
Abstract
Burdwood Bank is a shallow seamount located south of the Malvinas (Falkland) Islands and east of Tierra del Fuego. In 2013, the area down to 200 m depth of this bank was declared the first open-sea (non-coastal) Marine Protected Area (MPA) in Argentina, and named “Namuncurá” (MPAN-BB). The present study aimed to evaluate the biodiversity of the isopods and cumaceans from the MPAN-BB. The studied material was collected on board the Argentine RV Puerto Deseado during the Campaña Antártica de Verano 2013. Three stations from the MPAN-BB, respectively, located in the core, buffer, and transition sectors, were sampled. A total of 17,076 specimens were sorted and 70 taxa distributed in 22 families were identified. Thirty-five new records of distribution and eight species most probably new to science are herein reported. Paramunnidae and Diastylidae were the most abundant and diverse families among isopods and cumaceans, respectively. Our records, together with those obtained from the literature, make a total of 55 nominal species so far known from Burdwood Bank. Except for one nominal species, which had been reported from the Puerto Rico Trench and the Argentine Basin, all the remaining species had also been recorded from other localities within the Magellan Region. In contrast, only nine out of the 55 nominal species had also been reported from the Antarctic Peninsula and/or Scotia Arc. This indicates that the isopods and cumaceans from Burdwood Bank are typically Magellanic, and there is little correspondence between this fauna and that from the Antarctic Peninsula and Scotia Arc.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Burdwood Bank (55°S, 59°W) is a shallow seamount, 300 km long and 60 km wide, located about 200 km south of the Malvinas (Falkland) Islands and 150 km east of Isla de los Estados (Staten Island). This bank is part of the North Scotia Ridge, and along with the Davis and Aurora Banks, the Shag Rocks and the South Georgia Island form the northern boundary of the Scotia Sea (Pandey et al. 2010; Cavallotto et al. 2011; Torres Carbonell et al. 2014).
To the north, Burdwood Bank is surrounded by the Malvinas (Falkland) Channel, which extends in a west–east direction (Fig. 1). The depth of this channel increases from 400 to 3000 m toward the east. The bank is separated from the Patagonian shelf by a relatively narrow and shallow channel—80 km wide and 400 m deep—which connects the Scotia Basin with the Malvinas Channel. On its east margin, it is surrounded by another channel—130 km wide and as deep as 1800 m—which also connects the Scotia Basin with the Malvinas Channel (Guerrero et al. 1999). To the south, the bank extends along a complex and steep slope incised by a system of canyons (Harris and Whiteway 2011; Bozzano et al. 2017).

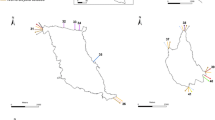
Stations sampled in the core (dark gray), buffer (dashed) and transition (light gray) sectors of the Marine Protected Area Namuncurá-Burdwood Bank (MPAN-BB)
Regarding the impact of Burdwood Bank on the oceanic circulation, Piola and Gordon (1989) reported that, upon encountering this bank, the Antarctic Circumpolar Current (ACC) splits into two branches, which flow around its margins. Matano et al. (2019) presented a suite of numerical simulations to characterize the circulation over Burdwood Bank, which showed energetic and persistent uplifting of deep waters in the area. These waters are potentially rich in micronutrients and might contribute to the fertilization of the upper layers of the southwestern Atlantic region, which is one of the most fertile areas of the Southern Ocean. However, Matano et al. (op. cit) stated that although the results of the model are robust, there is insufficient observational evidence to corroborate them.
Burdwood Bank is a hotspot of benthic diversity, including long-lived species with a very slow recovery time after perturbation events (Schejter et al. 2016, 2017b, 2020). To preserve this fragile ecosystem, in 2013, the Argentine government created the Marine Protected Area Namuncurá-Burdwood Bank (MPAN-BB). In addition, in 2018, the Argentine government established two other MPAs: the “Namuncurá-Burdwood Bank II” (MPAN-BB II), which extends the MPAN-BB southwards and to deeper waters, and the MPA “Yaganes”, which is located just off the southern tip of Tierra del Fuego, extending southwards into the Drake Passage.
Since 2013, an intensive sampling program of the benthic community at Burdwood Bank has been carried out on board the Argentine RV Puerto Deseado (surveys CAV 2013, BBB 2016, PD-BB-ABR 2017, among others). As a preliminary result of these surveys, Schejter et al. (2016) presented a checklist of benthic organisms (mainly mega and macrofauna). Later on, Schejter et al. (2017b) provided the first submarine images of the MPAN-BB, which revealed highly heterogeneous substrata in the area, and briefly described the faunas of two rocky environments and an area dominated by coarse biogenic sediments. More recently, Schejter et al. (2020) studied the megabenthic communities on the slope of Burdwood Bank and neighboring areas (≥ 200 m depth), reporting “coral gardens” mainly composed of alcyonaceans, as well as pennatulacean aggregations and communities dominated by sponges.
In addition, exhaustive species inventories, descriptions of new species, new records of distribution, and studies on reproductive biology and biogeography have been recently published on the fauna from Burdwood Bank (Güller and Zelaya 2017; Penchaszadeh et al. 2017, 2019; Schejter et al. 2017a; Fraysse et al. 2018, 2020; García Alonso et al. 2018; López Gappa et al. 2018; Taverna et al. 2018; Urcola and Zelaya 2018; Bremec et al. 2019; Di Luca and Zelaya 2019; Gordillo et al. 2019; Teso et al. 2019; Teso and Penchaszadeh 2019; Pérez-Barros et al. 2020; Roccatagliata 2020; among others).
Regarding the isopods and cumaceans, only a few species, mostly collected by foreign surveys (Swedish Antarctic Expedition 1901–1903, HMS William Scoresby 1926–1932, Soviet Antarctic Expedition 1955–1958, RV Vema 1961, RV Eltanin 1962, ANTARKTIS XIX/5 (LAMPOS) 2002), have been recorded from Burdwood Bank. Most recently, a few additional records also came from the Argentine expeditions CAV 2013 and BBB 2016. As a result of all these surveys, a total of 13 isopods and 8 cumaceans have been reported from the area (see Discussion section for citations). It should be noted that neither isopods nor cumaceans were reported by the Scottish National Antarctic Expedition 1902–1904, despite the exceptional abundant sample (“The largest and richest catch of the Expedition, totalling about half a ton.”) taken at Burdwood Bank during that survey (Wilton et al. 1908).
