
Abstract
As the Arctic becomes increasingly accessible to the resource extraction industry, an understanding of the impacts of infrastructure and anthropogenic disturbance on tundra-nesting avian populations is critical. We conducted breeding bird surveys using point counts and Program for Regional and International Shorebird Monitoring (PRISM) rapid surveys at the Hope Bay Project, an underground gold mine in Nunavut, Canada, from 2007 to 2015. We examined the relationship between abundance of total birds, songbirds, and shorebirds with distance from infrastructure, anthropogenic disturbance (i.e., activity at the mine site), and environmental covariates. Contrary to our predictions, anthropogenic disturbance showed no significant effect on bird abundance for either survey type or bird grouping. However, total bird abundance was associated with distance from infrastructure for both survey methods. A relationship with distance to infrastructure, with variability in influence, was also present for songbird point count data and shorebird PRISM data, but was not significant for songbirds recorded during PRISM surveys. Habitat type played a vital role in determining avian abundance, with PRISM surveys showing that wet lowland ecotypes supported higher bird numbers, whereas point count data revealed a more nuanced habitat association, likely reflecting the specific habitat preferences of different songbirds. Our study highlights the need for strategic planning of industrial development, incorporating comprehensive monitoring and ensuring the protection of preferred Arctic-breeding bird habitats. By aligning development projects with environmental objectives, we can ensure the coexistence of industrial interests and the future of avian populations in the Arctic.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Avian populations are frequently used as indicators of biodiversity and environmental health (Gregory and Strien 2010). The consistent long-term monitoring of avian species can play a crucial role in understanding population trends of Arctic birds and the impacts of anthropogenic development and climate change (Gregory and Strien 2010). Climate-driven loss of suitable Arctic-breeding habitat is predicted to impact 66–83% of tundra specialist species over the next 60 years (Wauchope et al. 2017). Additionally, as the Arctic becomes increasingly accessible due to the amplified impacts of climate change at high latitudes, resource exploration and development are growing concerns for species distribution (Staniforth 2002; Wauchope et al. 2017) and reproductive success (Kendall 2011). Across the circumpolar Arctic, breeding abundance of wading birds has declined by 50% while land birds have decreased by 20% (Smith et al. 2020). At local scales, populations of Arctic-breeding birds naturally fluctuate due to food availability, weather, timing of snow melt, species settlement strategies, and predation rates (Latour et al. 2005; Samelius et al. 2007; Mckinnon et al. 2013; Saalfeld and Lanctot 2015). Habitat quality also varies across the landscape; breeding birds are typically more abundant and diverse in lowland habitats, dominated by moist to wet sedge meadows (Johnston et al. 2000; Latour et al. 2005).
Industrial development is a primary driver of economic growth in the Arctic (Kondratiev 2020), and industrial projects such as mining operations are implementing increasingly more involved methods to monitor and mitigate effects to the ecosystem (Kullerud 2011; Tolvanen et al. 2019). Developers are required to conduct monitoring programs to determine whether the project is impacting wildlife or other ecosystem components. Industrial developments, such as the construction and operations of mines, can have several potential direct and indirect anthropogenic (human-derived) effects on bird populations. Anthropogenic effects include loss or degradation of habitat, disturbance (e.g., noise), and enhanced depredation (National Research Council 2003; Bayne et al. 2008; Francis et al. 2009). The spatial scale over which anthropogenic effects may reduce bird abundance, diversity, and nest survival are generally recorded at microscale extents of < 1 km (Bayne et al. 2008; Liebezeit et al. 2009; Benítez-López et al. 2010) but there is evidence of negative impacts out to 5 km (Liebezeit et al. 2009). Additionally, effects of industrial development may be challenging to differentiate from natural annual variation (Morrison et al. 2008).
Breeding bird populations can demonstrate temporal and spatial variation due to several environmental factors. Moisture levels and the associated vegetation community, along with tundra microrelief, are key indicators of nest-site selection and abundance (Latour et al. 2005; Cunningham et al. 2016). Snow cover and timing of snowmelt are also primary predictors of breeding phenology (Latour et al. 2005; Smith et al. 2010; Liebezeit et al. 2014; Cunningham et al. 2016; Saalfeld et al. 2017) and are correlated with prey availability, which is crucial for brood rearing (Meltofte et al. 2007; Cunningham et al. 2016; Saalfeld et al. 2019). Other weather variables may also influence spatiotemporal trends of birds; lower passerine abundance in the Arctic has been correlated with summers of higher precipitation and cooler temperatures (Robinson et al. 2014).
Our primary study objective was to measure breeding bird abundance from 2007 to 2015 in relation to an underground gold mining project in the Canadian Arctic, the Hope Bay Project. For the purpose of this study, breeding bird refers to songbirds, shorebirds, and ptarmigans. We tested whether any measurable effects on breeding birds could be detected in association with proximity to mine infrastructure and during years of higher anthropogenic disturbance defined as year with apparent activity at site (i.e., active construction and operations). Based on findings from existing anthropogenic effect studies (Bayne et al. 2008; Liebezeit et al. 2009; Benítez-López et al. 2010), we predicted a measurable impact of project infrastructure on breeding birds at distances of < 1 km from mine infrastructure, with a greater zone of influence in years with higher anthropogenic disturbance. We also tested whether natural spatial (habitat) and temporal (climate) variables accounted for disparity in bird abundance with the aim of disentangling mine effects from natural variation.
