
Abstract
This paper discusses changing patterns of resource utilisation over time in the locality of Chibuene, Vilankulos, situated on the coastal plain of southern Mozambique. The macroscopic charcoal, bone and shell assemblages from archaeological excavations are presented and discussed against the off-site palaeoecological records from pollen, fungal spores and microscopic charcoal. The Chibuene landscape has experienced four phases of land use and resource utilisation that have interacted with changes in the environment. Phase 1 (a.d. 400–900), forest savanna mosaic, low intensity cattle herding and cultivation, trade of resources for domestic use. Phase 2 (a.d. 900–1400), forest savanna mosaic, high intensity/extensive cultivation and cattle herding. Phase 3 (a.d. 1400–1800), savanna woodland and progressive decrease in forests owing to droughts. Decline of agricultural activities and higher reliance on marine resources. Possible trade of resources with the interior. Phase 4 (a.d. 1800–1900), open savanna with few forest patches. Warfare and social unrest. Collapse of trade with the interior. Decline in marine resources and wildlife. Loss of cattle herds. Expansion of agriculture locally and introduction of New World crops and clearing of Brachystegia trees. The study shows the importance of combining different environmental resources for elucidating how land use and natural variability have changed over time.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The understanding and appreciation of changing resource utilisation in southern Africa over time has long been limited owing to a lack of appropriate data. Most discussions on historic resource utilisation, environmental management and land use practices have been based on inferences from modern natural resources, as other data was lacking (Hall 1981; Sinclair 1987; Morais 1988). However, the value of these pioneering landscape studies is fundamental. We need to better understand the historical transformations of the physical landscape and their interaction with land use practices and resource use (Jonsson 1998; Eichhorn 2002; Manyanga 2006). Despite systematic efforts to retrieve environmental data from archaeological sites, the ideal combination of ecofacts such as seeds, bones, shell etc. from archaeological sites and off site palaeoecological analyses yielding a broad overview of local and extra-local changes in vegetation and landscape changes is extremely rare in the region. The combination of different proxies is of importance for the understanding of how land use and natural variability interact, and a historical understanding of these interactions provides us with an ecological understanding of landscape dynamics, many of which are historically constituted.
The aim of this paper is to discuss the long-term changes in land use and resource utilisation over time in the Chibuene area of Mozambique and their interrelation with environmental change. We will focus on the period from a.d. 400–1900. Archaeological excavations in the Chibuene area have been carried out since 1978 (Sinclair 1982, 1987, 1997; Sinclair and Ekblom 2004) and sampling for botanical and osteological material has been an integral part of all these excavations (Ekblom 2004; Badenhorst et al. 2011). At the same time, interviews with local people on present day resource use have been carried out (Berger 2004), together with detailed surveys of present day vegetation (Ekblom 2004). Palaeoecological studies presented in Ekblom (2008) and Ekblom and Stabell (2008) have been important for an overall understanding of landscape transformation and climate-vegetation dynamics. Unfortunately, few seeds or other archaeobotanical macroremains have been encountered during the excavations. Owing to this scarcity, our understanding of land use and plant resource utilisation in Chibuene is still limited. This paper therefore contributes with an analysis of wood charcoal fragments sampled from archaeological contexts and complementary palaeoecological analyses of fungal spores and possible cereal pollen grains that elucidate the link between land use, use of resources and environmental change.
The analyses of the charcoal assemblages, originating from the 1995 and 1999–2000 excavations, supply us with an understanding of human agency and changes in land use practices as both a cause of and a response to environmental change. Charcoal analysis (anthracology) yields complementary information on the local woody vegetation composition, as a high proportion of wood for fuel and other purposes is collected near a settlement (cf. Shackleton and Prins 1992; Eichhorn 2002, 2007; Marston 2009; Gelabert et al. 2011). Furthermore, firewood is actively selected and brought to the living site; thus, it is a reflection of land use practices or of cultural/individual preferences. Positive examples from southern Africa are Deacon et al. (1983), Dowson (1988), Tusenius (1989), February (1992), Esterhuysen and Mitchell (1996), Cartwright and Parkington (1997), Cowling et al. (1999), Eichhorn (2002, 2007), Eichhorn and Jürgens (2003) and Vogelsang and Eichhorn (2011). In East Africa, Schmidt (1994, 1997) and Thompson and Young (1999) have shown the potential of wood charcoal analyses, and in West Africa a number of studies have been carried out elucidating Holocene climatic change and human alteration of woody vegetation (Höhn 2007; Neumann et al. 1998, 2009; Neumann 1999; Höhn and Neumann 2012; Eichhorn 2012).
Spores of coprophilous fungi provide a potential source of information on past herbivore abundance and, by inference, grazing by domestic animals (Van Geel et al. 2003; Graf and Chmura 2006; Marinova and Atanassova 2006; Van Geel and Aptroot 2006; Raper and Bush 2009). Fungal spores have not yet been widely used in palaeoecological or archaeological contexts in Africa (see however Burney et al. 2003; Carrion et al. 2000; Lejju et al. 2005; Ekblom and Gillson 2010a), but the potential of spore analysis and especially of spores from coprophilous fungi for understanding land use and resource utilisation is shown here.
Introducing the Chibuene landscape
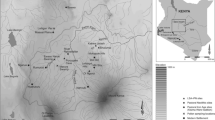
Chibuene is situated on the mainland side of Vilanculos bay, 7 km south of the town of Vilanculos (Fig. 1). Today it designates the name of a village and a developing resort. It is also the location of the archaeological site of Chibuene, dated from c. a.d. 600 onwards. The archaeological material suggests that this area has been more or less continuously occupied during the past 1,200 years, so it is ideal for discussing resource utilisation over time. The long occupation of this area can be explained by the rich availability of resources. Vilanculos bay provides plenty of marine resources, an important part of human diet over time. The lakes Nhaucati and Xiroche provide fresh water, and have probably retained water over most of the time during which this area was occupied.

Location of the research area (left) and the Chibuene locality (right) with excavation areas a and b marked
The present day landscape
The current vegetation in the Chibuene area is dominated by open grasslands with Eragrostis ciliaris, Panicum maximum, Cyperus spp., Cenchrus incertus and Digitaria spp. Representatives of the miombo woodland occur as smaller stands of Julbernardia globiflora and Strychnos spinosa. Within 500 m of the sea, the vegetation is characterised by dense shrub with Commiphora zanzibarica, Phyllanthus reticulatus and Turraea nilotica occurring together with Grewia monticola, Deinbollia oblongifolia, Clerodendrum glabrum and Acalypha glabrata. These species occur with climbers, such as Cocculus hirta and various herbs occurring in low frequencies (Ekblom 2004). The larger region consists of a miombo woodland savanna, where “Julbernardia globiflora and Brachystegia spiciformis are reported to be the main constituent but Brachystegia spiciformis is actually very rare in the region” (Wild and Fernandes 1968). Palaeoecological studies have shown that the vegetation before c. a.d. 1700 was considerably more wooded than today, consisting of a savanna-forest mosaic (Ekblom 2008). At present, the area is characterised by weakly developed soils with a low organic content and moderate weathering (Fränzle 1984, p. 85). The 1977–1992 civil war and the uncontrolled hunting which took place during this period of large-scale civil unrest are thought to have critically reduced the already stressed mammal populations in the region. Currently there are few large mammals in the area (Telford and de Castro 2001).
The local economy is based on farming, fishing and wage labour. The more common food crops are maize, groundnut, cassava and pearl millet together with other crops such as sorghum and sweet potato. Goats and chickens are common, but cattle are rare in the area. The collection of wild plants, fruits and berries is also an important part of everyday diets. Homesteads are continuously moved, as new fields are cleared and old plots are abandoned (Berger 2004).