So far, no comprehensive study on the Peracarida from Burdwood Bank has been performed. The large amount of peracarids, including thousands of amphipods, isopods, cumaceans and tanaidaceans, recently collected by the RV Puerto Deseado in the MPAN-BB gives us the opportunity to achieve better knowledge about this fauna. As the first step in this direction, the isopods and cumaceans are herein investigated. In particular, the aims of this paper were: (1) to analyze the species composition of Isopoda and Cumacea in the MPAN-BB, (2) to report new species and new records of distribution, and (3) to discuss the faunal similarities between Burdwood Bank and its neighboring areas.
Study area
The MPAN-BB has a minimum depth of about 50 m and is circumscribed by the 200 m isobath. Three sectors were defined for managing purposes (see Falabella 2017): (1) the core sector, which is the central portion of the MPAN-BB and is under strict protection; (2) the buffer sector, which surrounds the core, and where extracting activities are allowed only for scientific research; and (3) the transition sector, which is the most external area of the MPAN-BB, and where productive and extractive activities contemplated in the Management Plan are allowed (Fig. 1).
Material and methods
The studied specimens were collected on board the RV Puerto Deseado during the Campaña Antártica de Verano (CAV) 2013. Three stations, one in each sector of the MPAN-BB (core, buffer and transition; Fig. 1, Table 1), were sampled using a Rauschert sledge (see Lörz et al. 1999) equipped with a net of 1 mm mesh size. On each station, the sledge was hauled on the ocean floor for about 10 min with the vessel moving at 1–2 knots. On deck, the specimens were sieved with a 250-μm mesh, and then fixed in 10% sea-water buffered formalin. Additionally, some specimens were preserved directly in 96% ethanol for later molecular studies. Once in the laboratory, the samples fixed in formalin were transferred to 70% ethanol and all the isopods and cumaceans were sorted by species under a stereoscopic microscope Leica MZ8.
Species were identified to the lowest taxonomic level possible. The species that are most probably new to science were provisionally designated with letters (e.g., “n. sp. A”) and will remain in open nomenclature until the formal description is published. Species identified with the abbreviation “cf.” (Latin confer, “compare”) means that most diagnostic characters correspond to the given species but some characters are unclear and, thus, that the identification is provisional. Chaetarcturus sp. A and B are two distinct species with unresolved taxonomic status. Finally, taxa identified to the genus or family levels (denoted with “sp.” and “indet.”, respectively) include only single species.
The Jaccard similarity index (JSI) was used to compare how similar a pair of stations was: the closer the value to 1, the greater the similarity between the stations analyzed.
All previous records of isopods and cumaceans from Burdwood Bank were compiled from the literature. Besides, the Ocean Biogeographic Information System (OBIS) was checked, and only species identified by a taxonomist with expertise in isopods or cumaceans were considered.
Although MPAN-BB is defined as the submerged plateau limited by the 200 m isobath, the specimens obtained up to 500 m depth by previous expeditions were taken into account in the Discussion section and are listed in Table 3.
For the zoogeographic analysis, we investigated the occurrence in the neighboring localities of the isopods and cumaceans reported from Burdwood Bank. A group-averaged cluster analysis and a non-metric multidimensional scaling analysis (MDS), both based on a species presence/absence matrix, were performed with the PRIMER v5.2.2 program (Clarke and Warwick 2001). The Bray–Curtis similarity index was used. The localities considered were the following: Burdwood Bank, the Pacific Sector of the Magellan Region (from Drake Passage to Chiloé Island), the Magellan Strait, the Beagle Channel, the Atlantic Sector of the Magellan Region (from Drake Passage to Península Valdés, and, from there, extending north up to ∼35ºS at a distance of 100–150 km from the coast, see Doti et al. 2014, Fig. 4), the Malvinas (Falkland) Islands, the Scotia Arc and the Antarctic Peninsula (including the South Shetland Islands).
Results
Species richness and abundance were lowest in the buffer sector for both isopods and cumaceans, highest in the core sector for isopods, and highest in the transition sector for cumaceans (Table 2).
The Jaccard similarity index (JSI) for the comparison between the core and buffer sectors was 0.50, whereas that for the comparison between the transition and core sectors and that for the comparison between the transition and buffer sectors were lower (0.35 and 0.33, respectively).
A total of 70 taxa (57 isopods and 13 cumaceans) distributed in 22 families (18 of isopods and 4 of cumaceans) were identified (Table 2, Online Resource 1). Of the 70 taxa identified, 50 correspond to nominal species, eight are most probably new species to science (designated by code letters), two are species provisionally identified (with the abbreviation “cf.” preceding the specific epithet), and 10 were identified to genus or family level. As many as 35 (70%) nominal species are recorded from the MPAN-BB for the first time (Table 2).
At the family level, for Paramunnidae, Janiridae, Diastylidae, Munnidae, Leuconidae, Joeropsididae and Nanastacidae three or more species were recorded. These seven families were found in the three sectors and all together accounted for 88.4% of the total specimens collected. Antarcturidae was the exception because although four species were recorded, this family was found only in two sectors and was represented by a few specimens. In contrast, Santiidae, Lampropidae, Stenetriidae, Anthuroidea, Chaetiliidae, Aegidae and Pseudidotheidae were represented by one species each, and all together accounted for only 0.15% of the total specimens collected. The remaining families were represented by one or two species with a moderate number of specimens (Fig. 2).

Isopods and cumaceans collected in the core, buffer and transition sectors of the MPAN-BB. Families are sorted in decreasing order of number of species, and then of abundance. On the right of each bar, the number of species and abundance (between parentheses) are shown
The 35 new records herein reported, plus the previous records from the literature, make a total of 55 nominal species currently known from Burdwood Bank (Table 3). None of these species is endemic to Burdwood Bank. Moreover, all of them, except Abyssianira dentifrons, were recorded from the Magellan Region (Table 3). Furthermore, the eight most probably new species (herein designated as “n. sp. A, B or C”) were also found in the Atlantic Sector of the Magellan Region (Table 2).
According to their depth distribution, the species from Burdwood Bank are divided in three major groups: 13 are known only from shallow waters (< 200 m), 32 reach intermediate depths (as deep as 1000 m), and 10 extend beyond 1000 m depth. The bathymetric range of 21 species is herein extended (Table 3).
