
Abstract
In some continental and island sites in the western Mediterranean basin, the Holocene vegetation and climate dynamics seem to show the same patterns in time and space. Nevertheless, different synchronous scenarios have been proposed from other south European, North African and Near Eastern pollen records from around the Mediterranean basin. Striking similarities and synchronisms have been found between Sicily and the Balearic Islands. These islands, although under different bioclimatic influences, show similarities in the main trends of vegetation and climate dynamics during the Post-glacial. Lago di Pergusa is the only natural inland lake in Sicily and because of its geographical location, has a good potential sensitivity to the climatic changes of the Mediterranean basin. Likewise, coastal sediments from Minorca and Majorca, the Balearic Islands, have similar peculiarities. The present-day environmental situation, now that most of the natural vegetation in these islands has disappeared, has been brought about either by a climatic trend towards increasing aridity or an increase in human activities. It seems clear that prehistoric human people alone could not have caused all the environmental changes recorded in the last millennia in both places.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction and environmental setting
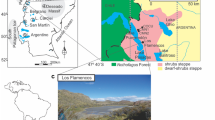
During the 1990s, the palynological laboratories of Universitat Autònoma de Barcelona and of Dipartimento di Biologia Vegetale, Università di Roma “La Sapienza” were involved in the study of desertification processes in southern Mediterranean Europe as part of two projects financed by the European Union. The interaction of climatic and anthropogenic factors has determined the present-day structure and evolution of the vegetation. The landscape history of each island can be read in the sediment records. In particular, the Balearic Islands and Sicily (Fig. 1) appear to have been very sensitive to climatic change in determining their geographical and biogeographical peculiarities.

Location map of the sites used for comparison: (1) Alcúdia (Majorca), (2) Algendar (Minorca) and (3) Lago di Pergusa (Sicily)
Its geographical position, geological, climatic and botanical features, prehistoric settlements and historical events make Sicily a privileged observation point for the climatic phenomena in the Mediterranean basin during the Holocene. Climate and geomorphology play a leading role in determining the present-day distribution of vegetation types. Sicily shows a considerable diversity from a floristic point of view, with its 3000 species of which 10% are endemic (Brullo et al. 1995). It is worth noticing that in few kilometres the vegetation can change from a maquis scrub with dwarf palm along the coastal area to beech woods in the mountains. The high biodiversity of Sicily was recently confirmed by the finding of small trees of Zelkova sicula (Di Pasquale et al. 1992) in the Iblei mountains.
The location of Lago di Pergusa (667 m a.s.l., 37°31′ N; 14°18′E) in the centre of Sicily, in the heart of the Mediterranean region, but a few km away from Enna, the highest provincial chief town in Italy (931 m a.s.l.) (Fig. 1), together with the human history of its district, make this site important for an understanding of environmental variability phenomena. The surroundings of the lake, with its central position in the island (at least 50 km from the sea) and with the elevation of the catchment between about 670 m and 910 m a.s.l., reduces any climatic moderation from the sea. This main extant inland natural lake of Sicily is also very sensitive to climatic change because of the absence of both inflows and outflows, being fed only by atmospheric precipitation and alkaline water-bearing strata. On the other hand, mean annual precipitation in the area is about 600 mm, reaching about 900 mm at the high weather station of Enna, and snow in winter is not so rare. In the absence of human impact, the vegetation of the zone should consist of thermophilous woods with deciduous and evergreen elements. The natural woodland consists of sparse trees of Quercus suber L. (cork oak), Q. virgiliana (Ten.) Ten. and Q. ilex L. (holm oak). This land is now widely cultivated for cereal crops.
The surroundings of the lake (Bernabò Brea 1961; Giannitrapani and Pluciennik 1998; Tusa 1992) have been inhabited since the Neolithic period; many important sites located in strategic hilly positions close to the lake date back to the Bronze Age; in pre-Roman times Enna was held by Greeks, Syracusans and Carthaginians. As a municipium at the time of the Roman Empire, it was not of great importance being mainly a hub centre, and then its history was the same as that of the rest of its region, with Greek and then Byzantine domination, followed by invasions of Arabs and Normans.