Vilanculos usually receives most of its rainfall (832 mm per year) between December and February, when temperatures are the highest (27 °C). Winter temperatures average c. 19 °C. Yearly average evapotranspiration is of a magnitude of 1,440 mm (FAO 1984). In ideal situations, such annual averages meet the requirements of a number of different crops, including maize, which requires high average rainfall. However, yearly averages are less important for agroecology than interannual variability, and the greatest challenge for farmers is the unpredictability of the beginning and end of the rainy season (FAO 2004). During the 20th century, the summer rainfall region has experienced persistent cycles of wet and dry phases with a near decadal cyclicity, a pattern which is thought to have existed during the last 6,000 years (Tyson and Preston-Whyte 2000, pp. 332–335; FAO 1984). In addition, the coupled oceanic and atmospheric phenomenon of the ENSO (El Niño-Southern Oscillation), which occurs with a cyclicity of 3–5 years, is estimated to account for about 25 % of rainfall variation (Hulme 1995).
Presentation of the archaeological site and a brief history of the area
The main part of the archaeological site (Figs. 1, 2, area a), an area of c. 10 ha, is marked by numerous large Adansonia digitata (baobab) trees and a relatively dense scrub. However, there are also satellites of the site in the larger area, for instance near lake Nhaucati, where palaeoecological studies have been carried out (Figs. 1, 2, area b). The site is divided into an early and a late occupation phase. Dates associated with the early occupation phase range from c. a.d. 600 to 1300–1400 and they cluster between a.d. 700 and 1000. This early occupation phase has tentatively been subdivided into two facies (c. a.d. 600–900 and 900–1200). This is an arbitrary division, as layers representative of the early occupation phase carry their main characteristics from natural soil formation processes. However, we can assume the relative age of these layers based on stratigraphy. The upper occupation phase dates from a.d. 1300–1400 to 1650–1700 and is clearly distinguishable on the basis of a change in the ceramics typology (Sinclair and Ekblom 2004; Ekblom 2004). It is not clear whether the site was continuously occupied from a.d. 1700 until the present, as 14C dates from this period are few and the calibration is uncertain for the last two centuries.

Detail of the Chibuene site and the location of trenches and test pits in excavation area a and b as shown in Fig 1
Chibuene has revealed the presence of early style pottery (Matola) associated with early farming communities, which may suggest a longer occupation than indicated by the 14C dates. Chibuene also has a large number of imported glass and glass beads, and the town has been identified as a port of entry for the glass beads found in the interior at the end of the first millennium (Sinclair 1982, 1987, 1995; Morais 1988; Wood 2000, 2012; Huffman 2009). Chibuene acted as a regional centre of trade during a.d. 700–1000. It has been suggested that occupation in Chibuene was discontinuous after this, and there are few later imports (Wood 2012; Sinclair et al. 2012). In the 12th century a.d., Chibuene probably came under tributary rule from Manyikeni, a Zimbabwian tradition style of stone walled settlement situated 10 km west of Chibuene (Barker 1978; Sinclair 1987). Later written documents suggest that the lineage of Sono of Manyikeni controlled the coastal area (Liesegang 1990). The pottery of the later occupation in Chibuene also shows great similarities with the Manyikeni pottery, indicating that the two settlements were in contact. The presence of marine shell and imports from the Indian Ocean trade in Manyikeni also confirms that the coast and the hinterland were connected through trade networks. In the 18th century, the Vilanculo chieftainship was established in the Vilanculo area with the movements of the Tsonga. The Vilanculo chieftainship claimed authority over the Chibuene area. The clashes between the Nguni and the Chibuene ancestors in the 19th century are a recurring theme in oral tradition, and written documents suggest that the social unrest in this period had important social and economic implications (Ekblom 2004, pp. 51–52). Migrant labour has been part of local economy since the end of the 19th century, providing household income and bride wealth. Migrant labour was also encouraged by the colonial authorities, as a monetary hut tax was demanded (Ekblom 2004, p. 64). At present, Chibuene is in a state of very rapid transformation, as a large number of land plots have been concessioned to private investors. As a result, many residents have recently moved their homesteads further inland.
Methodology
Microscopic fossil identification
The methodology of the pollen analysis from the Chibuene area has been presented in detail by Ekblom (2008). Pollen, spore and microscopic charcoal preparations were carried out through digestion of cellulose with NaOH and concentration of pollen through acetolysis. Lycopodium spores were added for an estimate of pollen concentration (Stockmarr 1971; Moore et al. 1991). A percentage summary diagram of the main pollen groups is shown here. This is based on the arboreal pollen, terrestrial herbs and grasses (see Table 1 for ecological groupings). Aquatic and limnic taxa (such as Cyperaceae) are not shown here. The arboreal pollen taxa have also been arranged in general ecological groupings such as riparian-forest, generalist and savanna environment, and the percentage distribution shown here is based on the arboreal pollen sum of 250. Total pollen counts ranged between 1,000 and 15,000 pollen grains.
The counting of microscopic charcoal particles was carried out over the slides until 300 pollen grains (aquatic/terrestrial) had been encountered. Size groupings have been made based on the longest axis of the charcoal particle. Here we separate two size classes, 10–100 μm interpreted as linked with local or regional fires and 100 to <1,000 μm interpreted as linked with local fires (see review in Pitkänen et al. 1999). For the larger size classes above 200 μm, all fragments encountered on the slides were recorded. The results are repeated here for comparison with the dung fungal spore data.
Spores were counted in relation to the added Lycopodium spores and are presented as concentrations. The references used were Jarzen and Elsik (1986), Van Geel et al. (1983, 1986, 2003), Van Geel and Aptroot (2006), Graf and Chmura (2006), Prager et al. (2006). Most types presented here have only been provisionally identified or grouped together into main type groups. Types that occur infrequently or in very low numbers are displayed as ‘ungrouped’. Spore types that could not be identified to a specific taxon were grouped according to the classification suggested by Jarzen and Elsik (1986), where spore types are given a form-generic name, for example Dicellaesporites.
As well as the spore assemblage identification, an attempt was made to separate possible domesticated cereal grasses from the wild grasses. The pollen of indigenous African cereal grasses (Pennisetum americanum, Sorghum bicolor, Eleusine coracana) overlap in size and are morphologically similar to wild grass pollen (Tomlinson 1973). It has been noted in pollen analyses from the Mozambique interior (Ekblom and Gillson 2010b) that larger grass pollen >40 μm tends to occur in larger numbers together with maize pollen; thus, it is likely that the grass pollen larger than 40 μm represents indigenous cereal grasses. We have therefore used a tentative size separation of grasses <40 μm here. As glycerol jelly tends to swell pollen grains by 10–15 %, pollen sizes have been quantified based on comparison with Lycopodium spores, which was conducted during the slide scanning for the spore analysis.
Macroscopic fossil identification
Macroscopic charcoal from archaeological contexts such as hearths, postholes and pits were retrieved through flotation in the field and retained on a 0.25 mm mesh. All samples were analysed using a stereomicroscope and screened for seeds and identifiable plant remains as a first step. In a second step, all charcoal fragments ≥2.5 mm were picked out from the samples. These were, on the one hand large enough to be split for the microscopic analyses and, on the other hand, displaying all relevant wood anatomical features.