Regarding the faunal similarities between Burdwood Bank and its neighboring areas, both multivariate analyses were congruent. The cluster analysis showed that Burdwood Bank and the Atlantic Sector of the Magellan Region (AS) presented the highest similarity (Fig. 3a), whereas the MDS analysis showed that Burdwood Bank was grouped more closely together with the AS (Fig. 3b). In particular, 42 out of the 55 nominal species from Burdwood Bank are also known from the AS. In contrast, only nine out of the 55 nominal species are known from the Antarctic Peninsula or the Scotia Arc (Table 3).

Similarity between Burdwood Bank and its neighboring areas based on the isopods and cumaceans listed in Table 3. a Hierarchical agglomerative clustering using group-average linking (dashed line, similarity threshold 50%). b Non-metric multidimensional scaling plot (MDS). AP Antarctic Peninsula (including South Shetland Is.), PS Pacific Sector of the Magellan Region, MS Magellan Strait, BC Beagle Channel, AS Atlantic Sector of the Magellan Region, BB Burdwood Bank, MI Malvinas/Falkland Is., SA Scotia Arc (includes South Georgia Is. and South Orkney Is., no records from Sandwich Is.)
Discussion
Biodiversity remarks
The large number of new records and new species of Isopoda and Cumacea herein reported can be ascribed to the fact that, for the samplings, we used an epibenthic sledge equipped with a net of 1-mm mesh size, which allowed us to collect a very large number of specimens (as many as 17,076 specimens in only three samples). It should be noted that the gears so far used in the MPAN-BB were not appropriate to collect small crustaceans. For instance, by using a bottom otter trawl, Schejter et al. (2016) reported only five species of isopods and no cumaceans.
Isopods have a remarkably high biodiversity in Burdwood Bank, with the following 13 species previously recorded from this area: Acanthoserolis schythei, Aega semicarinata, Ianthopsis laevis, Joeropsis curvicornis, J. intermedius, Munnogonium falklandicum, Neoserolis exigua, Paramunna integra, Pleurosigunum magnum, Natatolana pastorei, Serolis kempi, Iathrippa longicauda, and Cymodopsis cf. baegeli (Nordenstam 1933; Sheppard 1933; Kussakin 1967; De Broyer et al. 2003; Schejter et al. 2016; OBIS 2019).
Except for the last four, the remaining species listed above were also recorded in the current study (Table 2). Concerning the species not found in the current study, some clarifications are needed. Natatolana pastorei and Serolis kempi are distributed along the Patagonian coast (see Table 3) and although they were not present in any of the three samples herein analyzed, they have been found at other samples recently sorted from the MPAN-BB (unpublished data). Iathrippa longicauda was originally described from New Zealand, and afterwards reported from numerous subantarctic islands as well as from South America. In particular, Nordenstam (1933) recorded this species from the Malvinas (Falkland) Islands and from Burdwood Bank. Based on the broad geographic distribution and the variety of habitats reported for this species, Wilson and Wägele (1994) suggested that I. longicauda is a complex of species. It is noteworthy that none of the 155 specimens of Iathrippa herein examined belong to the I. longicauda complex.
The family Paramunnidae has a global distribution but its diversity in the temperate-cold waters of the southern hemisphere is remarkably high (Wilson 1980). In the current study, Paramunnidae was the most speciose and abundant family, which is in line with the results previously obtained for the Atlantic Sector of the Magellan Region (Doti et al. 2005, 2014).
Regarding Desmosomatidae, only one species, Eugerdella falklandica, is known from the Magellan Region (Malvinas/Falkland Islands, Nordenstam 1933). Among the material examined, 109 specimens were provisionally identified as Desmosoma sp. (Table 2). Among the Munnopsidae, Echinozone n. sp. A represents the first record of this genus for the South-West Atlantic. Regarding Stenetriidae and Antarcturidae, five and three species, respectively, have been reported from localities in the vicinity of Burdwood Bank (Schultz 1982; Kussakin and Vasina 1984, 1998). In the current study, only five specimens of Stenetriidae and 13 specimens of Antarcturidae were collected (Table 2). The Stenetriidae and Antarcturidae recorded from the South-West Atlantic were found from 400 to 4696 m depth. This may explain why, in our samples, these two families were represented by only a few specimens.
Very little information is available on the cumaceans from Burdwood Bank. Only eight species have been previously reported (Table 3). Six of them have been obtained from only two samples, viz., Cumella argentinae and Leucon assimilis were collected by the RV Vema 17, Sta. 59, 54º53.5′S, 60º26.5′W, 426–432 m, whereas Diastylis hammoniae, D. planifrons, Ekleptostylis heardi, and E. vemae were collected by the RV Eltanin Cr. 6, Sta. 344, 54º04′S, 58º46′W, 119 m (Petrescu 1994, 1995; McLelland and Meyer 1998; Roccatagliata and Mühlenhardt-Siegel 2000; Błazewicz-Paszkowycz and Heard 2005). More recently, Roccatagliata and Alberico (2016) recorded Holostylis uniramosa, based on specimens collected by the Argentine survey CAV 2013 at Sta. 4, 103 m (this is one of the stations herein studied) and Roccatagliata (2020) recorded Platytyphlops sarahae, based on specimens collected by the German survey ANTARKTIS XIX/5 (LAMPOS) at 271 m and by the Argentine survey BBB 2016 at 410–1000 m. The other five species found in the current contribution bring to 13 the total number of nominal species reported for Burdwood Bank (Table 3).
Concerning the family Leuconidae, the genera Eudorella and Leucon are herein reported from Burdwood Bank for the first time. Petrescu (1991) recorded E. gracilior Zimmer, 1907, E. sordida Zimmer, 1907, E. similis Calman, 1907 and E. fallax Zimmer, 1909 from the Magellan Region. Although the descriptions of these four species are incomplete, a fact that renders their identification problematic, none of them are conspecific with the two Eudorella species herein reported in Table 2. With regard to Leucon assimilis, this species has a wide range of distribution in the Southern Ocean and may indeed be a complex of cryptic species (see Mühlenhardt-Siegel 2011).
The Bodotriidae is a family of temperate latitudes, “negatively amphipolar” after Zimmer (1941) (see also Day 1975, 1978). Only one species, Leptocuma patagonicum Roccatagliata, 1993, is currently known from the Magellan Region and so far no species of this family have been recorded from Burdwood Bank.