Methods
Study area
The Hope Bay Belt is an 80 km long Archaean age greenstone belt in Nunavut, Canada, which contains three gold deposits (Doris, Madrid, and Boston). Our study area, Hope Bay, includes an underground gold mine, the Hope Bay Project, which consists of infrastructure and exploration projects along all three gold deposits of the Hope Bay Belt. From 2007 to 2010, Newmont Mining Corporation constructed the Doris Camp and began underground mine development. The project footprint resulted in the direct loss of 83.0 ha of natural terrain, with the largest loss occurring in Betula–Ledum–Lichen mix (BL; 33.6 ha), Eriophorum Tussock Meadow (TM; 29.5 ha), Dryas Herb Mat (DH; 4.1 ha), and Dry Carex-Lichen (CL; 3.9 ha) ecotypes. The site was mainly closed during the bird breeding season from 2013 to 2014 apart from basic care and maintenance, the occurrence of environmental compliance programs, and for exploration programs, but was re-opened for construction and underground mine development in 2015 after the mines’ acquisition by TMAC Resources Inc. in 2013. For this study, anthropogenic disturbance was split into two categories: potential effect or non-effect years. Potential effect years were those with active construction and/or operations (2007–2012 and 2015) which included active building of site infrastructure, digging mine portal access, hauling ore, and blasting, which were activities conducted daily or extensively throughout the breeding bird season and which are typically associated with high levels of noise and movement. Non-effect years were those with no construction or operations (2013–2014) and only included care and maintenance of existing infrastructure, basic project compliance and mandatory monitoring (e.g. water quality testing), and some deposit exploration.
Avian surveys
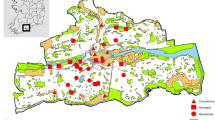
Between 2007 and 2015, we conducted avian surveys using two common methods for assessing avian abundance: point counts (Hutto et al. 1986; Ralph et al. 1995) and Program for Regional and International Shorebird Monitoring (PRISM) rapid surveys (Fig. 1; Skagen et al. 2003; Bart and Johnston 2012). Point count locations were established along transects radiating out from current and future planned infrastructure as part of the ongoing monitoring program at the mine. Additional clusters of point count locations were also established throughout the study area to have a greater variety of distances from mine infrastructure. PRISM survey, which were part of a collaborative effort with the Canadian Wildlife Service (CWS) at the Hope Bay Project, were established using a stratified random sampling approach, covering a variety of habitats.

Location of point count (n = 786) and PRISM (n = 72) plots surveyed for breeding birds, and mine infrastructure present at the Hope Bay Project, Nunavut, Canada, 2007–2015
A total of 786 point counts at 450 survey locations were completed between June 11 and July 6 in 2007, 2009–2010, and 2013–2015, with an average of 131 surveys/year (range 64–226 surveys/year). A total of 72 PRISM surveys were completed at 44 survey locations between June 11 and July 9 in 2011–2015, with an average of 14 surveys/year (range 10–22 surveys/year). Point count and PRISM surveys were completed in varying years because of monitoring program and logistical limitations. For point count surveys, 39.8% of sites were surveyed once, 45.8% were surveyed twice, and 14.4% were surveyed three times. For PRISM surveys, 45.5% of sites were surveyed once, 45.5% were surveyed twice, and 9.0% were surveyed three times. No survey locations were re-surveyed within the same year, only between years. Point count surveys were conducted between 0.0 and 11.4 km from the mine site (average distance = 2.58, SD = 2.50), and PRISM surveys were conducted between 0.1 and 20.0 km from the mine site (average distance = 6.69, SD = 6.39). As both survey methods had been completed for the project’s monitoring programs, we chose to use both datasets to maximize sample size and the inclusion of both songbirds and shorebirds.
Point counts (Hutto et al. 1986; Ralph et al. 1995) consisted of a 5-min stationary survey of all birds seen or heard within a 50 m radius of the observer. Point counts from 2013 onwards were conducted using a 100 m radius survey area. The observer estimated distances to all birds seen or heard during point counts conducted within the 100 m point counts. For consistency, we only used birds recorded between 0 to 50 m of the observer for all survey years. Point counts were spaced a minimum of 300 m apart from other point count centers to avoid double-counting of individual birds. Point counts were conducted between 7:30 am and 12:30 pm. The observer waited 2 min after arriving at point count locations prior to commencing the 5-min point count to allow bird activity to settle.
PRISM surveys (Skagen et al. 2003; Bart and Johnston 2012) involved two observers walking along sets of parallel lines through a 12-ha plot (300 m × 400 m) and recording all birds seen or heard within the plot boundaries (Skagen et al. 2003). For ease of navigation, PRISM plots were located with the longer side (400 m) oriented north–south. Observers systematically surveyed the plot area starting from one corner and walking in tandem along north–south transects at a distance of 25 m from one another. At the end of each 400 m long transect, each observer walked 50 m to their next transect line. This was repeated until each observer had walked six transect lines, for a total of 12 transect lines per plot. When conducting the surveys, observers used handheld Garmin GPS units to navigate along transect lines. The observers were not considered independent and mapped all birds within the plot and summarized results on a single data sheet by communicating during and after the survey.
For both survey methods, birds seen flying over the survey area or birds observed before or after surveys were completed were not included. Surveys were only conducted in fair weather, i.e., when wind speeds were < 25 km/h and temperatures were above 0 °C. Distance of all survey locations from project infrastructure was calculated in ArcGIS 10.4. For PRISM plot locations, the center of the plot was used to determine the distance to infrastructure.