Charcoal analysis was carried out using incident light microscopy with a Leica Laborlux S and Leica DM 4000, and with a Hitachi 4500 scanning electron microscope. Prior to analysis, each charcoal fragment was split along the three diagnostic planes (transversal, radial, and tangential). Wood anatomical characters relevant for determination were selected according to the standards of the “International Association of Wood Anatomists” (Wheeler et al. 1989). As no comprehensive wood anatomical atlas or key is available for the research area, we depended on published keys, databases and atlases for the Sahara, for western, southern and central Africa (Lebacq 1955–1957; Normand 1950–1960; Prior and Gasson 1990; Neumann et al. 2001; Eichhorn 2002; Höhn 2005). Miscellaneous data for the research area proper (Ferreirinha 1962; Barreto 1967; De Freitas 1986), the “Inside Wood” internet database (InsideWood 2004 onwards) and the collection of wood microscope slides from West and Central Africa housed at the Institute of Archaeological Sciences at Goethe University, Frankfurt, were also consulted. A limitation of the study is that identification to species level is rarely possible, partly owing to limited references for the regional woody flora, but mainly because of wood anatomical similarities within genera or families. Therefore, the “type” concept is used here, according to which a type is designated to a morphological (or in our case anatomical) category, distinguished either by one distinct character or a unique combination of characters. A type may represent one single or a number of species (cf. Punt et al. 2003). In order to narrow the identification down to the tree and shrub species possibly used in Chibuene over time, we rely on published data on the present day regional woody flora and vegetation (Coates Palgrave 2002; Hyde et al. 2002), supported by the understanding of landscape changes available from the pollen analyses.
Wood anatomical descriptions of the identified charcoal types are given in Appendix in Supplementary material. The most common and the most ecologically significant charcoal taxa were documented by micrographs taken with a Leica DFC 295 digital camera, using “multifocus” imagery. This method does not reach the same depth of focus as the SEM photographs often used in charcoal publications. Nevertheless, it yields an optical impression highly comparable to the image seen during charcoal identification under incident light and allows for illustration of most anatomical characters relevant for identification. For the illustration of minute characters such as pit vesturing, SEM imagery was used in addition.
The focus of macroscopic charcoal analysis at Chibuene was to detect changes in the selection of wood over time that resulted from corresponding changes in the surrounding vegetation. The charcoal data is presented based on a division of the site chronology into the three occupational periods. In many sampled archaeological contexts, charcoal fragments were too small for identification; thus, the number of analysed contexts is relatively low. This is especially the case for the late occupation phase and may have led to a bias in taxa representation. Sizes of the individual samples were often small and the number of identifiable charcoal fragments highly variable between the samples (between 1 and 108 fragments ≥2.5 mm/sample). In order to obtain as representative charcoal assemblages as possible for each of the three occupation periods, we analysed all available fragments of this size class.
Results
Dung fungal spore and microscopic charcoal analyses
Lake Nhaucati
The diagram has been divided into four phases, based on the percentage distribution of the main ecological groups of pollen, concentration of fungal spores and estimation of charcoal (Fig. 3). The first phase (a.d. 400–900) shows low concentrations of spores. Charcoal is represented, but in low numbers. The pollen record is dominated by grasses and the arboreal pollen taxa, mostly represented by riparian and generalist taxa.

Simplified pollen and spore diagram of the sequence from lake Nhaucati. The summary pollen diagram includes (from the left) arboreal pollen, grasses, herbs and ungrouped including unidentified taxa. The curve of total pollen concentration consists of arboreal pollen (dark grey) and grasses (light grey) together with possible cereals (black). Black bars show charcoal representation (in relation to pollen grains), the black silhouettes show concentration of moss/fern spores and fungal spores from coprophilous taxa. The grey silhouette to the far right shows the concentration of coprophilous spore taxa (dark grey) in relation to other spore taxa (light grey)
In the a.d. 900–1350 early-late occupation phase, sedimentation rates are very high, which has been suggested to be linked with high lake levels and high rainfall (Ekblom and Stabell 2008). Pollen and especially spore concentrations increase significantly in this phase. Coprophilous spores are well represented by c. 40 % of total spores and are dominated by Sordariaceae and Coniochaeta lignaria, and a few Sporormiella also occur. In the period a.d. 900–1350, there is also an increase in arboreal pollen and both savanna and riparian pollen types. Microscopic charcoal particles remain well represented until c. a.d. 1300, when there is also a decline in locally transported microscopic particles.
Phase 3, a.d. 1400–1600 late occupation phase shows a progressive decline in riparian taxa and possible cereal pollen is no longer represented. Overall, spore concentrations are very low and are mostly represented by monolete spores, although Coniochaeta cf. lignaria is present in very low numbers. Locally transported charcoal is not well represented compared to the previous phase.
In Phase 4, a.d. 1600 to present, late occupation phase, the contribution of riparian taxa to the arboreal pollen sum is less than 10 %, while savanna and generalist taxa increase. Coniochaeta cf lignaria and Sordariaceae show a small increase in the uppermost part of the sequence.
Lake Xiroche
Spores are not well represented in the lake Xiroche sequence and the total spore concentration is low (Fig. 4). Nevertheless, its spore assemblage sequence provides a comparison to lake Nhaucati. Arboreal pollen is well represented in the very beginning of this phase c. a.d. 700, (early occupation phase) but declines in the upper part of the phase. Smaller charcoal particles occur but charcoal is generally not well represented.

Simplified pollen and spore diagram of the sequence from lake Xiroche. For further explanations see Fig. 3. Note that the peak in riparian taxa c. a.d. 600 is owing to a local increase of Canthium-type
From a.d. 950, Phase 2, early-late occupation phase, there is an increase in spore concentration and Sordariaceae and cf. Coniochaeta are well represented. Larger particles of microscopic charcoal, which can be associated with local fires, are well represented and possible cereal pollen grains appear at the same time. Savanna type taxa dominate the arboreal pollen signal. The overall amount of charcoal is very high between a.d. 1100 and 1200. The amount of possible cereal pollen grains peaks from 1200 a.d.
Phase 3 (a.d. 1400 to present) is marked by the decline in riparian taxa in favour of savanna type taxa. Larger charcoal particles are fewer than before. The total spore concentration is higher in this phase than earlier, and it is dominated by non-coprophilous spores. An increase in coprophilous spore types, mainly Sordariaceae, can be seen in the top of the sequence where there is also an increase in charcoal.
Macrofossil and wood charcoal analyses
Only a few carbonised seeds were found, including four cereal grains (Sorghum and Eleusine) associated with the upper facies of the early occupation phase and the late occupation phase. Chenopodiaceae/Amaranthaceae, Caryophyllaceae and Grewia seeds were found in layers associated with the early occupation (Ekblom 2004). As this material category was so sparse, it will not be further discussed in this paper. Instead we will focus on the wood charcoal analyses.
Early occupation phase
The charcoal spectra of both early and later facies of the early occupation phase at Chibuene (Table 2) are clearly dominated by Sapotaceae in association with Androstachys johnsonii-type (Fig. 5a–c) and Dialium (Fabaceae-Caesalpinioideae; Fig. 5d–f). The genus Dialium is represented by two species in Mozambique, D. schlechteri and D. holtzii, but only D. schlechteri occurs in the research area proper (Coates Palgrave 2002; Hyde et al. 2002, Table 3). In southern Africa in general, this species is confined to coastal forest. In the Inhambane province of Mozambique it has been recorded as one of the dominant species of rare semi-deciduous forest patches (Wild and Fernandes 1968). Most other members of the Fabaceae-Caesalpinioideae are much fewer than in the late occupation phase. The evergreen species Androstachys johnsonii, (Picrodendraceae) occurs at present in Mozambique most frequently in forest stands on sandy soil, in a mosaic with the Julbernardia globiflora savanna (Alvin 1987).