The Balearic Islands lie 80–300 km east of the Spanish mainland and have a land area of 5014 km2. There are two groups of islands. The eastern Balearic Islands include Majorca and Minorca (Fig. 1) and the small island of Cabrera. Minorca, with an area of 701 km2, has rugged hills rising in the north to the hill of Toro (358 m). The south coast, where the studied sequences are located, is more regular and has long stretches of cliffs. The western Balearic Islands include Eivissa (Ibiza) and Formentera.
The climate of the island of Minorca can be described as typically Mediterranean, with a dry period in summer, two equinoctial rainfall maxima and mild winter temperatures. Annual precipitation is low, rarely exceeding 450 mm, and it occurs mainly in autumn and spring. The mean annual temperature is about 16.8 °C.
Bolòs (1981) remarks that the temperature decrease during glacial periods in the Balearic Islands was not as marked as in continental environments, since it permitted the survival of thermophilous taxa from the Tertiary flora. In Minorca, the potential vegetation is typically Mediterranean, made up of Balearic evergreen oak woods with Quercus ilex as the main component and, in lower and windier areas, shrub vegetation formations related to thermomediterranean woods (Peinado and Rivas-Martínez 1987). These are dominated by Olea europaea var. sylvestris (wild olive), Prasium majus, Euphorbia dendroides and Phillyrea rodriguezii (Rivas Martínez 1987).
The Balearic Islands have been, from prehistoric times to more recent times, and because of their strategic location in the heart of the western Mediterranean, a staging point of different cultures (Micó 2005). Since the dawn of civilization their location has attracted different peoples who have coveted the islands as a stopover port and a shelter, and who have left a rich historical legacy there. Human presence in the Balearic Islands has been verified from the beginning of the Bronze Age (2000 b.c.), called the pre-Talayotic period, from which there are important burial monuments such as megalithic tombs and collective tombs. Around 1400 b.c., the evolutionary process of this culture produced various large stone tower constructions known as talayots. This word gives rise to the name of the richest prehistoric period of the island, namely, the Talayotic period, at which time there was a great increase in the number of settlements. The indigenous culture soon came under significant foreign influence from expanding merchant peoples such as the Carthaginians, who above all were known for introducing new tools and ornaments. In 123 b.c. the Roman conquest occurred and this brought with it the transformation of the Talayotic settlements and the development of important ports. Pottery, mostly of the native Talayotic types, seems to have persisted with little change until the Roman occupation. Historical evidence points to at least 2600 years of settlement on the islands, which were successively ruled by Carthaginians, Romans, Vandals, Moors, Catalans and Spaniards, all of whom have left their mark. The Balearics were conquered by the Vandals in a.d. 526 and fell to the Byzantines in a.d. 534. The Muslim occupation of the islands was complete in a.d. 903. Jaume I d’Aragó (Aragon) conquered the islands of Majorca and Ibiza between 1229 and 1235, and Minorca fell to his descendant, Alfonso III, in 1287.