Ecotype mapping
To determine ecotypes within the study area, terrestrial ecosystem mapping (TEM) was completed in 2010 for 56,138 ha of the Hope Bay region. TEM is the process of using ecological features such as vegetation, soil, and terrain to delineate ecotype units on a map within terrain polygons (Resources Inventory Committee 2000). Each polygon contains a maximum of three ecotypes from one to three deciles with the first decile being the most dominant ecotype unit and the third being the least dominant.
Statistical analyses
All analyses were completed using R 4.2.1 (R Core Team 2022). Analyses were completed separately for both PRISM and point count data, including investigations of total bird abundance (PRISM and point counts), songbird abundance (PRISM and point counts), and shorebird abundance (PRISM). Analyses considered temporal, and spatial variables, as well as distance to infrastructure and anthropogenic disturbance. There were a total of 25 ecotypes identified from the TEM data (Online Resource 1) and the proportion of each ecotype was calculated within the PRISM and point count spatial areas.
To reduce the number of variables considered in modelling, we clustered the PRISM and point count plots into groups with similar ecotype compositions using hierarchical clustering, as implemented in the vegan package (Oksanen et al. 2022). Distance among plots was calculated using Bray–Curtis dissimilarity, and clustering was performed using Ward linkage. We determined the optimal number of clusters using a combination of average silhouette width, and Pearson gamma index. Point count locations clustered into 5 groups and PRISM locations clustered into 4 groups (Online Resource 2). These groups were labelled based on the most abundant ecotype or ecotypes of the plots in each group. The 5 point count ecotype groups consisted of BL (betula-ledum-lichen), DH/RO (dryas herb mat/rock outcrop), DW (dry willow), TM (eriophorum tussock meadow), and WM/TM (wet meadow/eriophorum tussock meadow). The 4 PRISM ecotype groups were Mixture (a general mix of many ecotypes), PG (polygonal ground), WM/DH (wet meadow/dryas herb mat), and WM/TM (wet meadow/eriophorum tussock meadow).
Climate variables for the study area included NDVI as a measure of annual climatic variation, wind speed, and air temperature. Wind speed and average air temperature were taken from the hourly Doris Camp weather station for each survey day and averaged over the hours of 0900 and 1200. Using the MOD13A3 v061 (MODIS/Terra Vegetation Indices Monthly L3 Global 1 km SIN Grid V061; Didan 2021), NDVI tiles were downloaded for the month of June for each year of the study (2007–2015). A central point in the study area was chosen to extract the annual NDVI values, ensuring that the 1 km tile in which the extraction point was located was terrestrial habitat and representative of the study area. Bilinear interpolation was used to extract a single average value for June annually using the 9 tiles surrounding the central point. Low NDVI corresponds to sparser vegetation and higher NDVI corresponds to denser vegetation. In the Arctic, it will represents snowmelt or areas of barren rock and green up of vegetation once the snow recedes.
Avian abundance
The objective of the analysis was to determine if distance from infrastructure had a differential effect on breeding bird relative abundance in potential effect versus non-effect years (anthropogenic disturbance). All available habitat and environmental variables were controlled for in the model rather than implementing model selection. Thus, even if a covariate effect was not significant, the covariate was included in the model to control for extraneous variation. However, we did use AIC to determine if adding an interaction term between distance and anthropogenic disturbance significantly increased the fit of the model, as interactions in gamms with smooth terms can be difficult to interpret.
We modelled distance from infrastructure as a continuous variable using negative binomial mixed effects generalized additive models as implemented in the mgcv package (Wood et al. 2016). We allowed for both random slopes and intercepts by year. We included linear terms for NDVI (scaled to unit variance), average windspeed, average temperature, and a categorical term for ecotype cluster. An interaction term between distance to infrastructure and potential effect vs non-effect year was included to assess the differential effect of distance to infrastructure on relative abundance in potential effect years relative to non-effect years. A flat trajectory in the regression spline (and associated confidence intervals) over distance from infrastructure was an indication that there was no relationship between distance to infrastructure and relative abundance (i.e., relative abundance does not either increase or decrease with increasing distance to infrastructure, it remains constant across all distances). A non-flat trajectory of regression spline is an indication that there was a relationship between distance to infrastructure and relative abundance. Model fit was assessed using Akaike’s Information Criterion (AIC) with the correction for finite sample sizes (AICc). Specifically for the interaction between distance as a continuous variable and potential effect, models with smaller AICc values were interpreted to efficiently balance model complexity with the ability to describe the relationships in the data and allow for interpretation of whether the interaction term substantially increased the fit of the model justifying its inclusion. Model assumptions for all models were assessed using diagnostic plots of model residuals.
Results
A total of 20 breeding bird species, excluding unidentified species groupings, were recorded: 12 songbird, 6 shorebird, and 2 ptarmigan species. The most frequently encountered species during point counts were Savannah Sparrow (Passerculus sandwichensis), Lapland Longspur (Calcarius lapponicus), Hoary Redpoll (Acanthis hornemanni), Horned Lark (Eremophila alpestris), American Tree Sparrow (Spizelloides arborea), White-crowned Sparrow (Zonotrichia leucophrys), and Redpoll sp. (Acanthis sp.). The most common species recorded during PRISM surveys were Savannah Sparrow, Lapland Longspur, Redpoll sp., American Tree Sparrow, and Horned Lark. The most commonly recorded shorebirds for PRISM surveys were Least Sandpiper (Calidris minutilla), Red-necked Phalarope (Phalaropus lobatus), and Pectoral Sandpiper (Calidris melanotos). PRISM surveys resulted in a greater amount of shorebird species counted compared to point count surveys, allowing for guild-specific results (Table 1; Online Resource 3).