SEM (a–c, e–f) and incident light multifocus (d) images of wood charcoal from Chibuene: a Androstachys johnsonii-type transversal view, exclusively solitary vessels; b A. johnsonii-type tangential view, exclusively uniseriate rays; c A. johnsonii-type radial view, homocellular rays and vessel-ray pits similar to intervessel pits; d Dialium schlechteri-type, transversal view, banded axial parenchyma; e D. schlechteri-type transversal view, banded axial parenchyma; f D. schlechteri-type transversal view, arrow silica body in axial parenchyma cell
The identified Sapotaceae charcoal fragments can hardly be assigned to particular genera or species. Unfortunately, the Sapotaceae genera represented in the area display little anatomical variation (mainly restricted to the presence/absence, the size and the location of calcium oxalate crystals and silica) which does not allow for safe separation at a low taxonomic level. In most of the identified fragments, neither crystals nor silica were observed. We distinguished two types, the Sideroxylon/Chrysophyllum-type (no silica observed) and the Mimusops-type (large silica bodies present, Fig. 6a–c). Nevertheless, each of those types may represent several species.

SEM images of wood charcoal from Chibuene a–c Mimusops-type: a transversal view, vessels in radial rows, apotracheal diffuse and banded parenchyma; b radial view: thin-walled tyloses, heterocellular rays; c transversal view, large silica body in parenchyma cell (globular decorated type); d–f Phyllanthus-type; d transversal view, numerous vessels in radial rows; e tangential view, intervessel pits; f vessel-ray pits
The Phyllanthus-type (Fig. 6d–f) probably represents P. reticulatus, a species of riverine thicket and coastal scrub, whereas Bruguiera gymnorrhiza (Fig. 7a–c), the large-leafed mangrove, indicates the nearby presence of mangrove vegetation. It grows on the seaward side of mangrove swamps (Coates Palgrave 2002). The few other taxa represented during this phase have a wide ecological spectrum and thus provide little palaeoecological information, for example Vitex/Bridelia, Ochna (Fig. 7d–f), Cordia and Alchornea laxiflora.

SEM images of wood charcoal from Chibuene: a–c Bruguiera gymnorrhiza-type; a transversal view, vessels in radial rows; b tangential view, wide and high rays, scalariform intervessel pits; c radial view, damaged scalariform perforations; d–f Ochna-type; d transversal view, exclusively solitary vessels and wide rays with crystal idioblasts; e tangential view, wide and high rays with idioblasts; f radial view, heterocellular ray with idioblasts and vessel-ray pitting with reduced borders
Late occupation phase
The charcoal spectra differ distinctly from those of the early occupation phase. The over-all dominant taxa are legumes of the Caesalpinioideae subfamily (Tamarindus indica/Cassia-type). T. indica, the tamarind tree and Cassia species are very close in wood anatomy. Although Cassia has generally more paratracheal axial parenchyma than Tamarindus, the distinction of both taxa in charcoal assemblages is not always secure due to feature overlap according to the wood anatomical descriptions in Neumann et al. (2001) and Appendix in Supplementary material. However, both taxa indicate the presence of open woodland or potentially of agricultural parkland and fallow land. This clearly points to vegetation alteration and the opening of vegetation compared to the preceding settlement phases. Cassia is represented in the Chibuene area by C. abbreviata. Tamarindus indica has been observed today, although it was not recorded by Massinga (Cited in Ekblom 2004). As a valuable tree for consumption and use, it may have been more common during the late occupation of Chibuene. Sapindaceae, possibly Deinbollia or Dodonea was tentatively identified, indicating the persistence of coastal shrub taxa in the late occupation phase. This is not surprising, as coastal scrub still occurs at present on the seawards facing slope of the site.
Bones and shellfish
The results of osteological analyses and of the shellfish assemblage have been reviewed by Badenhorst et al. (2011), but a short summary is presented here as it provides a context to the changes in the charcoal and pollen assemblages.
The bone assemblage
Of the bones excavated between 1995 and 2001, only those from the 1995 field season have been analysed in detail (Badenhorst et al. 2011). Even though this faunal assemblage is small, we can make some tentative statements about the differential representation of taxa over the three periods. The weight of bone specimens suggests a higher contribution of marine resources in the early occupation phase. The marine contribution during the second facies of the early occupation phase is higher in its late facies, and in the late occupation phase. Cattle and caprines (sheep and goats) are present throughout the deposits, but cattle are slightly more common in the early occupation phase. Caprines occur in more even numbers. Few wild animals are present in the sample, and no inferences can be made about their palaeoecology.
Shellfish
The 3 m high shell midden was analysed in detail by Sinclair with technical support from the Museum of Natural History, Maputo (Ekblom 2004). Increased use of shellfish has been observed for the late occupation phase (Sinclair, in Ekblom 2004). Oysters in the large shell midden and other parts of the site get progressively smaller over time. This indicates that they were gradually being fished out and the wider range of shellfish in the late occupation phase may perhaps be explained in this light.
Discussion
Pollen and spore analyses
The pollen diagrams of both lake Nhaucati and Xiroche show extensive changes in vegetation cover in Chibuene over the last 1,600 years, as discussed in detail in Ekblom (2008). From c. a.d. 400, around the time when the first farmers were supposedly settling the area, the Chibuene landscape was covered by a mosaic of savanna and riverine forests. Pollen from savanna and savanna woodland type trees, particularly Brachystegia, is well represented at this time as well as pollen types associated with forests and riverine forests such as Moraceae, Dialium-type, Trema and Celtis (Figs. 3, 4; Table 3). The latter are all taxa that are not found in the Vilanculos region today. Fluctuations in the relative distribution of savanna, forest and generalist ecology trees can be seen over time.
Microscopic charcoal is well represented in both the lake Xiroche and Nhaucati sequences from a.d. 900 and 1100 respectively. The representation of spores of coprophilous fungi is also high in both cores from the same period. At the same time, the concentrations of possible cereal pollen grains increase in both sequences and show their highest values between a.d. 1200 and 1300. The increase in pollen concentration here is owing to low erosion of sediments and a higher accumulation of organic material due to the higher lake levels, in turn linked to a wetter climate. However, in lake Xiroche, pollen concentrations remain moderate. Thus, we argue that the increase in coprophilous spores and possible cereal grains is a reflection of landscape changes and is not merely an artefact of differential preservation or sediment accumulation in the records. The combination of local microscopic charcoal and cereal pollen grains is a strong indicator of farming activities; therefore, we assume that much of the dung fungal spore record is related to domestic animals. Both cattle and sheep/goat are represented in the bone material, although it is impossible to quantify how large these populations were on the basis of the osteological material (Badenhorst et al. 2011).
Despite evidence of the use of fire, presumably from clearances of fields, there are no indications of linked vegetation changes such as a decline in forest taxa in this period. However, the relative representation of pollen taxa is highly variable, which may perhaps be associated with shifting agriculture and successive phases of clearing and regrowth. See Ekblom et al. (2011) for a similar discussion on pollen analysis in the lower Limpopo valley.