Material and methods
The material used in this paper consists of the sediment records recently available from Sicily and the Balearic Islands (Pérez-Obiol et al. 2000; Sadori and Narcisi 2001). For coring operations, sediment descriptions, methodology and detailed pollen diagrams we refer the reader to the available papers quoted below. The Holocene lake sediments of Lago di Pergusa were investigated for magnetic susceptibility, lithostratigraphy, tephra, pollen, AMS radiocarbon dating (Sadori 2001; Sadori and Narcisi 2001; Narcisi 2002) and micro-charcoal analyses (Sadori and Giardini 2004a, b). The sediment records from the Balearic Islands were obtained from littoral deposits in lagoons and marshlands on the coastal plains and were investigated for pollen and chronology, with AMS and conventional radiocarbon dates (Burjachs et al. 1994; Yll et al. 1997; Pérez-Obiol et al. 2000). Even if the environments from where the pollen records were taken seem quite different, coastal lagoons and marshes on the Balearic Islands, and an inland mountain lake in Sicily, more factors have to be considered. All the presented sites have water bodies fed mainly by precipitation and ground waters; the strict correlation with the water table makes them very sensitive to climate change. The present-day lacustrine brackish vegetation of Lago di Pergusa (the lake water has a chloride-sulphate alkaline composition) is typical of a marine coastal environment and not observed in any other inland lake in Sicily.
Another point, which must be considered, is the chronology of the records. In terrestrial environments, except for long sedimentary sequences with annual laminations that are radiocarbon and/or tephrochronologically controlled, no records can have a strong chronology. The common practice in palynology of using pollen stratigraphies for comparing not only between terrestrial records, but also terrestrial with marine ones, should be carefully avoided in absence of independently dated sediment records.
There is no doubt that the radiocarbon chronology of the pollen records presented here could have been better assessed if, for example, more terrestrial plant macro remains had been available. Nevertheless some efforts were made in this study to put together some tesserae to better reconstruct the vegetation history of the Mediterranean basin. Due to the doubts on chronological assessments of pollen records just explained and to avoid further approximations, extremely precise timings and calibration of the millennial sediment scales were not tried. Superimposed pollen curves are used to ease the direct comparison between the vegetation histories of Sicily and the Balearic Islands. AP (arboreal pollen) percentage and concentration curves (as well as for single taxa) are overlapped for comparison among different sites (Figs. 2–6). The aim is to observe similarities and/or dissimilarities both in long trends and in rapid changes of the vegetation cover and to evaluate the possibility to point out synchronisms and/or diachronisms in its development on the Balearic Islands and on Sicily.

Lago di Pergusa (Sicily) and Algendar (Minorca). Arboreal pollen (AP) percentage diagrams

Lago di Pergusa (Sicily) and Algendar (Minorca). Total pollen concentration diagrams

Lago di Pergusa (Sicily) and Alcúdia (Majorca). Fagus pollen percentage diagrams

Lago di Pergusa (Sicily), Alcúdia (Majorca) and Algendar (Minorca). Corylus pollen percentage diagrams

Lago di Pergusa (Sicily) and Algendar (Minorca). Olea pollen percentage diagrams
Results
Comparison between the studied sites
In the Balearic Islands, sediment accumulation is strongly controlled by changes in eustatic sea level. In the study area, Quaternary sedimentation began in coastal areas after the last important regression-transgression cycle, at about 8000 b.p.
All the pollen diagrams from the Balearic Islands follow a similar pattern. An initial assessment shows a change in taxa during the middle Holocene that implies changes in the landscape. The replacement took place rapidly, from around 5000 to around 4000 b.p. Some taxa as Buxus and Corylus were replaced by others such as Olea, a phenomenon which reflects, in our point of view, a global climatic trend. At the same time the disappearance of endemic animal taxa such as Myotragus and Hypnomys took place. The landscape changes were probably caused mainly by an increase in dryness. Several mesophilous taxa (for example Corylus, Alnus, deciduous Quercus and Buxus) do not grow on the island any more. At present, Corylus grows in a very restricted area on the northern mountains of Majorca. There, the mist in the mountains, favoured by the topography, helped some communities of Fagus to grow around the mid-Holocene (Fig. 4).
In Sicily at the end of the last glacial period, the land was covered by an open vegetation with dominant Artemisia, Gramineae and Chenopodiaceae. Sparse pollen grains of Pinus (pine) (probably overrepresented) and angiosperm trees are the only traces of trees.
