam
Raemaekers DCM (1999) The articulation of a ‘New Neolithic’. Archaeological Studies Leiden University, vol. 3. Faculty of Archaeology, Leiden University, Leiden
Regnell M (2012) Plant subsistence and environment at the Mesolithic site Tågerup, southern Sweden: new insights on the “Nut Age”. Veget Hist Archaeobot 21:1–16
Reille M (1992) Pollen et spores d’Europe et d’Afrique du Nord. Laboratoire de Botanique historique et Palynologie, Marseille
Rowley-Conwy P (2013) North of the frontier: early domestic animals in Northern Europe. In: Colledge S, Conolly J, Dobney K, Manning K, Shennan S (eds) The origins and spread of domestic animals in Southwest Asia and Europe. Left Coast Press, Walnut Creek
Schiltmans DEA, in cooperation with Houkes RA, Kooistra LI (2010) Rotterdam IJsselmonde ‘t Hart. Een opgraving ter plaatse van het nieuwe stadshart van IJsselmonde (vindplaats 13-66). BOORrapporten 424. Bureau Oudheidkundig Onderzoek, Rotterdam
Sugita S (1994) Pollen representation of vegetation in Quaternary sediments: theory and method in patchy vegetation. J Ecol 82:881–897
Timpany S (2005) A palaeoecological study of submerged forests in the Severn estuary and Bristol Channel, UK. Unpubl. PhD thesis, Reading University
Van de Plassche O, Makaske B, Hoek WZ, Konert M, Van der Plicht J (2010) Mid-Holocene water-level changes in the lower Rhine-Meuse delta (western Netherlands): implications for the reconstruction of relative mean sea-level rise, palaeoriver-gradients and coastal evolution. Neth J Geosci 89:3–20
Van der Linden M (2010) Verlaten donken onder het veen? Paleoecologisch onderzoek aan een veenpakket bij Dinteloord. BIAXiaal 493. BIAX Consult, Zaandam
Van der Meijden R (1996) Heukels’ flora van Nederland, 23rd edn. Wolters-Noordhoff, Groningen
Van der Wiel AM (1982) A palaeoecological study of a section from the foot of the Hazendonk (Zuid-Holland, the Netherlands), based on the analysis of pollen, spores and macroscopic plant remains. Rev Palaeobot Palynol 38:35–59
Van der Woude JD (1983) Holocene paleoenvironmental evolution of a perimarine fluviatile area. Geology and paleobotany of the area surrounding the archaeological excavation at the Hazendonk river dune (western Netherlands). Analecta Praehist Leiden, vol 16. Faculty of Archaeology, Leiden University, Leiden
Van Geel B, Aptroot A (2006) Fossil ascomycetes in Quaternary deposits. Nova Hedwigia 82:313–329
Van Geel B, Bohncke SJP, Dee H (1981) A palaeoecological study of an upper Late Glacial and Holocene sequence from “De Borchert”, the Netherlands. Rev Palaeobot Palynol 31:415–445
Van Geel B, Buurman J, Brinkkemper O, Schelvis J, Aptroot A, Van Reene G, Hakbijl T (2003) Environmental reconstructions of a Roman Period settlement site in Uitgeest (the Netherlands), with special reference to coprophilous fungi. J Archaeol Sci 30:873–883
Van Haaster H (1997) De introductie van cultuurgewassen in de Nederlanden tijdens de Middeleeuwen. In: Zeven AC (ed) De introductie van onze cultuurplanten en hun begeleiders, van het Neolithicum tot 1500 a.d. Vereniging voor Landbouwgeschiedenis, Wageningen, pp 53–104
Van Zeist W (1967) Archaeology and palynology in the Netherlands. Rev Palaeobot Palynol 4:45–65
Vanmontfort B (2008) A southern view on north-south interaction during the Mesolithic-Neolithic transition in the Lower Rhine Area. Analecta Praehist Leiden 40:85–97
Verbruggen M (1992) Geoarchaeological prospection of the Rommertsdonk. Analecta Praehist Leiden 25:117–128