Point counts
For all of the point count bird groups, which included total bird and songbird abundance, the model with no-interaction term between distance to infrastructure and potential effect/non-effect year was a nearly identical fit to the data as one with an interaction term (AICc = 2321.31 vs. 2321.21 for total birds, and 2300.25 vs. 2299.95 for songbirds), favoring the less complicated no-interaction model. We found no significant difference between potential effect and non-effect years on total bird abundance. Conversely, distance from infrastructure was significantly associated with total bird abundance (edf = 1.26, p = 0.0001; Table 2) with a weak increase in bird abundance linearly with distance from infrastructure (Fig. 2a). Holding all other continuous covariates constant at their mean, ecotype cluster at TM, and year in 2007: at 500 m the model predicts a bird abundance of 1.3 in non-effect years and 1.3 in potential effect years, these estimates increase to 1.6 and 1.7 at 4 km, respectively, and 2.1 and 2.2 at 10 km. There was also a significant positive linear association between bird abundance and air temperature (p < 0.001; Table 2), and a negative relationship with NDVI (p = 0.007; Table 2). There was significantly lower abundance in DH/RO ecotype plots compared to BL (RR = 0.74; 95% CI 0.56–1.00, p = 0.0470), and WM/TM compared to BL (RR = 0.72; 95% CI 0.54–0.97, p = 0.0286; Table 2). When completing the analysis using only songbirds, identical significant variables were present (Table 2). The analysis was not completed for shorebirds or ptarmigans as the sample size was too low.

Model predicted abundance from generalized additive mixed model results for point count a total breeding birds and PRISM b total breeding birds, c songbirds, and d shorebirds. The point count songbird abundance trend was nearly identical to that of a and is therefore not illustrated. Abundance significantly increased with distance to infrastructure for all groups apart from PRISM songbirds (c), although shorebirds illustrated a unimodal pattern. Shaded areas represent 95% confidence intervals
PRISM
For PRISM total bird abundance, the model with no-interaction term between distance to infrastructure and potential effect/non-effect year was the best fit to the data (AICc = 501.50 vs 505.39 for interaction model). We found no significant difference between potential effect and non-effect year on total bird abundance. Conversely, distance from infrastructure had a significant non-linear relationship with bird abundance (edf = 1.52, p = 0.0171; Table 2) with a gradual increase in bird abundance to 10 km, followed by a shallow decline (Fig. 2b). At 500 m, the model predicted a bird abundance of 10.9 in non-effect years and 12.7 in potential effect years. These estimates increase to 15.5 and 18.1 at 10 km, respectively. There was significantly higher abundance in WM/DH ecotype plots compared to Mixture plots (RR = 2.17, 95% CI 1.38–3.41, p = 0.0008; Table 2).
When analyzing songbirds only, the AIC value was nearly identical for the non-interaction model, therefore, the less complicated no-interaction term model was preferred (AICc = 481.15 vs 481.53). We found no significant difference between potential effect and non-effect years and no relationship between songbird abundance with distance from infrastructure (Table 2, Fig. 2c). Comparable to overall bird abundance, there was higher abundance in WM/DH ecotypes and PG compared to Mixture ecotypes (RR = 1.94; 95% CI 1.23–3.06, p = 0.0046; Table 2).
For the shorebird models, the AIC value was lower for the no-interaction model, therefore, the less complicated no-interaction term model was preferred (AICc = 249.56 vs 254.69). Shorebird abundance was not significantly associated with effect years. Distance from infrastructure had a significant non-linear association with shorebird abundance (edf = 2.18, p = 0.0005; Table 2) with an increase in the number of birds out to approximately 10 km from infrastructure followed by a decline out to 20 km (Fig. 2d). Similarly to songbirds, there was higher shorebird abundance in WM/DH ecotypes compared to Mixture ecotypes (RR = 9.86; 95% CI 2.22–43.80, p = 0.0027; Table 2).
Discussion
Our study revealed mixed impacts of proximity to infrastructure on overall breeding bird and guild-specific abundance. Although our prediction was partially supported, we did not find as clear of a distance effect threshold as predicted, which has been observed in certain studies (Bayne et al. 2008; Liebezeit et al. 2009; Benítez-López et al. 2010). For point counts surveys, total breeding bird and songbird abundance slightly increased with distance to infrastructure, regardless of effect years. Total upland birds in PRISM surveys showed a significant increased out to 10 km with a shallow decline past this distance. PRISM songbird abundance did not have a significant relationship with distance to infrastructure, which could be due to an artifact of low sample size for PRISM surveys at close proximity to infrastructure. PRISM shorebirds showed a unimodal relationship with distance, with a peak in abundance at approximately 10 km from infrastructure (Fig. 2, Table 2). The slight variability in trends for total bird abundance between point count and PRISM results may be because PRISM surveys were overly dispersed and of too small a sample size compared to point count surveys, in addition to variability in influence of different bird guilds, with the number of shorebirds observed being higher for PRISM compared to point counts (Table 1). As opposed to our predictions, anthropogenic disturbance did not influence breeding bird abundance for either survey type or bird guild. Although anthropogenic disturbances such as noise have been recorded to have significant impacts (Van Der Zande et al. 1980; Bayne et al. 2008), some studies suggest that distance to infrastructure may be a more crucial variable for breeding birds (Bernath-Plaisted and Koper 2016).