During the Post-glacial period, wet conditions occurred for about two millennia from 9000 b.p. when the maximum expansion of woodland was recorded, with a peak at the estimated age of 8100 b.p. The same phenomenon took place in the Balearic Islands where, at ca. 8000 b.p., the climate favoured the formation of mesophilous vegetation in a landscape that has no present-day analogy. No evidence of the aridity trend beginning around 7200 b.p. which was recorded at Lago di Pergusa, is found at Minorca. An aridity period was however pointed out in continental Spain by Jalut et al. (2000) from 7500 to 7000 b.p. A slow and prolonged climate forcing probably accounts for its time-transgressive character.
On both the Balearic islands and on Siciliy, the middle Holocene marks important changes when the AP concentrations declined in a few centuries and never significantly recovered (Fig. 3). In Sicily, until about 2800 b.p., the structure of the vegetation showed some changes, with deciduous oaks decreasing and evergreen elements becoming relatively more important, the main vegetation change being found in the reduced evidence for woodland and in slowly increasing numbers of Mediterranean taxa. Then, between about 2800 and 2400 b.p. evergreen elements became dominant and Olea reached its maximum expansion, which is interpreted as the clear effect of human impact on the landscape. In the last two millennia, evergreen and deciduous vegetation types show similar low frequencies. In the Balearic Islands, the process was similar with differences in the vegetation composition. The significant landscape change is reflected in composition and structure, resulting in a vegetation adapted earlier to Mediterranean conditions.
The different importance of the wooded periods in Sicily and in the Balearic Islands is reflected in the AP values (Fig. 2). In the period between 9000 and 4000 b.p. at Lago di Pergusa the trees show percentages mainly at 80%. In contrast, the AP values are generally below 50% in the Balearic Islands. Apart from the dissimilarity in the vegetation, consisting of a marked difference in the woodland canopy and explicable by the present-day phytogeographical differences between the sites, it is interesting that the two curves show parallel trends. Nevertheless, the fall of the AP percentages recorded in both areas somewhat before 4000 b.p. is not synchronous, with the most important decrease in the pollen concentration (Fig. 3), which appears to be still earlier. It could be affirmed that while the biomass, the extension of the vegetation cover, decreases during the first part of the Holocene, there is no significant change in the AP/NAP ratio. On the other hand, the diachronisms present in both Figs. 2 and 3 between the two sites may be explained (in absence of evidence of wrong datings) as a time lag between the plant biomass decrease (detected by pollen concentration) and percentage pollen representation.
If we assume that a strong decrease in pollen concentration could be mainly related to a decrease in trees, it appears clear that the vegetation structure would be the first to change in response to a climate change. Only subsequently, a change in the vegetation, consisting in this case of an opening of the woodland in favour of herbs, could be found. In Fig. 3 we can appreciate two clear stages with different pollen concentration values and a third one. The first phase shows high pollen concentration values indicating important biomass values until ca. 7000 b.p., while the second one consists of a strong decrease coinciding with the expansion of Mediterranean and more xeric elements in both sites. A third stage, recorded only in Sicily, since the most recent sediments from this part in the Balearic Islands are absent, indicates that the phytomass decreased even more in historical times.
Different behaviours are found in some specific taxa in both studied sites (Fig. 4). In Majorca, the slight increase of Fagus at around 5000 b.p. can be interpreted as the response to an increase in sea surface temperature that promoted the formation of mist in the mountains. It is difficult to explain the Fagus curve from Majorca according to its Holocene spread in the rest of Europe (Petit et al. 2003). Nowadays Fagus is not present in Majorca. In the absence of further phylogeographical data on beech distribution, taking into account the geographical distribution of a taxon in establishing its phylogeny, and understanding the geographical patterns that may result from divergence, ultimately leading to speciation, it could be suggested that Fagus stands were present in the island then. In Sicily, Fagus was continuously present even if with low values. This seems to indicate the persistence of scattered beech trees in the vicinity of the lake or of Fagus woods in mountain areas of the island throughout the Holocene. At present, beech woods are found all around Pergusa, at a distance of some tens of kilometres, and on the mountains Etna, Madonie, Nebrodi and Iblei.