Forest pollen types are well represented at both sites until a.d. 1400–1600, when there is a decline of pollen types associated with forests and riverine forests (Moraceae, Celtis, Trema). The decrease in forest types is particularly clear if grass pollen is excluded from the pollen sum, as was done in Ekblom (2008). Instead, other taxa in the generalist group such as Alchornea, Spirostachys and Brachylaena become common, particularly in the lake Xiroche diagram, which is a smaller basin and thus would reflect a more local signal (Jacobsen and Bradshaw 1981). There is also a concurrent increase of types associated with open savanna and savanna woodland such as Brachystegia, Dodonaea and Sclerocarya. This is probably owing to one or several hiatuses in the sediment record caused by the drying out of the lakes associated with the extreme droughts that took place between c. a.d. 1600 and 1800 and around a.d. 1700 in particular (Ekblom and Stabell 2008). There are no corresponding increases in local charcoal or possible cereal grains to suggest that the decline of forest taxa was linked with clearings or intensification of farming at this time. Rather, the agricultural signal is significantly lower than before. The decline of forest taxa is less marked in lake Nhaucati than in lake Xiroche, which may indicate that forest taxa continued to be present in the larger region, but not locally. Lake Nhauhache, located 6 km northwest from Chibuene, did retain water in this time period, as shown by diatom and sedimentary analyses (Holmgren et al. 2012).
Local changes in vegetation as suggested by the macroscopic charcoal data
The identifications of the charcoal assemblages are biased, particularly in the late occupation phase, as few contexts contained sufficient amounts of identifiable fragments. Furthermore, the chronostratigraphical resolution is low; stratigraphies have been modified by secondary processes and few of the analysed contexts have been dated. Despite this, the charcoal assemblages of Chibuene allow for inferences on wood fuel selection and collection, particularly between the early and the late occupation phase and therefore provide an important parallel to the pollen analyses.
During the entire early occupation phase, wood was probably mainly brought into the settlement from the forest communities. This is indicated by the dominance of Sapotaceae, mainly Sideroxylon/Chrysophyllum-type, and Androstachys johnsonii-type as well as the presence of Dialium schlechteri. Dialium-type pollen was common in the lower part of the pollen diagram, particularly in lake Nhaucati (Ekblom 2008). Of the Sapotaceae genera, only Mimusops has been positively identified in the pollen record. Other Sapotaceae types have been grouped as unidentified in the pollen analyses. A number of Sapotaceae species potentially occur in southeastern Mozambique (Flora of Mozambique Zone GI (Hyde et al. 2002, Table 2). Today Sapotaceae are rare in the Chibuene area proper (Ekblom 2004, pp. 22–25), but the southern Mozambique vegetation map (Wild and Fernandes 1968) shows a littoral dune vegetation unit characterised by Mimusops caffra and semi-deciduous forest strips close to the coast with Sideroxylon. Androstachys johnsonii, which at present occurs in forest patches, is not represented in the pollen spectra. Phyllanthus-type was also a common wood fuel in the early occupation phase. Phyllanthus-type pollen (Ekblom 2004) was included in the Fagara-type pollen group (Ekblom 2008) and is grouped within the generalist pollen group as it may include a number of different taxa. Phyllanthus-type pollen is common in the pollen diagrams throughout time in lake Xiroche, but declines in lake Nhaucati after c. a.d. 1300. The most likely candidate for the Phyllanthus-type wood fuel is P. reticulatus, and today it is found in the coastal scrub present on the seaward facing slope of Chibuene (Ekblom 2004). In the past, it may also have been a constituent of the forest vegetation nearer to the lakes. In the second facies of the early occupation phase, this type occurs less frequently in the charcoal assemblage than in the first facies, whereas Vitex/Bridelia-type increases at the same time.
The dominant charcoal taxa during the late occupation phase are legumes of the Fabaceae-Caesalpinioideae (Tamarindus indica/Cassia-type). Cassia pollen was identified and is here included in the savanna pollen group. Brachystegia/Julbernardia-type charcoal, which can be separated from the Tamarindus indica/Cassia-type due to its exclusively uniseriate rays, is only present in the early occupation phase charcoal assemblages, but always in low numbers. It represents Julbernardia globiflora and Brachystegia spiciformis (also Fabaceae-Caesalpinioideae). It is, however, clear from the pollen analyses that Brachystegia, which forms the miombo woodland community together with Julbernardia, was present in the area already from a.d. 400, but in much lower numbers than at present.
Conclusion: landscape changes and changing patterns of resource use
The Chibuene landscape has experienced four phases of land use and resource use that have interacted with changes in the environment.
Phase 1 (a.d. 400–900): Forest savanna mosaic, low intensity cattle herding and cultivation, trade of resources for domestic use.
Phase 2 (a.d. 900–1400): Forest savanna mosaic and high intensity or extensive cultivation and cattle herding.
Phase 3 (a.d. 1400–1800): Savanna woodland and progressive decrease in forests due to droughts, decline of agricultural activities and higher reliance on marine resources, possible trade in resources with the interior.
Phase 4 (a.d. 1800–1900): Open savanna with few woodland patches, warfare and social unrest. Collapse of trade with the interior. Decline in marine resources and wildlife, loss of cattle herds. Expansion of agriculture locally, introduction of new world crops and clearing of Brachystegia.
The first farmers who came to the Chibuene area between a.d. 400 and 700 settled a landscape covered by a mosaic of savanna and riverine or coastal forests. Forests were well represented in the area surrounding the freshwater lakes and in the immediate seaward area. Clearings for farming were small and the natural savanna patches were probably used for farming. There is no evidence of significant clearances of vegetation in the period a.d. 600–1000 locally in the lake Xiroche case, or more regionally in the lake Nhaucati case, and the amount of microscopic charcoal is low in both cores. The archaeological material, local ceramics and trade goods, suggest that Chibuene was a busy node of trade between the coast and the interior of southern Africa at this time. The low agricultural signal in the pollen diagrams must therefore be explained. The animal bone suggests that cattle were more common in the early occupation phase, specifically in the early facies (a.d. 600–900) (Badenhorst et al. 2011). It is therefore possible that cattle herding was more important for the local subsistence and economy at this time and that grain was traded from the larger area. The low agricultural signal thus underlines the importance of Chibuene as a place of trade.
From c. a.d. 950–1000, the archaeological material suggests a change in the role of Chibuene. Long-distance imports of glass beads are no longer part of the archaeological material (Sinclair et al. 2012; Wood 2012). Sinclair (1987) suggested a break in occupation around this time, mainly on the basis of the complete change in ceramic style between the early and late occupations. Somewhat unexpectedly, the pollen diagrams from this time suggest an increase in agricultural activity. There is a significant increase in microscopic charcoal at this time in both lake Nhaucati and Xiroche. There is also a higher concentration of coprophilous spores in both lakes, and from a.d. 1100, possible cereal grass pollen. Above we have argued that this is not merely a reflection of better preservation or faster accumulation of sediments, but a reflection of changes in land use. From this, we can conclude that although the settlement in the Chibuene area may have been discontinued, agricultural activities in this area expanded. Possibly, the change in trade patterns caused a complete transformation in the economy of Chibuene that moved the focus primarily from exchange and trade, so therefore farming and herding activities were intensified or expanded. Despite evidence of expansions of farming, forests were as common in the area as before. There is no evidence of decline of forest taxa during the phase of agricultural intensification that lasted until a.d. 1400. Possibly settlements were also more dispersed in the larger region in this period.
From a.d. 1400, the pollen diagrams show a decrease in agricultural activities, with a decline in charcoal values, coprophilous spore concentrations and cereal pollen. Meanwhile, typical forest-type pollen taxa decline. The extension of forests during the early occupation phase and their subsequent decline in the late occupation phase shown in the pollen analyses are supported by the selection of wood fuel by the Chibuene inhabitants. Although patches of savanna and savanna type trees were present in the area, the forest-type taxa were mainly selected for fuel and other purposes. This was probably a matter of choice. Some of the taxa, such as Androstachys johnsonii, also called the “Lebombo Ironwood” because of its hard, dense and durable wood, yield highly valuable firewood and wood for construction purposes (Coates Palgrave 2002; Lemmens 2011). In the late occupation phase, however, savanna trees were mainly used. This was probably not for reasons of preference, but a reflection of availability, as forest was significantly reduced in this period.