Our findings are consistent with other studies demonstrating an impact of natural resource industry infrastructure on avian populations. Hoary Redpolls, American Tree Sparrows, and Savannah Sparrows, species commonly recorded during our study, were recorded at higher densities within control plots away from a Canadian diamond mine (Smith et al. 2010). Grassland and sagebrush songbirds in western North America declined in abundance near oil infrastructure, irrespective of anthropogenic noise presence (Gilbert and Chalfoun 2011; Mutter et al. 2015). Similarly, forest birds experienced declines near shale pad edges (Barton et al. 2016). Previous studies have also associated renewable energy infrastructure with the displacement of breeding birds (Pearce‐Higgins et al. 2009; Sansom et al. 2016; Fernández‐Bellon et al. 2019). Our study reflects, in part, a global trend of decreasing bird population densities as proximity to infrastructure increases (Benítez-López et al. 2010).
While we found an overarching decrease of breeding bird abundance as proximity to infrastructure increases (Fig. 2), many studies have observed mixed effects of development when evaluating individual species. Typically, habitat generalists and synanthropic (i.e., human-associated) species are documented to increase in abundance near various energy sector developments and associated anthropogenic disturbances (Barton et al. 2016; Bayne et al. 2016; Farwell et al. 2019). Certain species were found to be more common on mine plots rather than control plots at an Arctic diamond mine (Smith et al. 2005). At Baker Lake in the Canadian Arctic, several shorebirds and songbirds were positively associated with towns, roadways, and gravel pits, and did not reveal lower bird densities near a mine site (Staniforth 2002). Similarly, certain species increased or showed no response to oil and natural gas development (Gilbert and Chalfoun 2011; Hamilton et al. 2011). As we did not account for species-specific trends, certain breeding birds at Hope Bay may demonstrate inverse or neutral relationships in comparison to our overall findings. For example, Horned Lark, Semipalmated Plover (Charadrius semipalmatus), and White-crowned Sparrow, which were species observed during our study (Online Resource 3), were recorded at significantly higher densities within mine plots compared to control plots at a diamond mine in the Northwest Territories (Smith et al. 2005).
Our results illustrate that habitat (in this case measured by ecotype), in addition to mine infrastructure, plays an important role in breeding bird abundance on the landscape surrounding the Hope Bay Project. For point counts, the DH/RO (dry upland ecotypes) and WM/TM (moist to wet lowland ecotypes) groups had significantly lower abundance compared to the BL grouping (dry to mesic upland ecotypes; Table 2; Online Resource 1). For PRISM surveys, the WM/DH grouping, primarily associated with wet lowland ecotypes, was positively associated with total breeding bird and shorebird abundance (Table 2; Online Resource 1). PRISM songbird abundance was positively associated with both WM/DH and PG ecotypes, which are a combination of mixed dry to wet lowland and upland ecotypes. Arctic-breeding birds, particularly shorebirds, are typically associated with wetland and semi-wetland habitats (Latour et al. 2005; Liebezeit et al. 2011; Jóhannesdóttir et al. 2014; Cunningham et al. 2016; Hawkshaw et al. 2021) with habitat suitability decreasing at higher elevation (Saalfeld et al. 2013). Higher wetland density and wet habitat on the landscape has also been positively associated with shorebird density and richness (Brown et al. 2007; Martin et al. 2020). Arctic-breeding birds are not only abundant in these ecotypes, but also preferentially nest in wet and emergent habitats (Liebezeit et al. 2011). This abundantly recorded trend is reflected in our PRISM findings of higher avian abundance, particularly shorebirds, in ecotypes associated with wet lowland habitats. In contrast, the mixed habitat relationships for point counts and PRISM songbirds may be due to the lower number of shorebirds observed for point counts, which are associated with wetter habitats, and songbird species-specific habitat preferences. As species such as the Horned Lark and Lapland Longspur, which are associated with drier upland or mixed habitats (Latour et al. 2005), accounted for a large portion of birds observed for both survey methods, this may explain variability in habitat associations.
In addition to ecotype, the timing of snow melt has also been recorded to have a strong impact on Arctic bird nesting phenology (Smith et al. 2010; Liebezeit et al. 2014) and habitat selection (Cunningham et al. 2016). We used the NDVI, which broadly illustrates green up in the Arctic, as an indicator of annual weather patterns, since we did not have snow melt data available, and found a significant negative relationship between NDVI and point count total bird and songbird abundance (Table 2). This finding, along with the positive significant association with higher air temperature, may be related to detection probability rather than true bird abundance. In years where green up occurred earlier, associated with earlier snowmelt, birds may exhibit earlier breeding initiation and become less detectable as a result (Best and Peterson 1985; Wilson and Bart 1985). Several studies have recorded clutch initiation advancement in association with earlier snow melt (Smith et al. 2010; Liebezeit et a. 2014; Saalfeld and Lanctot 2017). A decrease in detectability related to timing within the breeding season is particularly important for surveys, such as point counts, which rely more heavily on auditory activity (i.e., singing and displaying males) compared to PRISM surveys where surveyors are actively moving throughout the plot area and have a likelihood of encountering birds through direct flushing of incubating birds or general disturbance.