The behaviour of Corylus (Fig. 5) also deserves comparison. It is well known that in several regions of Europe Corylus pollen peaked during the Boreal period and then decreased. This pattern seems to be followed in Sicily but not in the Balearic Islands, where high values of Corylus persisted until the middle Holocene. On the other hand, Olea curves follow similar tendencies in both sites, increasing from the middle Holocene (Fig. 6). It is clear that this phenomenon is related to the establishment of a Mediterranean climate with increased seasonality and a significant dry period. It is however quite impossible to decide if the early Neolithic increase of Olea and its Bronze Age significant expansion found at Lago di Pergusa could be due to climate change or early cultivation. The best explanation can probably be found in a combination of the two causes. It should be considered that the vegetation in large areas of both islands is a maquis of Olea europaea var. sylvestris (wild olive). Another important question is to discover whether the beginning of intensive Olea cultivation can be detected. A pollen diagram from the east of the Iberian Peninsula shows an increase of Olea pollen during medieval times. The same phenomenon occurs in the southeast of the Iberian Peninsula where the steppe-like vegetation was modified during the Middle Ages by cultivated Olea and other anthropogenic indicators (Follieri et al. 2000), which is consistent with the available historical data from both quoted localities. At Lago di Mezzano (Sadori, in prep.) the Olea curve seems to have largely the same trend as in Spain, with an increase in medieval times. As already mentioned, something different is found in Sicily at Lago di Pergusa, where an important expansion is found at the end of the Bronze Age and during the Iron Age, when anthropogenic indicators such as legumes, cereals, Castanea (chestnut) and Juglans (walnut) are found. In the same period, a similar expansion of Olea is recorded 40 km north of Rome, at Lago di Vico (Magri and Sadori 1999) where an undiscussed human impact is found, clearly indicated by intensive cultivation of trees, first Castanea and then Juglans.
Conclusions
The pollen data from Sicily show that the wettest conditions were attained in the period between 9000 and 7200 b.p. Then a trend towards aridification began, leading to very dry conditions at about 3000 b.p. An unquestionable human impact on vegetation is found since 2800 years b.p. As climate forcing and increasing human land use had already induced vegetation change, the human occupation of the last three millennia did not produce further significant effects on the environment. The pollen data from the Balearic Islands reflects important changes in the vegetation during the middle Holocene (5000–4000 b.p.) showing the substitution of mesophilous communities by a maquis scrub. The causes of these events remain unclear since they seem to have been increased, but not caused, by human action. Recently, it has been pointed out that many south-western European pollen records show rather substantial differences between the early and late Holocene, suggesting a general change from wetter to drier climatic conditions with clear aridification phases (Jalut et al. 2000; Magny et al. 2002). This trend was indicated since around 7500 b.p. (as detected in Sicily) by slight increases of sclerophyllous taxa and culminated around 4500–4000 b.p. (as strongly indicated in Minorca). The most reasonable hypothesis is that the changes were accelerated by human activity.
These similarities between the two areas under consideration are found in parallel vegetational trends, reflected both in AP percentages and in total pollen concentration data, suggesting the fundamental role of the climate in shaping the different environments and landscapes in the Mediterranean basin, in sites chosen for their high sensitivity to climate change. Dissimilarities are found in single taxon percentage values and trends, indicating that the vegetation dynamics and even the floristic composition were deeply different in the two considered areas. Fagus pollen can be given as an example of this diversity, continuously present in the diagram from Sicily and present around the middle of the Holocene in Majorca.