Biogeography, endemism, and mechanisms of species dispersion
The isopods and cumaceans from Burdwood Bank have shown to be typically Magellanic (Fig. 3). In fact, all the species (except Abyssianira dentifrons) reported from Burdwood Bank are also present in the Magellan Region (see Table 3). It is worth noticing that high faunal affinities between Burdwood Bank and the Magellan Region have also been reported for other taxa, such as mollusks, bryozoans, polychaetes, decapods, and amphipods (Linse et al. 2003; Romero et al. 2003; Lovrich et al. 2005; Moyano 2005; De Broyer and Jażdżewska 2014; Bremec et al. 2019). These results suggest that the West and North channels that separate Burdwood Bank from the Argentine Continental Shelf do not form an effective barrier to species dispersion and gene flow.
No endemic isopods or cumaceans are known from Burdwood Bank (Table 3). Abyssianira dentifrons has not been reported from the Magellan Region but is known from the Puerto Rico Trench and the Argentine Basin (Doti and Roccatagliata 2006, and references therein). In addition, all the species most probably new to science herein reported had also been found in samples recently collected by the RV Puerto Deseado in other localities from the Argentine Continental Shelf (unpublished data).
Most of the Argentine Continental Shelf, which is one of the most extensive submarine platforms in the world, has been repeatedly uncovered during glacial times and flooded again during glacial terminations. During the Last Glacial Maximum (ca. 24,000 years BP), the sea level became established between −120 and −140 m below the present sea level and Burdwood Bank emerged. This paleo-island would have also emerged during older glacial cycles as well, and may have even been connected to the continent in the distant past (see Ponce et al. 2011; and references therein).
Few studies have described the biogeographic responses of marine species to the Last Glacial Maximum in the Magellan Region (see González-Wevar 2016; and citations therein). Leese et al. (2008) studied Serolis paradoxa (Fabricius 1775), a shallow-water isopod widely distributed in the Magellan Region. According to these authors, there is currently no effective gene flow for S. paradoxa between the Magellan Strait and the Malvinas (Falkland) Islands, and it has been absent for time exceeding the Last Glacial Maximum. Even more, they concluded that the specimens from these two localities very likely represent two cryptic species. Leese et al. (op. cit.) study deals with the connectivity between Patagonia and Malvinas (Falkland) Islands. Investigations over the effects of the glacial Quaternary cycles on the connectivity between Patagonia and Burdwood Bank are required.
Floating seaweed rafts, which are very abundant in the Magellan Region, could play an important role in maintaining the genetic connectivity between the populations from Patagonia and Burdwood Bank. Crustaceans are commonly reported from macrophyte rafts and peracarids, in particular, often dominate the rafting communities (see Rothäusler et al. 2012; Thiel and Fraser 2016). Helmut et al. (1994a, b) underlined the importance of the West Wind Drift (= ACC) in transporting floating macroalgae and their associated fauna, from the vicinity of Cape Horn towards South Georgia Island. Also, Nikula et al. (2010) reported that the macroalgal rafting is an important long-distance dispersal mechanism for intertidal kelp epifauna in the Subantarctic Region.
Among peracarids, several species of isopods and amphipods (but no cumaceans) have been reported on macrophyte rafts in the Magellan Region (Thiel and Gutow 2005; Hinojosa et al. 2007; Wichmann et al. 2012). Furthermore, Leese et al. (2010) studied the isopod Septemserolis septemcarinata (Miers 1881) from three remote and isolated Antarctic/Subantarctic islands (South Georgia, Bouvet, and Marion Islands), and concluded that, after the initial colonization of these islands, long-distance dispersal events occurred repeatedly in the life history of this species and maybe continue to occur today. As this is a strictly benthic species and lacks (like all peracarids) pelagic larvae, Leese et al. (op. cit.) suggested that this species disperses from west to east by passive rafting on floating substrata in the ACC.
Since Burdwood Bank is a submarine plateau with a minimum depth of 50 m, its environmental conditions are quite different from those of the coastal habitats, and not necessarily favorable for the settlement, expansion and persistence of the species coming via rafting. Nevertheless, it should be noted that four asellote species, i.e., Inathopsis laevis, Neojaera antarctica, Meridiosignum menziesi (= Paramunna menziesi) and Uromunna nana, recorded on rafting substrata from southern Chile by Wichmann et al. (2012) were also recorded in the benthic community from Burdwood Bank (Table 3). In contrast, Sphaeromatidae and the valviferans Idoteidae, which are common on macrophyte rafts from southern Chile, seem to be very scarce or even absent on the Burdwood Bank benthos (Thiel and Gutow 2005; Wichmann et al. 2012; and Table 3). To the best of our knowledge, there are no studies on kelp rafting in the MPAN-BB. This matter needs to be further investigated.
Finally, very little is known about the isopods and cumaceans that inhabit deeper waters around Burdwood Bank. In the current study, we report four isopod species belonging to the deep-sea families Desmosomatidae, Munnopsidae and Stenetriidae (only Tenopedunculus) (Table 2). These four species (414 specimens in total) were found exclusively in the transition sector (202 m depth). Schejter et al. (2016) analyzed the faunal composition of three stations from Burdwood Bank (core: 101 m, buffer: 113 m, shelf break: 232 m) and found that the core and buffer stations were more related to each other than to the shelf break station. Our results also show that the species composition of the transition station (the deepest one) differs from those of the core and buffer stations, which are more alike. Moreover, Fraysse et al. (2018) studied the sea stars (Asteroidea) from the southern tip of South America and stated that the shelf break stations (203–785 m depth) from Burdwood Bank form a cluster on its own. More recently, Teso et al. (2019) found that the chitons and gastropods from Burdwood Bank (80–200 m depth) and two localities on the Argentine Continental Shelf clustered together, whereas those of the slope of Burdwood Bank (200–1000 m depth) grouped with those of the Mar del Plata submarine canyon. Based on these previous results and the fact that the transition station of the MPAN-BB included some deep-sea isopods and cumaceans, it seems likely that the recently established MPAN-BB II (depth > 200 m) contains a distinct assemblage of species.
Only 16.4% of the nominal species reported from Burdwood Bank are also known from the Antarctic Peninsula or Scotia Arc (Table 3). This result agrees with the little correspondence previously observed at species level between the Magellanic and the Antarctic faunas, both for isopods and cumaceans (see Winkler 1994; Mühlenhardt-Siegel 1999; Castelló 2004; Choudhury and Brandt 2009).