Accounting for detection probability is a limitation of our study as surveys were not repeated within years due to logistical and monitoring program constraints. Repeated surveys within and between years would allow for measures of detection probability and a more precise account of mine and environmental effects on bird abundance. Particularly, the unimodal trend of shorebirds with distance to infrastructure would benefit from technical and biological replicates to ensure that this trend reflects biological reality. Although we did not detect an impact of anthropogenic disturbance during this study, the classification was coarse (potential effect or non-effect years). Future studies should consider more specific variables relating to anthropogenic disturbances such as noise, vehicle movements, or lighting schemes which may provide a more detailed understanding of the influence of potential anthropogenic disturbances on avian populations at Arctic industrial developments. Additionally, avian abundance may be a misrepresentative indicator of adverse anthropogenic impacts. Other factors to consider for future studies are reproductive success, survival, and nest and territory establishment in relation to infrastructure (Male and Nol 2005; Shochat et al. 2005; Liebezeit et al. 2009; Kendall 2011; Ludlow et al. 2015; Bernath-Plaisted and Koper 2016). Moreover, the influence of infrastructure on predator abundance may indirectly impact nesting timing and breeding bird predation risk (Truett et al. 2006; Meltofte et al. 2007; Liebezeit et al. 2009; Smith et al. 2010; Mckinnon et al. 2013). The relationship between nest success and infrastructure can also vary between bird guilds, and show further interspecific discrepancies (Liebezeit et al. 2009).
Breeding bird populations have decreased across the Arctic (Smith et al. 2020) while stressors of climate-change intensify (Wauchope et al. 2017), and improved resource accessibility drive development across the Arctic (Kondratiev 2020). It is increasingly important for industry developments, such as mine sites, to understand impacts on avian populations for effective mitigation, management, or compensation programs (Tolvanen et al. 2019). The findings from our study at an underground gold mine in the Canadian Arctic offer valuable insight into the relationship between industrial development and breeding bird abundance. Despite the anticipated impacts of mine development effects (i.e., distance to infrastructure and anthropogenic disturbance), our results reveal that distance from infrastructure and habitat type are significant determinants of bird abundance, overshadowing anthropogenic disturbance measured at this scale. This underscores the resilience of Arctic-breeding birds to a certain extent, while highlighting the crucial role of preserving suitable breeding habitats to support Arctic bird populations. By planning industrial developments around preferred breeding habitats and implementing and engaging in strategic monitoring and management, mining and other industries have the potential to minimize their environmental footprint and actively contribute to the protection and understanding of Arctic-breeding birds. Incorporating repeated surveys and expanding PRISM data collection in future studies will enable a more dynamic understanding of avian responses to industrial development to ensure the coexistence of Arctic development and avian populations.
Data availability
The datasets generated and analyzed during the study, and the code required to reproduce the results in the manuscript are publicly available in Zenodo (https://doi.org/10.5281/zenodo.8270783).
References
Bart JR, Johnston VH (2012) Arctic shorebirds in North America: a decade of monitoring. University of California Press, Berkeley
Barton EP, Pabian SE, Brittingham MC (2016) Bird community response to Marcellus shale gas development: shale gas development and forest birds. J Wildl Manag 80:1301–1313. https://doi.org/10.1002/jwmg.21117
Bayne EM, Habib L, Boutin S (2008) Impacts of chronic anthropogenic noise from energy-sector activity on abundance of songbirds in the boreal forest. Conserv Biol 22:1186–1193. https://doi.org/10.1111/j.1523-1739.2008.00973.x
Bayne E, Leston L, Mahon CL et al (2016) Boreal bird abundance estimates within different energy sector disturbances vary with point count radius. The Condor 118:376–390. https://doi.org/10.1650/CONDOR-15-126.1
Benítez-López A, Alkemade R, Verweij PA (2010) The impacts of roads and other infrastructure on mammal and bird populations: a meta-analysis. Biol Conserv 143:1307–1316. https://doi.org/10.1016/j.biocon.2010.02.009
Bernath-Plaisted J, Koper N (2016) Physical footprint of oil and gas infrastructure, not anthropogenic noise, reduces nesting success of some grassland songbirds. Biol Conserv 204:434–441. https://doi.org/10.1016/j.biocon.2016.11.002
Best LB, Petersen KL (1985) Seasonal changes in detectability of Sage and Brewer’s sparrows. The Condor 87(4):556–558. https://doi.org/10.2307/1367964
Brown S, Bart J, Lanctot RB et al (2007) Shorebird abundance and distribution on the coastal plain of the Arctic National Wildlife Refuge. The Condor 109:1–14. https://doi.org/10.1093/condor/109.1.1
Cunningham JA, Kesler DC, Lanctot RB (2016) Habitat and social factors influence nest-site selection in Arctic-breeding shorebirds. Auk 133:364–377. https://doi.org/10.1642/AUK-15-196.1
Didan K (2021) MODIS/Terra vegetation indices monthly L3 Global 1km SIN Grid V06 distributed by NASA EOSDIS Land Processes Distributed Active Archive Center. https://doi.org/10.5067/MODIS/MOD13A3.061