As a consequence of the similarities and dissimilarities in the pollen diagrams, some synchronisms and diachronisms are shown. The main and similar Holocene vegetation changes, which can be considered to have been determined by climatic events, seem to happen in the same chronological intervals. Expansions of single taxa such as Corylus were not only different from a quantitative point of view, but also from a chronological perspective, showing a remarkable diachronism in their histories in the two studied areas.
To conclude, the two main vegetation changes found in the Balearic Islands and Sicily, recorded before 7000 b.p. and at about 4500 b.p. were (1) a change of structure (detected by strong decreases in pollen concentration) and then (2) a change of landscape (decreases in tree pollen percentages). The first change may be a consequence of the well known north Atlantic 8200 cal b.p. short and abrupt event. Its effect seems to have been detected in many European sediment records (Magri and Sadori 1999; Mercuri et al. 2004; Ramrath et al. 2000; Seppä and Veski 2004; Tinner and Lotter 2001). This sharp and short climate signal induced a dry period in the southwestern Mediterranean area (Jalut et al. 2000) and could have triggered a change in the woodland canopy and vegetation composition of the investigated islands. The second event is also clearly visible in many southern European lacustrine sites (Catalan et al. 2001; Follieri et al. 1988; Jalut et al. 2000; Magri and Sadori 1999; Sadori 2003; Schneider 1985) where significant changes in the vegetation are found around 4500 b.p.
References
Bernabò Brea L (1961) La Sicilia prima dei Greci, 3rd edn. Il Saggiatore, Milano
de Bolòs O (1981) El Clima i la Vegetació post-glacial als Països Catalans [The Post-glacial climate and vegetation of Catalunya, in Catalan]. Treballs Institució Catalana Història Natural 9:83–91
Brullo S, Minissale P, Spampinato G (1995) Considerazioni fitogeografiche sulla flora della Sicilia. Ecologia Mediterranea 21:99–107
Burjachs F, Pérez-Obiol R, Roure JM, Julià R (1994) Dinámica de la vegetación durante el Holoceno en la isla de Mallorca. In: Mateu I, Dupré M, Güemes J, Burgaz ME (eds) Trabajos de palinología básica y aplicada. X Simposio de Palinología:199–210. Universitat de València, València
Catalan J, Camarero L, Pla S, Buchaca T, Rieradevall M, Bordonau J, Josep Peñuelas JG, Pérez R (2001) A climatic interpretation of 15,000 years of environmental variability in the central Pyrenees. Abstract PAGES – PEPIII: Past Climate Variability Through Europe and Africa. August 27–31, 2001. Aix-en-Provence, France
Di Pasquale G, Garfi G, Quézel P (1992) Sur la presence d’un Zelkova nouveau en Sicile sud-orientale (Ulmaceae). Biocosme Mésogéen 8/9:401–409
Follieri M, Roure JM, Giardini M, Magri D, Narcisi B, Pantaleón-Cano J, Pérez-Obiol R, Sadori L, Yll EI (2000) Desertification trends in Spain and Italy based on pollen analysis. In: Balabanis P, Peter D, Ghazi A, Tsogas M (eds) Mediterranean desertification. Research results and policy implications. Proceedings of the International Conference, 29 October to 1 November 1996, Crete, Greece, vol. 2, pp. 33–44
Giannitrapani E, Pluciennik M (1998) La seconda campagna di ricognizione (settembre 1997) del progetto Archeologia nella valle del Torcicoda. Sicilia Archeologica 96:59–69
Jalut G, Esteban Amat A, Bonnet L, Gauquelin T, Fontugne M (2000) Holocene climatic changes in the Western Mediterranean, from south-east France to south-east Spain. Palaeogeogr Palaeoclim Palaeoecol 160:255–290