The decline of forests has earlier been related to the onset of droughts in the summer rainfall region related to a climatic cooling (Ekblom 2008). Lee-Thorp et al. (2001) and Holmgren et al. (2003) date the onset of cool/dry conditions in the summer rainfall region from a.d. 1600, and lasting until 1800. They found that several decades of cool–dry conditions took place from around a.d. 1100, with shorter cool–dry events at around 1350, 1450 and 1500. In the lower Limpopo valley, the effect of cool/dry conditions can be seen from a.d. 1450 onwards as a reduction of riparian forests (Ekblom et al. 2011). Thus, we still argue that the decline of forest was most probably driven by climate change rather than human activities. We would have expected earlier signs of forest reductions, if this last had been the case, and agricultural activities seem to have declined at the same time as the forest decline occurred. Again, this is most probably linked to the droughts that caused a great stress on the natural vegetation, as well as on farming communities. However, although rainfall increased after a.d. 1800, forests did not spread again in the area, which may be owing to a combination of poor soils, the establishment of grasslands in the formerly forested areas and farming.
The decline in agriculture probably led to a higher reliance on marine resources. In the late occupation phase, there was also an aggregation of population in Chibuene. This may be connected with the higher reliance on marine resources. Cattle are less common in the late occupation phase and the contribution of wild animals remains low throughout the early and late occupation phases. Wild resources were probably traded with the interior at this time and written documents from the 19th century relate how animal skins were brought from the interior. In the late occupation phase, the shellfish assemblage suggests a diversification in the use of resources with a wider range of taxa collected than previously. The exploitation of shellfish and pearl fishing in the Bazaruto area is described in written sources (Duarte Barbosa 16th century, in Theal 1964, p. 93). The economic importance of the shellfish trade between India and Bazaruto is also mentioned in written sources dating to a famine in 1893, when shellfish were exchanged for grain (Rita-Ferreira 1999, p. 15). The decreasing size of oyster shells in the upper part of a large shell midden in Chibuene, dating to the late occupation phase, suggests that they were gradually being fished out. This should perhaps be best explained as a result of the increased reliance on marine resources caused by the droughts.
Agricultural activities naturally continued in the area but on a smaller scale than at a.d. 950–1400. Meanwhile cattle herding was discontinued, and today no cattle can be seen in the area (Berger 2004). However, the 20th century probably saw a re-expansion of agricultural activities. Ekblom (2004) has suggested that the combination of repeated droughts and later warfare in the 17th–19th centuries constituted a socio-ecological crisis. By the beginning of the 20th century, old trade routes with the interior had been abandoned, wildlife was diminished and marine resources had declined, which increased the population’s dependence on local agricultural production. In the course of the 20th century, Brachystegia spiciformis disappeared from the area. Brachystegia is represented in the pollen diagram but it is not seen in the area today (Ekblom 2004, p. 24). At the same time, new world crops such as cassava, maize, and cashew became more popular and families became increasingly dependent on migrant labour as a source of income (Ekblom 2004).
In conclusion and as shown here, the combination of several different environmental sources, pollen analyses, macroscopic charcoal, bones and mollusc shells enables us to elucidate how land use and natural variability have interacted over time, even on the basis of very small assemblages. The macroscopic charcoal assemblage provides important information on local vegetation changes which is complementary to the pollen analyses of more regional value, and the archaeological data. Similarly, the supplementary analyses of microscopic dung fungal spores provide important indicators when interpreting the palaeoenvironmental record and we recommend that these sources should be used more frequently in the future.
References
Alvin KL (1987) Leaf anatomy of Androstachys johnsonii Prain and its functional significance. Ann Bot 59:579–591
Badenhorst S, Sinclair P, Ekblom A, Plug I (2011) Faunal remains from Chibuene, an Iron Age coastal trading station in central Mozambique. South Afr Humanit 23:1–15
Barker G (1978) Economic models for the Manekweni Zimbabwe, Mozambique. Azania 13:71–100
Barreto LS (1967) Madeiras de Moçambique. Estrutura do lenho (1a série). Revista dos estudos gerais universitarios de Moçambique, Lourenço Marques (sér. II) 4:59–183
Berger A (2004) Food resources at Manyikeni and Chibuene, two archaeological sites from southern Mozambique. In: Oestigaard T, Anfinset N, Saetersdal T (eds) Combining the past and the present: archaeological perspectives on society. (BAR, Int Ser 1210), Archaeopress, Oxford, p 149–159
Burney DA, Robinson GS, Pigott Burney L (2003) Sporormiella and the late Holocene extinctions in Madagascar. Proc Natl Acad Sci 19:10,800–10,805
Carrion JS, Scott L, Huffman T, Dreyer C (2000) Pollen analysis of Iron Age cow dung in southern Africa. Veget Hist Archaeobot 9:239–249
Cartwright C, Parkington J (1997) The wood charcoal assemblages from Eland’s Bay Cave, southwestern Cape: principles, procedures and preliminary interpretation. S Afr Archaeol Bull 52:59–72
Coates Palgrave K (2002) Trees of southern Africa, 3rd edn. Struik, Cape Town
Cowling RM, Cartwright CR, Parkington JE, Allsopp JC (1999) Fossil wood charcoal assemblages from Eland’s Bay Cave, South Africa: implications for Late Quaternary vegetation and climates in the winter rainfall Fynbos biome. J Biogeogr 26:367–378
De Freitas MCPG (1986) Madeiras de Moçambique. Characteristicas anatómicas, fisicas e mecânicas. Instituto de Investigação Científica Tropical, Lisboa
Deacon HJ, Scholtz A, Daitz LD (1983) Fossil charcoals as a source of palaeoecological information in the Fynbos region. In: Deacon HJ, Hendey B, Lambrechtts JJN (eds) Fynbos palaeoecology; a preliminary synthesis. South African National Scientific Programmes Unit, Pretoria, pp 174–182
Dowson TA (1988) Shifting vegetation zones in the Holocene and later Pleistocene: preliminary charcoal evidence from Jubilee shelter, Magaliesberg, Southern Transvaal. Palaeoecol Afr 19:233–239
Eichhorn B (2002) Anthrakologische Untersuchungen zur Vegetationsgeschichte des Kaokolandes, Nordwest-Namibia. Dissertation, University of Cologne
Eichhorn B (2007) Use of firewood resources in a hyperarid environment: charcoal analysis from sites in the Skeleton Coast Park, Northern Namib Desert. In: Cappers RTJ (ed) Fields of change. Progress in African archaeobotany. Groningen Institute of Archaeology, Groningen, pp 21–34