Conclusions
The current research provides additional evidence to support the protection of the MPAN-BB. For the two taxonomic groups studied (isopods and cumaceans), we found not only a remarkably high diversity in the MPAN-BB but also a close affinity at species level between the MPAN-BB and the Magellan Region, particularly with the Argentine Sector of this region. These results prove that although the MPAN-BB is separated from the Argentine Continental Shelf by the West Channel (400 m depth) and the Malvinas Channel (400 to 3000 m depth), it definitely forms part of the Magellan Region.
The MPAN-BB II was recently created to protect the large biomass of vulnerable and fragile species (mainly corals and sponges) living on the southern slope of the bank. Very little is known about the peracarids from the slopes of the bank; however, its faunal composition is expected to differ from that reported from shallower waters (< 200 m depth). An indication of this change in the faunal composition is already present in the sample taken at the transition sector (202 m depth), which includes some deep-sea species that were absent in the core and buffer sectors (103 and 117 m depths, respectively).
This is the first comprehensive contribution on the isopods and cumaceans from the MPAN-BB. However, further studies on the peracarids from the MPAN-BB and its southern slope are necessary. These studies, together with those published and in progress on other taxonomic groups, are essential for the development of adequate conservation and management plans in the two marine protected areas mentioned above.
References
Alberico NA, Roccatagliata D (2008) Diastylis fabrizioi, a new species and a brief redescription of D. planifrons Calman, 1912 (Crustacea: Cumacea: Diastylidae) from South America. J Nat Hist 42:1039–1063
Alberico NA, Roccatagliata D (2011) On two species of Diastylis (Cumacea) from the southwestern Atlantic: D. sexpectinata n. sp. and D. hammoniae Zimmer, 1902. J Crust Biol 31:192–208
Bastida R, Torti MR (1973) Los isópodos Serolidae de la Argentina. Clave para su reconocimiento. Physis Sec A 32:19–46
Błazewicz-Paszkowycz M, Heard RW (2005) Observations on Cumacea (Malacostraca: Peracarida) from Antarctic and subantarctic waters. II. The rediscovery and redescriptions of Diastylis hammoniae Zimmer, 1902 and Diastylis planifrons Calman, 1912. Proc Biol Soc Wash 118:649–659
Bozzano G, Martín J, Spoltore DV, Violante RA (2017) Los Cañones Submarinos del Margen Continental Argentino: una síntesis sobre sus génesis y dinámica sedimentaria. Lat Am J Sedimentol Basin Anal 24:85–101
Brandt A (1998) Sphaeromatidae (Crustacea, Isopoda) of the Beagle Channel and description of Cymodopsis beageli n. sp. Beaufortia 48:137–162
Brandt A, Mühlenhardt-Siegel U, Schmidt A (1999) Density, diversity, and community patterns of selected peracarid taxa (Malacostraca) in the Beagle Channel, South America. In: Schram FR, von Vaupel Klein JC (eds) Crustaceans and the biodiversity crisis: Proceedings of the Fourth International Crustacean Congress. Koninklijke Brill NV, Leiden, pp 541–558
Bremec C, Elías R, Calla S, Genzano G, Puente Tapia A, Schejter L (2019) Polychaetes from Burdwood bank: “Namuncurá I” Marine Protected Area and slope, SW Atlantic Ocean. Rev Biol Trop 67:119–135
Bruce NL (2009) The marine fauna of New Zealand: Isopoda, Aegidae (Crustacea). NIWA Biodiv Mem 122:1–252
Castelló J (2004) Isopods (Crustacea, Isopoda) from the Spanish “Bentart-94/95” expeditions to the South Shetland Islands (sub-Antarctic). Polar Biol 28:1–14
Cavallotto JL, Violante RA, Hernández-Molina FJ (2011) Geological aspects and evolution of the Patagonian Continental Margin. Biol J Linn Soc 103:346–362
Choudhury M, Brandt A (2009) Benthic isopods (Crustacea, Malacostraca) from the Ross Sea, Antarctica: species checklist and their zoogeography in the Southern Ocean. Polar Biol 32:599–610
Clarke KR, Warwick RM (2001) Change in marine communities: an approach to statistical analysis and interpretation, 2nd edn. PRIMER-E, Plymouth
Corbera J (2000) Systematics and distribution of cumaceans collected during BENTART-95 cruise around South Shetland Islands (Antarctica). Sci Mar 64:9–28
Corbera J, San Vicente C, Sorbe JC (2009) Cumaceans (Crustacea) from the Bellingshausen Sea and off the western Antarctic Peninsula: a deep-water link with fauna of the surrounding oceans. Polar Biol 32:611–622
Day J (1975) South African Cumacea. Part 1. Family Bodotriidae, subfamily Vaunthompsoniinae. Ann S Afr Mus 66:177–220
Day J (1978) Southern African Cumacea. Part 2. Family Bodotriidae, subfamily Bodotriinae. Ann S Afr Mus 75:159–290
De Broyer C, Jażdżewska A (2014) Chapter 5.17. Biogeographic patterns of Southern Ocean benthic amphipods. In: De Broyer C, Koubbi P, Griffiths HJ, Raymond B, Udekemd’Acoz CD, et al. (eds) Biogeographic Atlas of the Southern Ocean. Scientific Committee on Antarctic Research, Cambridge, pp 155–165
De Broyer C, Lörz A-N, Nyssen F, Rauschert M, Cariceo Y, Ríos C (2003) Biodiversity, biogeography, phylogeny and trophodynamics of amphipod and isopod crustaceans. In: Arntz W, Brey T (eds) Expedition ANTARKTIS XIX/5(LAMPOS) of RV ‘‘Polarstern’’ in 2002. Ber Polarforsch Meeresforsch 462, pp 34–44
Di Luca J, Zelaya DG (2019) Gastropods from the Burdwood Bank (southwestern Atlantic): an overview of species diversity. Zootaxa 4544:41–78