Farwell LS, Wood PB, Brown DJ, Sheehan J (2019) Proximity to unconventional shale gas infrastructure alters breeding bird abundance and distribution. The Condor. https://doi.org/10.1093/condor/duz020
Fernández-Bellon D, Wilson MW, Irwin S, O’Halloran J (2019) Effects of development of wind energy and associated changes in land use on bird densities in upland areas. Conserv Biol 33:413–422. https://doi.org/10.1111/cobi.13239
Francis CD, Ortega CP, Cruz A (2009) Noise pollution changes avian communities and species interactions. Curr Biol 19:1415–1419. https://doi.org/10.1016/j.cub.2009.06.052
Gilbert MM, Chalfoun AD (2011) Energy development affects populations of sagebrush songbirds in Wyoming. J Wildl Manag 75:816–824. https://doi.org/10.1002/jwmg.123
Gregory RD, Strien AV (2010) Wild bird indicators: using composite population trends of birds as measures of environmental health. Ornithol Sci 9:3–22. https://doi.org/10.2326/osj.9.3
Hamilton LE, Dale BC, Paszkowski CA (2011) Effects of disturbance associated with natural gas extraction on the occurrence of three grassland songbirds. Avian Conserv Ecol. https://doi.org/10.5751/ACE-00458-060107
Hawkshaw KA, Foote L, Franke A (2021) Ecological determinants of avian distribution and abundance at Rankin Inlet, Nunavut in the Canadian Arctic. Polar Biol 44:1–15. https://doi.org/10.1007/s00300-020-02766-4
Hutto RL, Pletschet SM, Hendricks P (1986) A fixed-radius point count method for nonbreeding and breeding season use. Auk 103:593–602. https://doi.org/10.1093/auk/103.3.593
Jóhannesdóttir L, Arnalds Ó, Brink S, Gunnarsson TG (2014) Identifying important bird habitats in a sub-arctic area undergoing rapid land-use change. Bird Study 61:544–552. https://doi.org/10.1080/00063657.2014.962481
Johnston VH, Gratto-Trevor CL, Pepper ST (2000) Assessment of bird populations in the Rasmussen Lowlands, Nunavut. Can Wildl Serv 101:1–58
Kendall S (2011) Impacts of climate change and development on shorebirds of the Arctic national wildlife refuge. In: Gyrfalcons and ptarmigan in a changing world. The Peregrine Fund, Boise
Kondratiev VB (2020) Mineral resources and future of the Arctic. Min Ind J. https://doi.org/10.30686/1609-9192-2020-1-87-96
Kullerud L (2011) A green Arctic. Nat 478:179–180. https://doi.org/10.1038/478179a
Latour PB, Machtans CS, Beyersbergen GW (2005) Shorebird and passerine abundance and habitat use at a high Arctic breeding site: Creswell Bay, Nunavut. Arct 58:55–65. https://doi.org/10.14430/arctic389
Liebezeit JR, Kendall SJ, Brown S et al (2009) Influence of human development and predators on nest survival of tundra birds, Arctic Coastal Plain, Alaska. Ecol Appl 19:1628–1644. https://doi.org/10.1890/08-1661.1
Liebezeit JR, White GC, Zack S (2011) Breeding ecology of birds at Teshekpuk lake: a key habitat site on the Arctic Coastal Plain of Alaska. Arct. https://doi.org/10.14430/arctic4078
Liebezeit JR, Gurney KEB, Budde M et al (2014) Phenological advancement in arctic bird species: relative importance of snow melt and ecological factors. Polar Biol 37:1309–1320. https://doi.org/10.1007/s00300-014-1522-x
Ludlow SM, Brigham RM, Davis SK (2015) Oil and natural gas development has mixed effects on the density and reproductive success of grassland songbirds. The Condor 117:64–75. https://doi.org/10.1650/CONDOR-14-79.1
Male SK, Nol E (2005) Impacts of roads associated with the Ekati Diamond Mine TM, Northwest Territories, Canada, on reproductive success and breeding habitat of Lapland Longspurs. Can J Zool 83:1286–1296. https://doi.org/10.1139/z05-128
Martin EC, Jochum KA, Bagley CF, Doherty PF Jr (2020) Shorebird abundance estimates in interior Alaska. Wildlife Manag. https://doi.org/10.1002/jwmg.21913
Mckinnon L, Berteaux D, Gauthier G, Bêty J (2013) Predator-mediated interactions between preferred, alternative and incidental prey in the arctic tundra. Oikos 122:1042–1048. https://doi.org/10.1111/j.1600-0706.2012.20708.x
Meltofte H, Høye TT, Schmidt NM, Forchhammer MC (2007) Differences in food abundance cause inter-annual variation in the breeding phenology of High Arctic waders. Polar Biol 30:601–606. https://doi.org/10.1007/s00300-006-0219-1
Morrison ML, Block WM, Strickland MD, Collier BA, Peterson MJ (2008) Wildlife study design. Springer, New York
Mutter M, Pavlacky DC, Van Lanen NJ, Grenyer R (2015) Evaluating the impact of gas extraction infrastructure on the occupancy of sagebrush-obligate songbirds. Ecol Appl 25:1175–1186. https://doi.org/10.1890/14-1498.1
National Research Council (2003) Cumulative environmental effects of oil and gas activities on Alaska’s North Slope. The National Academies Press, Washington
Oksanen J, Simpson G, Blanchet F et al (2022) vegan: Community Ecology Package
Pearce-Higgins JW, Stephen L, Langston RHW et al (2009) The distribution of breeding birds around upland wind farms. J Appl Ecol 46:1323–1331. https://doi.org/10.1111/j.1365-2664.2009.01715.x
R Core Team (2022) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Ralph CJ, Sauer JR, Droege S (1995) Monitoring bird populations by point counts. U.S. Department of Agriculture, CA
Resources Inventory Committee (2000) Standard for digital terrestrial ecosystem mapping (TEM) data capture in British Columbia: Ecosystem Technical Standards and Database Manual
Robinson BG, Franke A, Derocher AE (2014) The influence of weather and lemmings on spatiotemporal variation in the abundance of multiple avian guilds in the Arctic. PLoS ONE. https://doi.org/10.1371/journal.pone.0101495