Verhoeven K (2003) Palynologisch onderzoek op een veenboring uit de nabijheid van een mesolithische donkbewoning te Hardinxveld-Giessendam De Bruin (Nederland). Unpublished MA thesis Faculty of Archaeology, Leiden University, the Netherlands
Vos P, Bazelmans J, Weerts HJT, Van der Meulen M (2011) Atlas van Nederland in het Holoceen. Bakker, Amsterdam
Waller MP, Binney HA, Bunting MJ, Armitage RA (2005) The interpretation of fen carr pollen diagrams: pollen-vegetation relationships within the fen carr. Rev Palaeobot Palynol 133:179–202
Weeda EJ, Westra R, Westra C, Westra T (1985–1994) Nederlandse oecologische flora. (Wilde planten en hun relaties 1-5) KNNV/IVN, Amsterdam
Weijdema F, Brinkkemper O, Peeters H, Van Geel B (2011) Early Neolithic human impact on the vegetation in a wetland environment in the Noordoostpolder, central Netherlands. J Archaeol Low Countries 3:31–46 http://dpc.uba.uva.nl/jalc/03/nr01/a01. Accessed 1 Sept 2012
Zagwijn WH (1966) Pollen-analytical correlations in the coastal-barrier deposits near The Hague (the Netherlands). Meded Geol Sticht NS 17:83–88
Zagwijn W (1994) Reconstruction of climate change during the Holocene in western and central Europe based on pollen records of indicator species. Veget Hist Archaeobot 3:65–88
Zeiler JT (1997) Hunting, fowling and stock-breeding at Neolithic sites in the western and central Netherlands. Ph.D. thesis University of Groningen, the Netherlands. Archaeobone, Groningen
Zijl W, Niekus MJLTh, Ploegaert PHJI, Moree JM (2011) Rotterdam Beverwaard Tramremise. De opgraving van de top van een donk met sporen uit het Mesolithicum en Neolithicum (vindplaats 13-83). BOORrapporten 439. Bureau Oudheidkundig Onderzoek, Rotterdam
Acknowledgments
The authors thank C.C. Bakels, W.J. Kuijper and L.P. Louwe Kooijmans for their kind support and advice, A. Louwe Kooijmans for inspiring discussion about the pollen sum, O. Brinkkemper and an anonymous reviewer for their helpful and valuable comments, H. van der Plicht and colleagues for dating, L. Amkreutz and J. Mol for fruitful discussions, and M. Doorenbosch for literature suggestions and help with the figures. The research was part of a project on neolithisation in Dutch wetlands (“From Hardinxveld to Noordhoorn: from forager to farmer”), primarily carried out at Leiden University and funded by the Netherlands Organization for Scientific Research (2003–2009). WO is a member of the research cluster Complexity and Socio-Ecological dynamics (cases-bcn.net) and is funded by an EU Marie Curie Intra-European Fellowship (273610). The analysis of the core was carried out by KV for his master thesis at the Faculty of Archaeology, Leiden University, the Netherlands. He is now assistant professor at the Research Unit Palaeontology at Ghent University, where A. Gautier provided valuable suggestions on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by K.-E. Behre.
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
Hardinxveld-Giessendam De Bruin, on-site sample series, square 24, pollen diagram based on an upland pollen sum (Bakels et al. 2001). Exaggeration, if applied, is fivefold (EPS 2047 kb)
ESM 2
Hardinxveld-Giessendam De Bruin, off-site core, pollen, NPP and macrofossil data (XLS 51 kb)
Rights and permissions
About this article
Cite this article
Out, W.A., Verhoeven, K. Late Mesolithic and Early Neolithic human impact at Dutch wetland sites: the case study of Hardinxveld-Giessendam De Bruin. Veget Hist Archaeobot 23, 41–56 (2014). https://doi.org/10.1007/s00334-013-0396-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-013-0396-0