Doti BL (2017) Three new paramunnids (Isopoda: Asellota: Paramunnidae) from the Argentine Sea, South-west Atlantic. J Mar Biol Assoc UK 97:1695–1709
Doti BL, Roccatagliata D (2006) On the Atlantic species of the genus Abyssianira Menzies, 1956 (Isopoda: Asellota: Paramunnidae). Zootaxa 1252:1–35
Doti BL, Roccatagliata D (2013) Two new species of the genus Munnogonium (Isopoda: Asellota: Paramunnidae) from Argentina. Zootaxa 3717:301–319
Doti BL, Roccatagliata D, López Gappa J (2014) An inverse latitudinal biodiversity pattern in asellote isopods (Crustacea, Peracarida) from the Southwest Atlantic between 35º and 56ºS. Mar Biodiv 44:115–125
Doti BL, Roccatagliata D, Scarabino F (2008) Range extension of Pseudidothea miersii (Studer, 1884) (Isopoda, Valvifera) and new evidence of its likely commensal relationship with a sea urchin. Crustaceana 81:883–888
Doti BL, Roccatagliata D, Zelaya DG (2005) The shallow-water Asellota (Crustacea: Isopoda) from the Beagle Channel: preliminary taxonomic and zoogeographical results. Sci Mar 69:159–166
Falabella V (2017) Área Marina Protegida Namuncurá—Banco Burdwood. Contribuciones para la línea de base y el plan de manejo. Jefatura de Gabinete de Ministros, Buenos Aires
Fraysse C, Calcagno J, Pérez AF (2018) Asteroidea of the southern tip of South America, including Namuncurá Marine Protected Area at Burdwood Bank and Tierra del Fuego Province, Argentina. Polar Biol 41:2423–2433
Fraysse CP, Pérez AF, Calcagno JA, Boy CC (2020) Energetics and development modes of Asteroidea (Echinodermata) from the Southwestern Atlantic Ocean including Burdwood Bank/MPA Namuncurá. Polar Biol 43:175–186
García Alonso VA, Brown D, Martín J, Pájaro M, Capitanio FL (2018) Seasonal patterns of Patagonian sprat Sprattus fuegensis early life stages in an open sea Sub-Antarctic Marine Protected Area. Polar Biol 41:2167–2179
Gerken S, Haye PA (2018) Hemilamprops chilensis sp. nov. (Crustacea: Cumacea: Lampropidae) from the coast of Chile, with a key to the Chilean Lampropidae and remarks on the status of H. ultimaespei Zimmer, 1921 and H. lotusae Băcescu, 1969. Zootaxa 4399:351–360
González-Wevar CA, Rosenfeld S, Segovia NI, Hüne M, Gérard K, Ojeda J, Mansilla A, Brickle P, Díaz A, Poulin E (2016) Genetics, Gene Flow, and Glaciation: The Case of the South American Limpet Nacella mytilina. PLoS ONE 11:e0161963. https://doi.org/10.1371/journal.pone.0161963
Gordillo S, Bayer MS, de Aranzamendi MC, Taverna A, Morán GA (2019) The ecological role of brachiopods in the Namuncurá MPA/Burdwood Bank, off southern South America. Mar Biodiv 49:1667–1681
Guerrero RA, Baldoni A, Benavides H (1999) Oceanographic conditions at the southern end of the Argentine continental slope. INIDEP Doc Cient 5:7–22
Güller M, Zelaya DG (2017) New insights into the diversity of rissoids from sub-antarctic and antarctic waters (Gastropoda: Rissooidea). Polar Biol 40:1923–1937
Harris PT, Whiteway T (2011) Global distribution of large submarine canyons: geomorphic differences between active and passive continental margins. Mar Geol 285:69–86
Helmut B, Veit RR, Holberton R (1994a) Dispersal of benthic invertebrates in the Scotia Arc by kelp rafting. Antarct J US 29:145–147
Helmut B, Veit RR, Holberton R (1994b) Long-distance dispersal of a subantarctic brooding bivalve (Gaimardia trapesina) by kelp-rafting. Mar Biol 120:421–426
Hinojosa I, González E, Ugalde P, Valdivia N, Macaya E, Thiel M (2007) Distribución y abundancia de macroalgas flotando a la deriva y su fauna peracarida asociada en los canales de la XI Región, Chile. Cienc Tecnol Mar 30:37–50
Jones NS (1984) The family Nannastacidae (Crustacea: Cumacea) from the deep Atlantic. Bull Br Mus (Nat Hist). Zool Ser 46:207–289
Keable SJ (2006) Taxonomic revision of Natatolana (Crustacea: isopoda: Cirolanidae). Rec Aust Mus 58:133–244
Kensley B (1975) Five species of Jaeropsis from the southern Indian Ocean (Crustacea, Isopoda, Asellota). Ann S Afr Mus 67:367–380
Kussakin OG (1967) Fauna of Isopoda and Tanaidacea in the coastal zones of the Antarctic and Subantarctic waters. Biol Rep Soviet Antarct Exped 1955–1958(3):220–389
Kussakin OG, Vasina GS (1984) Deep-sea lower asellotes from the Scotia Sea and South Sandwich Trench. Biol Morya (Vladivostok) 6:9–17 (in Russian)
Kussakin OG, Vasina GS (1998) New bathyal and abyssal arcturids from the western Antarctic and Subantarctic (Crustacea: Isopoda: Arcturidae). Zoosyst Ross 7:55–75
Ledoyer M (1993) Cumacea (Crustacea) de la campagne EPOS 3 du R.V. Polarstern en mer de Weddell. Antarctique J Nat Hist 27:1041–1096
Leese F, Agrawal S, Held C (2010) Long-distance island hopping without dispersal stages: transportation across major zoogeographic barriers in a Southern Ocean isopod. Naturwissenschaften 97:583–594
Leese F, Kop A, Wägele JW, Held C (2008) Cryptic speciation in a benthic isopod from Patagonian and Falkland Island waters and the impact of glaciations on its population structure. Front Zool 5:19
Linse K, Schrödl M, Zelaya D (2003) Biodiversity, Biogeography and Evolution of Mollusca. In: Arntz W, Brey T (eds) Expedition ANTARKTIS XIX/5(LAMPOS) of RV ‘‘Polarstern’’ in 2002. Ber Polarforsch Meeresforsch 462, pp 19–28
López Gappa J, Liuzzi MG, Zelaya DG (2018) A new genus and species of cheilostome bryozoan associated with hermit crabs in the subantarctic Southwest Atlantic. Polar Biol 41:733–741
Lorenti M, Mariani S (1997) Isopod assemblages in the Straits of Magellan: structural and functional aspects. Polar Biol 18:254–259