Eichhorn B (2012) Woody resource exploitation for iron metallurgy of the Fiko tradition: implications for the environmental history of the Dogon Country, Mali. In: Robion-Brunner C, Martinelli B (eds) Métallurgie du Fer et Sociétés Africaines. Bilans et nouveaux paradigmes dans la recherche anthropologique et archéologique (Cambridge Monographs in African Archaeology 81, BAR S 2395). Archaeopress/Hadrian Books, Oxford, p 139–148
Eichhorn B, Jürgens N (2003) The contribution of charcoal analysis to the late Pleistocene and Holocene vegetation history of north-western Namibia. In: Neumann K, Butler A, Kahlheber S (eds) Food, fuel and fields. Progress in African archaeobotany (Africa Praehistorica 15). Heinrich-Barth-Institut, Köln, p 151–162
Ekblom A (2004) Changing landscapes: an environmental history of Chibuene, southern Mozambique. Dissertation, Uppsala University
Ekblom A (2008) Forest-savanna dynamics in the coastal lowland of southern Mozambique, c. 400 AD–present. Holocene 18:1,247–1,257
Ekblom A, Gillson L (2010a) Using dung fungal as indicators of past herbivore abundance, Kruger and Limpopo National Park. Palaeogeogr Palaeoclim Palaeoecol 296:14–27
Ekblom A, Gillson L (2010b) Fire history and fire ecology of Northern Kruger (KNP) and Limpopo National Park (PNL), southern Africa. Holocene 20:1,063–1,077
Ekblom A, Stabell B (2008) Paleohydrology of Lake Nhaucati (southern Mozambique), c. 400 AD to present. J Paleohydrol 40:1,127–1,141
Ekblom A, Gillson L, Notelid M (2011) A historical ecology of Limpopo and Kruger National Parks and lower Limpopo valley. J Archaeol Anc Hist 1:1–29
Esterhuysen A, Mitchell PJ (1996) Palaeoenvironmental and archaeological implications of charcoal assemblages from Holocene sites in western Lesotho, southern Africa. Palaeoecol Afr 24:203–232
FAO (1984) Agroclimatological data for Africa, countries south of the equator. FAO, Rome
FAO (2004) Drought impact mitigation and prevention in the Limpopo river basin. A situation analysis. FAO, Harare
February E (1992) Archaeological charcoals as indicators of vegetation change and human fuel choice in the late Holocene at Eland’s Bay, western Cape Province, South Africa. J Archaeol Sci 19:347–354
Ferreirinha MP (1962) Madeiras de Angola—2.a série. Garcia de Orta 10:111–123
Fränzle O (1984) Afrika-Kartenwerk : Beiheft. Ser. S, Südafrika, Beih. zu Blatt 4, Bodenkunde - Südafrika (Mocambique, Swaziland, Republik Südafrika). Bodenkunde–Südafrika (23°10′–26°52′S, 29°50′–35°40′E). Borntraeger, Berlin
Gelabert LP, Asouti E, Marti EA (2011) The ethnoarchaeology of firewood management in the Fang villages of Equatorial Guinea, central Africa: implications for the interpretation of wood fuel remains from archaeological sites. J Anthropol Archaeol 30:375–384
Graf M, Chmura GL (2006) Development of modern analogues for natural, mowed and grazed grasslands using pollen assemblages and coprophilous fungi. Rev Palaeobot Palynol 141:139–149
Hall M (1981) Settlement patterns in the Iron Age of Zululand, an ecological interpretation. BAR Int Ser 119, Oxford
Höhn A (2005) Zur eisenzeitlichen Entwicklung der Kulturlandschaft im Sahel von Burkina Faso. Untersuchungen von archäologischen Holzkohlen. Dissertation, Frankfurt University
Höhn A (2007) Where did all the trees go? In: Cappers R (ed) Fields of change. Progress in African archaeobotany. Barkhuis Publishing, Eelde, Groningen, pp 35–42
Höhn A, Neumann K (2012) Shifting cultivation and the development of a cultural landscape during the Iron Age (0–1500 AD) in the northern Sahel of Burkina Faso, West Africa: insights from archaeological charcoal. Quat Int 249:72–83
Holmgren K, Lee-Thorp JA, Cooper GRJ, Lundblad K, Partridge TC, Scott L, Sithaldeen R, Siep Talma A, Tyson PD (2003) Persistent millennial scale climatic variability over the past 25,000 years in Southern Africa. Quat Sci Rev 22:2,311–2,326
Holmgren K, Risberg J, Freudendahl J, Ekblom A, Mugabe J, Sitoe S, Achimo M (2012) Variations in precipitation during the last 2300 years at Lake Nhauhache, Mozambique. J Paleolimnol 48:311–322
Huffman TN (2009) Mapungubwe and Great Zimbabwe: the origin and spread of social complexity in southern Africa. J Anthropol Rev 28:37–54
Hulme M (1995) Climate change within the period of meteorological records. In: Goudie AS (ed) The physical geography of Africa. Oxford University press, Oxford, pp 88–102
Hyde MA, Wursten BT, Ballings P, Dondeyne S (2002) Flora of Mozambique: Home page. http://www.mozambiqueflora.com/index.php. Accessed 16 Jan 2012
InsideWood (2004 onwards) http://insidewood.lib.ncsu.edu/search, Accessed 1 March 2012
Jacobsen GL, Bradshaw RHW (1981) The selection of sites for paleovegetational studies. Quat Res 16:80–96
Jarzen DM, Elsik WC (1986) Fungal palynomorphs recovered from recent river deposits, Luangwa Valley, Zambia. Palynology 10:35–60
Jonsson J (1998) Early plant economy in Zimbabwe. Dissertation, Uppsala University
Lebacq L (1955–1957) Atlas anatomique des bois du Congo. L’institut National Pour L’etude Agronomique Du Congo Belge, Brussels
Lee-Thorp JA, Holmgren K, Lauritzen S, Linge H, Moberg A, Partridge TC, Stevenson C, Tyson PD (2001) Rapid climate shifts in the southern African interior throughout the mid to late Holocene. Geophys Res Lett 28:4,507–4,510
Lejju BJ, Taylor D, Robertshaw P (2005) Late-Holocene environmental variability at Munsa archaeological site, Uganda. A multicore, multiproxy approach. Holocene 15:1,044–1,061
Lemmens RHMJ (2011) Androstachys johnsonii Prain. Internet Record from PROTA4U. In: Lemmens RHMJ, Louppe D, Oteng-Amoako AA (eds) PROTA (Plant Resources of Tropical Africa/Ressources végétales de l’Afrique tropicale), Wageningen, Netherlands. http://www.prota4u.org/search.asp. Accessed 2 March 2012
Liesegang G (1990) Achegas para o estudo das biografias de autores de fontes narratives e outros documentos da história de Moçambique, II, III: Três autores sobre Inhambane: Vida e obra de Joaquim de Santa Rita Montanha (1806–1870), Aron S. Mukhombo (ca. 1885–1940) e Elias S. Mucambe (1906–1969). Arquivo (Boletim do Arquivo Histórico de Moçambique) 8:61–142
Manyanga M (2006) Resilient landscapes: socioenvironmental dynamics in the Shashi-Limpopo basin, southern Zimbabwe c. AD 800. Dissertation, Uppsala university
Marinova E, Atanassova J (2006) Anthropogenic impact on vegetation and environment during the Bronze Age in the area of Lake Durankulak, NE Bulgaria: pollen, microscopic charcoal, non-pollen palynomorphs and plant macrofossils. Rev Palaeobot Palynol 141:165–178
Marston JM (2009) Modelling wood acquisition strategies from archaeological charcoal remains. J Archaeol Sci 36:2,192–2,200
Moore PD, Webb JA, Collins ME (1991) Pollen analysis. Blackwell, Oxford
Morais J (1988) The early farming communities of Southern Mozambique. Central Board of National Antiquities, Stockholm
Neumann K (1999) Charcoal from West African savanna sites: Questions of identification and interpretation. In: Van der Veen M (ed) The exploitation of plant resources in ancient Africa. Kluwer, Dordrecht, pp 205–219