Magny M, Miramont C, Sivan O (2002) Assessment of the impact of climate and anthropogenic factors on Holocene Mediterranean vegetation in Europe on the basis of palaeohydrological records. Palaeogeogr Palaeoclim Palaeoecol 186:47–59
Magri D, Sadori L (1999) Late Pleistocene and Holocene pollen stratigraphy at Lago di Vico (central Italy). Veget Hist Archaeobot 8:247–260
Mercuri AM, Accorsi CA, Bandini Mazzanti M (2004) Genesis and evolution of the cultural landscape in Italy as suggested by central Adriatic pollen diagrams (PALICLAS project). Polen 14:229–230
Micó R (2005) Cronología absoluta y periodización de la prehistoria de las Islas Baleares. British Archaeological Reports International Series 1373, Archaeopress, Oxford
Narcisi B (2002) Tephrostratigraphy of the late Quaternary lacustrine sediments of Lago di Pergusa (central Sicily). Boll Soc Geol It 121:211–219
Peinado M, Rivas-Martínez S (1987) La vegetación de España. Servicio Publicaciones Universidad de Alcalá de Henares, Alcalá de Henares
Pérez-Obiol R, Yll EI, Pantaleón-Cano J, Roure JM (2000) Evaluación de los impactos antrópicos y los cambios climáticos en el paisaje vegetal de las Islas Baleares durante los últimos 8000 años. In: Guerrero V, Gornés S (eds) Prehistoria de Baleares, pp. 73–89
Petit RJ, Aguinagalde I, de Beaulieu JL, Bittkau C, Brewer S, Cheddadi R, Ennos R, Fineschi S, Grivet D, Lascoux M, Mohanty A, Muller-Starck GM, Demesure-Musch B, Palme A, Martin JP, Rendell S, Vendramin GG (2003) Glacial refugia: hotspots but not melting pots of genetic diversity. Science 300:1563–1565
Ramrath A, Sadori L, Negendank JFW (2000) Sediments from Lago di Mezzano, central Italy: a record of Late Glacial/Holocene climatic variations and anthropogenic impact. The Holocene 10:87–95
Rivas Martínez S (1987) Memoria del mapa de series de vegetación de España. 1:400.000, ICONA, Madrid
Sadori L (2001) Holocene climatic change in central Sicily (Italy). Terra Nostra 3:181–186
Sadori L (2003) Late-Glacial and Holocene fire and vegetation history from Lago di Mezzano, central Italy. 16th INQUA Congress, Reno, Nevada, 23/30 luglio, p. 193
Sadori L, Giardini M (2004a) A new combined approach to detect Holocene environmental changes: the different roles of man and climate. 32nd International Geological Congress, 20–28 August 2004, Florence, Italy
Sadori L, Giardini M (2004b) The role of fire in the Mediterranean landscapes: an intriguing question. 3rd International Meeting of Anthracology. Charcoals from the past, Lecce June 28th–July 1st, pp. 40–41
Sadori L, Narcisi B (2001) The postglacial record of environmental history from Lago di Pergusa (Sicily). The Holocene 11:655–671
Schneider R (1985) Analyse palynologique dans l’Aspromonte en Calabre (Italie meridionale). Cahiers ligures de préhistoire et de protohistoire, Nouvelle série 2:279–288
Seppä H, Veski S (2004) The response of the vegetation to the cold event 8200 years ago in Estonia, Eastern Europe. Polen 14:210–211
Tinner W, Lotter AF (2001) Central European vegetation response to abrupt climate change at 8.2 ka. Geology 29:551–554
Tusa S (1992) La Sicilia nella preistoria, 2nd edn., Sellerio, Palermo
Yll EI, Perez-Obiol R, Pantaleon-Cano J, Roure JM (1997) Palynological evidence for climatic change and human activity during the Holocene on Minorca (Balearic Islands). Quat Res 48:339–347
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Pérez-Obiol, R., Sadori, L. Similarities and dissimilarities, synchronisms and diachronisms in the Holocene vegetation history of the Balearic Islands and Sicily. Veget Hist Archaeobot 16, 259–265 (2007). https://doi.org/10.1007/s00334-006-0038-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-006-0038-x