Lörz AN, di Renzo A, Nickel J (1999) Comparative analysis of three sampling gear types for marine macrobenthos. Ber Polarforsch 330:134–151
Lovrich GA, Romero MC, Tapella F, Thatje S (2005) Distribution, reproductive and energetic conditions of decapod crustaceans along the Scotia Arc (Southern Ocean). Sci Mar 69:183–193
Matano RP, Palma ED, Combes V (2019) The Burdwood Bank circulation. JGR. Oceans 124:6904–6926
McLelland JA, Meyer GH (1998) Ekleptostylis heardi (Diastylidae), a new cumacean species from South Atlantic waters. Proc Biol Soc Wash 111:278–287
Moretti AN, Roccatagliata D (2007) On two allied species of the genus Diastylis from southern South America: D. granulata Zimmer, 1921 and D. argentata Calman, 1912 (Crustacea: Cumacea: Diastylidae). J Nat Hist 41:2621–2639
Moyano HI (2005) Scotia Arc bryozoan from the LAMPOS expedition: a narrow bridge between two different faunas. Sci Mar 69:103–112
Mühlenhardt-Siegel U (1999) On the biogeography of Cumacea (Crustacea, Malacostraca). A comparison between South America, the Subantarctic Islands and Antarctica: present state of the art. Sci Mar 63:295–302
Mühlenhardt-Siegel U (2003) Redescription of Hemilamprops ultimaespei Zimmer, 1921 (Crustacea: Cumacea: Lampropidae) from the Beagle Channel, South America. Mitt Hamb Zool Mus Inst 100:119–126
Mühlenhardt-Siegel U (2011) New and known species of the family Leuconidae (Cumacea, Peracarida) from Antarctic deep-sea basins. Zootaxa 3117:1–68
Nikula R, Fraser CI, Spencer HG, Waters JM (2010) Circumpolar dispersal by rafting in two subantarctic kelp-dwelling crustaceans. Mar Ecol Prog Ser 405:221–230
Nordenstam A (1933) Marine Isopoda of the families Serolidae, Idotheidae, Pseudidotheidae, Arcturidae, Parasellidae and Stenetriidae mainly from South Atlantic. Further Zool Res Swed Antarct Exped 1901–1903(3):1–284
OBIS (2019) Ocean Biogeographic Information System. Intergovernmental Oceanographic Commission of UNESCO. Available at http://www.iobis.org. Accessed 2019
Pandey A, Parson L, Milton A (2010) Geochemistry of the Davis and Aurora Banks: possible implications on evolution of the North Scotia Ridge. Mar Geol 268:106–114
Park JY, Wägele JW (1995) On a small collection of Valvifera (Crustacea Isopoda) from the Magellan Strait, with description of Platidotea magellanica n.gen. n.sp. Bull Zoöl Mus Univ Amst 14:57–99
Penchaszadeh PE, Pastorino G, Martinez MI, Miloslavich P (2019) Spawn and development of the gastropod Americominella longisetosa (Castellanos and Fernández, 1972) (Mollusca: Buccinidae) from the Southwestern Atlantic deep sea. Deep Sea Res I 143:43–49
Penchaszadeh PE, Teso V, Pastorino G (2017) Spawn in two deep-sea volute gastropods (Neogastropoda: Volutidae) from southwestern Atlantic waters. Deep Sea Res I 130:55–62
Pérez-Barros P, Albano M, Diez MJ, Lovrich GA (2020) Pole to pole: the deep-sea king crab Lithodes couesi (Decapoda: Lithodidae) in the Burdwood Bank, Southwestern Atlantic Ocean. Polar Biol 43:81–86
Petrescu I (1991) Contribution to the knowledge of genus Eudorella Norman, 1867 (Crustacea, Cumacea, Leuconidae) with description of two new species: Eudorella bacescui n.sp. and Eudorella menziesi n.sp. Trav Mus Hist Natl Gr Antipa 31:375–385
Petrescu I (1994) Contributions to the knowledge of Leucon genus (Crustacea: Cumacea) from the waters of South America. Trav Mus Hist Natl Gr Antipa 34:325–345
Petrescu I (1995) Cumaceans (Crustacea: Peracarida) from the South American coasts collected by the R/V “Vema”. Trav Mus Hist Natl Gr Antipa 35:49–86
Petrescu I, Heard RW (2000) The status of the genus Campylaspis Sars, 1865 (Crustacea: Cumacea) from the Antarctic Ocean. Trav Mus Hist Natl Gr Antipa 42:75–97
Piola AR, Gordon AL (1989) Intermediate waters in the southwest South Atlantic. Deep Sea Res Part A 36:1–16
Ponce JF, Rabassa J, Coronato A, Borromei AM (2011) Palaeogeographical evolution of the Atlantic coast of Pampa and Patagonia from the last glacial maximum to the Middle Holocene. Biol J Linn Soc 103:363–379
Rehm P, Thatje S, Mühlenhardt-Siegel U, Brandt A (2007) Composition and distribution of the peracarid crustacean fauna along a latitudinal transect off Victoria Land (Ross Sea, Antarctica) with special emphasis on the Cumacea. Polar Biol 30:871–881
Ríos C, Mutschke E, Morrison E (2003) Biodiversidad bentónica sublitoral en el estrecho de Magallanes, Chile. Rev Biol Mar Oceanogr 38:1–12
Roccatagliata D (2020) On the deep-sea lampropid Platytyphlops sarahae n. sp. from Argentina, with remarks on some morphological characters of Cumacea. Zool Anz 286:135–145
Roccatagliata D, Alberico NA (2016) Two new cumaceans (Crustacea: Peracarida) from the South-West Atlantic with remarks on the problematic genus Holostylis Stebbing, 1912. Mar Biodiv 46:163–181
Roccatagliata D, Mühlenhardt-Siegel U (2000) Remarks on the taxonomy of the genus Ekleptostylis Stebbing, 1912 (Crustacea: Cumacea: Diastylidae). Proc Biol Soc Wash 113:696–709
Romero MC, Tapella F, Lovrich GA, Thatje S (2003) Decapod Reproduction and Distribution. In: Arntz W, Brey T (eds) Expedition ANTARKTIS XIX/5(LAMPOS) of RV ‘‘Polarstern’’ in 2002. Ber Polarforsch Meeresforsch 462, pp 67–72
Rothäusler E, Gutow L, Thiel M (2012) Floating Seaweeds and Their Communities. In: Wiencke C, Bischof K (eds) Seaweed biology. Ecological studies (analysis and synthesis), vol 219. Springer, Berlin, pp 359–380