Saalfeld ST, Lanctot RB (2015) Conservative and opportunistic settlement strategies in Arctic-breeding shorebirds. Auk 132:212–234. https://doi.org/10.1642/AUK-13-193.1
Saalfeld ST, Lanctot RB (2017) Multispecies comparisons of adaptability to climate change: a role for life history characteristics? Ecol Evol 7:10492–10502. https://doi.org/10.1002/ece3.3517
Saalfeld ST, Lanctot RB, Brown SC et al (2013) Predicting breeding shorebird distributions on the Arctic Coastal Plain of Alaska. Ecosphere. https://doi.org/10.1890/ES12-00292.1
Saalfeld ST, McEwen DC, Kesler DC, Butler MG, Cunningham JA, Doll AC, English WB, Gerik DE, Grond K, Herzog P, Hill BL, Lagassé BJ, Lanctot RB (2019) Phenological mismatch in Arctic-breeding birds: impact of snow melt and unpredictable weather conditions on food availability and chick growth. Ecol Evol 9:6693–6707. https://doi.org/10.1002/ece3.5248
Samelius G, Alisauskas RT, Hobson KA, Larivière S (2007) Prolonging the arctic pulse: long-term exploitation of cached eggs by arctic foxes when lemmings are scarce. J Anim Ecol 76:873–880. https://doi.org/10.1111/j.1365-2656.2007.01278.x
Sansom A, Pearce-Higgins JW, Douglas DJT (2016) Negative impact of wind energy development on a breeding shorebird assessed with a BACI study design. Ibis 158:541–555. https://doi.org/10.1111/ibi.12364
Shochat E, Patten MA, Morris DW et al (2005) Ecological traps in isodars: effects of tallgrass prairie management on bird nest success. Oikos 111:159–169. https://doi.org/10.1111/j.0030-1299.2005.13907.x
Skagen SK, Bart J, Andres B, Brown S, Donaldson G, Harrington B, Johnston V, Jones SL, Morrison RIG (2003) Monitoring the shorebirds of North America: towards a unified approach. Wader Study Group Bull 100:102–104
Smith AC, Virgl JA, Panayi D, Armstrong AR (2005) Effects of a diamond mine on Tundra-breeding birds. Arct 58:295–304. https://doi.org/10.14430/arctic431
Smith PA, Gilchrist HG, Forbes MR et al (2010) Inter-annual variation in the breeding chronology of arctic shorebirds: effects of weather, snow melt and predators. J Avian Biol 41:292–304. https://doi.org/10.1111/j.1600-048X.2009.04815.x
Smith PA, McKinnon L, Meltofte H et al (2020) Status and trends of tundra birds across the circumpolar Arctic. Ambio 49:732–748. https://doi.org/10.1007/s13280-019-01308-5
Staniforth RJ (2002) Effects of urbanization on bird populations in the Canadian Central Arctic. Arct 55:87–93. https://doi.org/10.14430/arctic693
Tolvanen A, Eilu P, Juutinen A et al (2019) Mining in the Arctic environment—a review from ecological, socioeconomic and legal perspectives. J Environ Manag 233:832–844. https://doi.org/10.1016/j.jenvman.2018.11.124
Truett JC, Senner RGB, Kertell K et al (2006) Wildlife responses to small-scale disturbances in Arctic Tundra. Wildl Soc Bull 22:317–324
Van Der Zande AN, Ter Keurs WJ, Van Der Weijden WJ (1980) The impact of roads on the densities of four bird species in an open field habitat—evidence of a long-distance effect. Biol Conserv 18:299–321. https://doi.org/10.1016/0006-3207(80)90006-3
Wauchope HS, Shaw JD, Varpe Ø et al (2017) Rapid climate-driven loss of breeding habitat for Arctic migratory birds. Glob Change Biol 23:1085–1094. https://doi.org/10.1111/gcb.13404
Wauchope HS, Amano T, Geldmann J, Johnston A, Simmons BI, Sutherland WJ, Jones JPG (2021) Evaluating impact using time-series data. Trends Ecol Evol. https://doi.org/10.1016/j.tree.2020.11.001
Wilson DF, Bart J (1985) Reliability of singing bird surveys: effects of song phenology during the breeding season. The Condor 87(1):69–73. https://doi.org/10.2307/1367132
Wood SN, Pya N, Säfken B (2016) Smoothing parameter and model selection for general smooth models. J Am Stat Assoc 111:1548–1563. https://doi.org/10.1080/01621459.2016.1180986
Acknowledgements
We would like to thank the many people who contributed to field studies and data management over the years. This includes but is not limited to (in alphabetical order): Christine Kent, Julia Shewan, Leslie Bol, Laurie Ainsworth, and Oliver Curran. We also give our thanks to Nancy Duquet-Harvey who provided suggestions and reviewed the manuscript.
Funding
This work was primarily funded by owners and operators of the Hope Bay site, including TMAC Inc. and Agnico Eagle Mines Ltd. Funding included field travel and accommodations, as well as financial support for statistical analyses, consultation and discussion with stakeholders, and preparation of the manuscript. Additional support was provided through ERM Canada Ltd.
Author information
Authors and Affiliations
Contributions
GS conceived the research design. GS and HV collected data and/or supervised field collection. GS, HV, and ALM conceived the research idea and questions. AA analyzed the data. ALM wrote the paper, and HV and GS edited the paper. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Competing interests
All years of data collection were carried out as part of compliance monitoring for potential effects at the Hope Bay Project, with feedback and oversight from the Government of Nunavut, Environment and Climate Change Canada, and the Canadian Wildlife Service.
Ethical approval
Surveys were conducted with annual General Wildlife permits from the Government of Nunavut. All work for this project was non-invasive and did not require animal care permits.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Merondun, A.L., Albert, A., Visty, H. et al. Proximity to infrastructure and ecotype influence breeding bird abundance at an Arctic mine, the Hope Bay Project, Nunavut, Canada. Polar Biol 47, 475–486 (2024). https://doi.org/10.1007/s00300-024-03244-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-024-03244-x