Neumann K, Kahlheber S, Uebel D (1998) Remains of woody plants from Saouga, a medieval west African village. Veget Hist Archaeobot 7:57–77
Neumann K, Schoch W, Détienne P, Schweingruber FH (2001) Hölzer der Sahara und des Sahel. Woods of the Sahara and the Sahel. Bois du Sahara et du Sahel. Paul Haupt, Bern
Neumann K, Fahmy A, Lespez L, Ballouche A, Huysecom E (2009) The Early Holocene palaeoenvironment of Ounjougou (Mali): Phytoliths in a multiproxy context. Palaeogeogr Palaeoclim Palaeoecol 276:87–106
Normand D (1950–1960) Atlas des bois de la Cote d’Ivoire. Centre Technique Forestier Tropical, Nogent-sur-Marne
Pitkänen A, Lehtonen H, Huttunen P (1999) Comparison of sedimentary microscopic charcoal particle records in a small lake with dendrochronological data: evidence for the local origin of microscopic charcoal produced by forest fires of low intensity in eastern Finland. Holocene 9:559–567
Prager A, Barthelmes A, Theuerkauf M, Joosten H (2006) Non-pollen palynomorphs from modern alder carrs and their potential for interpreting microfossil data from peat. Rev Palaeobot Palynol 141:7–31
Prior JAB, Gasson PE (1990) Comparative wood anatomy of afromontane and bushveld species from Swaziland, southern Africa. IAWA Bull n.s. 2:319–336
Punt W, Blackmore S, Hoen PP, Stafford PJ (2003) The Northwest European pollen flora. Elsevier, Amsterdam
Raper D, Bush M (2009) A test of Sporormiella representation as a predictor of megaherbivore presence and abundance. Quat Res 71:490–496
Rita-Ferreira A (1999) African kingdoms and alien settlements in central Mozambique (c.15th–17th cent.). Centro de Estudos Africanos, Departamento de Antropologia, Universidade de Coimbra
Schmidt PR (1994) Historical ecology and landscape transformation in Eastern equatorial Africa. In: Crumley CL (ed) Historical ecology, cultural knowledge and changing landscapes. School of American Research Press, Santa Fe, pp 99–126
Schmidt PR (1997) Archaeological views on a history of landscape change. J Afr Hist 38:395–421
Shackleton CM, Prins F (1992) Charcoal analysis and the principle of least effort—a conceptual model. J Archaeol Sci 19:631–637
Sinclair PJJ (1982) Chibuene an early trading site in southern Mozambique. Paideuma 28:49–64
Sinclair PJJ (1987) Space, time and social formation: A territorial approach to the archaeology and anthropology of Zimbabwe and Mozambique c. 0–1700 AD. Dissertation, Uppsala University
Sinclair PJJ (1995) The origins of urbanism in Eastern and southern Africa: A diachronic perspective. In: Sahlström B, Ådal K (eds) Islamic art and culture in Africa. Acta Universitatis Upsaliensis, Uppsala, pp 99–110
Sinclair PJJ (1997) Human responses and contributions to environmental change in Africa. In: Sitter-Liver B, Uehlinger C (eds) Partnership in archaeology: perspectives in a crosscultural dialogue. Fribourg University Press, Fribourg, pp 179–197
Sinclair PJJ, Ekblom A (2004) Chibuene an early first millenenium trading site in southern Mozambique. A field report of the 1995–1999 excavations. Uppsala (Unpublished MS)
Sinclair PJJ, Ekblom A, Wood M (2012) Chibuene, understanding the dynamics between evolving social complexity, long distance trade and state formation in southern Africa in the late first millennium AD. Antiquity 86:723–737
Stockmarr J (1971) Tablets with spores used in absolute pollen analyses. Pollen Spores 13:615–621
Telford S, De Castro A (2001) Specialist study 4, flora and fauna. Sasol. http://www.sasol.com/natural_gas/Environment/GASFIELD%20Document%20PDF/GASFIELD%20Specialist%20Reports/Report%204%20-%20Flora%20&%20Fauna/Gas%20Field%204%20-%20Flora%20and%20Fauna.pdf. Accessed 1 Aug 2001
Theal G (1964) Records of southeastern Africa. C. Struik Ltd, Cape Town
Thompson G, Young R (1999) Fuels for the furnace. Recent and prehistoric ironworking in Uganda and beyond. In: Van der Veen M (ed) The exploitation of plant resources in ancient Africa. Kluwer, Dordrecht, pp 221–239
Tomlinson RW (1973) The Inyanga area, an essay in regional biogeography. University of Rhodesia, Salisbury
Tusenius M (1989) Charcoal analytical studies in the northeastern Cape, South Africa. S Afr Archaeol Bull Goodwin Ser 6:77–83
Tyson PD, Preston-Whyte RA (2000) The weather and climate of Southern Africa. Oxford University Press, Oxford
Van Geel B, Aptroot A (2006) Fossil ascomycetes in Quaternary deposits. Nova Hedwigia 82:313–329
Van Geel B, Hallewas DP, Pals JP (1983) A Late Holocene deposit under the Westfriese Zeedijk near Enkhuizen (Prov. of N-Holland, The Netherlands): palaeoecological and archaeological aspects. Rev Palaeobot Palynol 38:269–335
Van Geel B, Klink AG, Pals JP, Wiegers J (1986) An Upper Eemian lake deposit from Twente, eastern Netherlands. Rev Palaeobot Palynol 47:31–61
Van Geel B, Buurman J, Brinkkemper O, Schelvis J, Aptroot A, Van Reenen G, Hakbijl T (2003) Environmental reconstruction of a Roman Period settlement site in Uitgeest (The Netherlands), with special reference to coprophilous fungi. J Archaeol Sci 30:873–883
Vogelsang R, Eichhorn B (2011) Under the mopane tree. Africa Praehistorica 24. Heinrich-Barth-Institut, Köln
Wheeler EA, Baas P, Gasson PE (1989) IAWA list of microscopic features for hardwood identification by an IAWA committee. IAWA Bull New Ser 10:219–332
Wild H, Fernandes A (1968) Vegetation map of the flora Zambesiaca area, supplement. Flora Zambesiaca. The Government Printer, Salisbury
Wood M (2000) Making connections: Relationships between international trade and glass beads from the Shashe-Limpopo area. In: Lesley M, Maggs T (eds) African Naissance: the Limpopo valley 1000 years ago. South African Archaeological Society, Vlaeberg, pp 78–90
Wood M (2012) Interconnections: glass beads and trade in southern and eastern Africa and the Indian Ocean—7th to 16th centuries AD. Department of Archaeology and Ancient History, Uppsala
Acknowledgments
The research was sponsored by SAREC (Swedish agency for research cooperation). Fieldwork was carried out as collaboration between Dept. of African and Comparative Archaeology, Uppsala University in cooperation and Dep de Anthropologia e Archeologia, Eduardo Mondlane University, Mozambique under the auspices of HRAC (Human responses and contributions to environmental change). Prof Louis Scott, Dept. of Plant Sciences, University of the Free State, Bloemfontein, contributed greatly in the pollen identification process. We would also like to thank Katharina Neumann for advice on wood charcoal analyses and Manfred Ruppel for SEM technical assistance. Elisabeth Green kindly revised the language.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by F. Bittmann.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Ekblom, A., Eichhorn, B., Sinclair, P. et al. Land use history and resource utilisation from a.d. 400 to the present, at Chibuene, southern Mozambique. Veget Hist Archaeobot 23, 15–32 (2014). https://doi.org/10.1007/s00334-013-0392-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-013-0392-4