Schejter L, Bertolino M, Calcinai B (2017a) Description of Antho (Plocamia) bremecae sp. nov. and checklist of Microcionidae (Demospongiae: Poecilosclerida) from Burdwood Bank and neighboring areas, SW Atlantic Ocean. Zootaxa 4312:580–594
Schejter L, Genzano G, Gaitán E, Perez CD, Bremec CS (2020) Benthic communities in the Southwest Atlantic Ocean: Conservation value of animal forests at the Burdwood Bank slope. Aquatic Conserv: Mar Freshw Ecosyst 30:426–439
Schejter L, Martin J, Lovrich G (2017b) Unveiling the submarine landscape of the Namuncurá Marine Protected Area, Burdwood Bank, SW Atlantic Ocean. Pan-Am J Aquat Sci 12:248–253
Schejter L, Rimondino C, Chiesa I, Díaz de Astarloa JM, Doti B, Elías R, Escolar M, Genzano G, López-Gappa J, Tatián M, Zelaya DG, Cristobo J, Perez CD, Cordeiro RT, Bremec CS (2016) Namuncurá Marine Protected Area: an oceanic hot spot of benthic biodiversity at Burdwood Bank, Argentina. Polar Biol 39:2373–2386
Schultz GA (1982) Species of Protallocoxoidea and Stenetrioidea (Isopoda, Asellota) from the Antarctic and southern seas. Antarct Res Ser 10:17–20
Sivertsen E, Holthuis LB (1980) The marine isopod Crustacea of the Tristan da Cunha Archipelago. Gunneria 35:1–128
Sheppard EM (1933) Isopod Crustacea, Part I. The family Serolidae. Discover Rep 7:253–362
Sheppard EM (1957) Isopod Crustacea, Part II. The sub-order Valvifera. Families: Idoteidae, Pseudidotheidae and Xenarcturidae fam. n. with a supplement to Isopod Crustacea, Part 1. The family Serolidae. Discover Rep 29:141–198
Song JH, Bruce NL, Min GS (2018) The first records of Stenobermuda Schultz, 1982 and Tenupedunculus Schultz, 1979 from Australia, with description of two new species from the Great Barrier Reef (Isopoda, Asellota, Stenetriidae). ZooKeys 733:1–24
Taverna A, Lagger C, Maggioni T, Reyna P, Lovrich G, Tatián M (2018) Ascidian distribution provides new insights to help define the biogeographic provinces in the South American Region. Polar Biol 41:1123–1131
Teso V, Penchaszadeh PE (2019) Development of the gastropod Trochita pileus (Calyptraeidae) in the sub-Antarctic Southwestern Atlantic. Polar Biol 42:171–178
Teso V, Urteaga D, Pastorino G (2019) Assemblages of certain benthic molluscs along the southwestern Atlantic: from subtidal to deep sea. BMC Ecol 19:49
Thiel M, Fraser C (2016) The role of floating plants in dispersal of biota across habitats and ecosystems. In: Ólafsson E (ed) Marine macrophytes as foundation species. CRC Press, Boca Raton, pp 76–99
Thiel M, Gutow L (2005) The ecology of rafting in the marine environment. II. The rafting organisms and community. Oceanogr Mar Biol 43:279–418
Torres Carbonell PJ, Dimieri LV, Olivero EB, Bohoyo F, Galindo-Zaldívar J (2014) Structure and tectonic evolution of the Fuegian Andes (southernmost South America) in the framework of the Scotia Arc development. Global Planet Change 123:174–188
Urcola MR, Zelaya DG (2018) The genus Cyamiocardium Soot-Ryen, 1951 (Bivalvia: Cyamiidae) in sub-Antarctic and Antarctic waters. Polar Biol 41:1157–1174
Wägele JW (1994) Notes on Antarctic and South American Serolidae (Crustacea, Isopoda) with remarks on the phylogenetic biogeography and a description of new genera. Zool Jahrb Abt Syst Oekol Geogr Tiere 121:3–69
Wichmann C, Hinojosa IA, Thiel M (2012) Floating kelps in Patagonian Fjords: an important vehicle for rafting invertebrates and its relevance for biogeography. Mar Biol 159:2035–2049
Wilson GD (1980) New insights into the colonization of the deep sea: Systematics and zoogeography of the Munnidae and the Pleurogoniidae comb. nov. (Isopoda; Janiroidea). J Nat Hist 14:215–236
Wilson GDF, Wägele J-W (1994) Review of the family Janiridae (Crustacea: Isopoda: Asellota). Invertebr Taxon 8:683–747
Wilton DW, Pirie JHH, Brown RNR (1908) Zoological log. Rep Sci Results Scot Natl Antarct Exped 4 (Zool), Part l:1–105
Winkler H (1994) Characterization of the Scotia Arc isopod fauna (Crustacea, Malacostraca) from a biogeographical point of view: a multivariate approach. Ber Polarforsch Meeresforsch 139:1–196 (in German)
Zimmer C (1941) Cumacea. Bronn’s Klassen des Tierreichs.V. I. Crustacea, 4th edn. Buch. Becker & Erler Akademische Verlagsgesellschaft, Leipzig, pp 1–222
Acknowledgements
We thank the officers and crew of the RV Puerto Deseado for their assistance during the CAV 2013 survey. We are also grateful to Alejandro Martínez (IBBEA, CONICET-UBA) for technical assistance at the laboratory, and Gustavo Lovrich (CADIC, CONICET), Juan López Gappa (MACN, CONICET) and Emanuel Pereira (IBEEA, CONICET-UBA) for their suggestions and help. We also thank Saskia Brix (DZMB, Germany), Martin Thiel (UCN, Chile) and an anonymous reviewer for their valuable comments, which helped to improve the quality of the manuscript. Finally, we thank María Victoria González Eusevi for revising the English. This study was supported by grants from the University of Buenos Aires (UBACyT 2018-2020 20020170100328BA) and CONICET (PIP 2014-2016 11220130100434CO), Argentina.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
All applicable international, national and/or institutional guidelines for the care and use of animals were followed.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Doti, B.L., Chiesa, I.L. & Roccatagliata, D. Biodiversity of Isopoda and Cumacea (Peracarida, Crustacea) from the Marine Protected Area Namuncurá-Burdwood Bank, South-West Atlantic. Polar Biol 43, 1519–1534 (2020). https://doi.org/10.1007/s00300-020-02725-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-020-02725-z