
Abstract
Key message
Mutation of the AM1 gene causes an albino midrib phenotype and enhances tolerance to drought in rice
Abstract
K+ efflux antiporter (KEA) genes encode putative potassium efflux antiporters that are mainly located in plastid-containing organisms, ranging from lower green algae to higher flowering plants. However, little genetic evidence has been provided on the functions of KEA in chloroplast development. In this study, we isolated a rice mutant, albino midrib 1 (am1), with green- and white-variegation in the first few leaves, and albino midrib phenotype in older tissues. We found that AM1 encoded a putative KEA in chloroplast. AM1 was highly expressed in leaves, while lowly in roots. Chloroplast gene expression and proteins accumulation were affected during chlorophyll biosynthesis and photosynthesis in am1 mutants. Interestingly, AM1 was induced by salt and PEG, and am1 showed enhanced sensitivity to salinity in seed germination and increased tolerance to drought. Taken together, we concluded that KEAs were involved in chloroplast development and played important roles in drought tolerance.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rice is one of the most important crops in the world. Photosynthesis, as a resource for energy in plants, is closely related to crops yield. Chloroplasts, where photosynthesis occurs, play essential roles not only in photoautotrophic growth but also in transduction of stress-associated signals. Most of chloroplast proteins (>80 %) are encoded within the nucleus and imported into the chloroplast after translation in the cytosol (Rodermel 2001). In rice, nuclear genes modulating chloroplast development have been largely studied. Mutations of them often cause the abnormal coloration from the seedling stage (Archer and Bonnett 1987).
Chlorophyll-deficient mutants are ideal genetic materials to gain insight into molecular mechanisms that modulate chlorophyll biosynthesis and chloroplast development in plants. To date, a considerable number of such mutants have been discovered in rice. Based on phenotypes, they were referred to as albino, pale-green chlorine (yellow), stripe and zebra (Jung et al. 2003). Almost all of known gene loci involved in these mutations were located to the chromosomes, and many have been cloned and characterized (Zhou et al. 2013; Dong et al. 2013; Su et al. 2012; Kusumi et al. 2011; Yoo et al. 2009; Sugimoto et al. 2004, 2007; Zhang et al. 2006; Song et al. 2004; Jung et al. 2003). However, few have been reported to associate with abiotic stresses.
The K+ efflux antiporters (KEAs) comprised a new family originated from bacterial glutathione-regulated K+ efflux transporters KefB and KefC with an N-terminal Na_H exchanger domain and a C-terminal KTN-NAD (H)-binding domain (also called TrkA_N domain) (Chanroj et al. 2012; Choe 2002). The KEA family was categorized into three types: KEAIa, KEAIb and KEAII (Chanroj et al. 2012). KEAIa and KEAIb contain a C-terminal KTN domain which is lost in the KEAII. Clades Ia and Ib can be distinguished by its N-terminal domain. Clades Ia has a long hydrophilic N-terminal domain of about 570–770 amino acids, while clades Ib does not have and its C-terminus is slightly longer than the other sequence (Chanroj et al. 2012).The KEA genes in higher plants were firstly reported and classified in the Arabidopsis thaliana (Maser et al. 2001), where KEA genes could be divided into two main branches with AtKEA1-3 and AtKEA4-6. AtKEA1-3 harbored a complete C-terminal KTN domain and was closely related to bacterial EcKefB and EcKefC-like proteins (Booth 2003), and AtKEA4-6 had lost the KTN domain in the C-terminus and shared the high homology to cyanobacterial sequences, similar to the TMCO3-like proteins from metazoan (Chanroj et al. 2012).
To date, little information is available on the function of KEAs in plants. Only a member of KEA family from Arabidopsis has been functionally characterized. AtKEA2, a homolog of bacterial KefC, encodes a K+/H+ antiporter and the truncated AtKEA2 (AtsKEA2) lacking the N-terminal domain can well complement disruption of the Na+(K+)/H+ antiporter Nhx1p to confer hygromycin resistance and tolerance to Na+ or K+ stress and mediated cation/H+ exchange with preference for K+ = Cs+ > Li+ > Na+ in yeast (Aranda-Sicilia et al. 2012). Additionally, the AtKEA2 was predicted to have a chloroplast transit peptide which was confirmed by subcellular localization, suggesting its functions in chloroplast or plastids development (Aranda-Sicilia et al. 2012). However, no genetic evidence with any relative KEA mutants is reported to support this hypothesis.
In this paper, we isolated and characterized the first KEA mutant am1 with a chlorophyll-deficient phenotype in the leaves by map-based cloning in rice. Abnormal chloroplast gene expression and protein accumulation in am1 suggested the crucial function of KEA in chloroplast development. In addition, am1 enhanced drought tolerance as compared to wild-type. Our findings revealed that the KEAs played important roles in tolerance to abiotic stresses in rice.
Materials and methods
Plant materials and growth conditions
The rice albino midrib (am1) mutant was isolated from a 60Co-irradiated mutant pool of japonica rice cultivar Wu B. The am1 was crossed with Jin 23B and Pei’ai64 to construct the F2 population for mapping. Rice plants were grown in a paddy field in Beijing (39°54′N, summer season, temperate climate) and Hainan (18°16′N, winter season, subtropical climate) under local conditions or in a growth chamber. For the temperature treatments, seedlings were planted in a growth chamber under 12 h of light/12 h of dark at a constant 30 or 20 °C.
Chlorophyll and carotenoid content measurement
Chlorophyll and carotenoid contents were measured using a spectrophotometer according to the method described by (Arnon 1949). Briefly, leaves (approximately 0.2 g FW) were cut and marinated in 95 % ethanol for 24 h under dark conditions. The supernatants were collected by centrifugation and analyzed with a Spectrophotometer (Beckman Coulter DU-800, USA) at 665, 649 and 470 nm, respectively.
Transmission electron microscopy
Samples of wild-type and am1 mutant leaves were fixed in 2.5 % glutaraldehyde at 4 °C for 4 h, rinsed and incubated overnight in 1 % OsO4 at 4 °C, dehydrated using an ethanol series, and finally embedded in Spurr’s medium prior to thin sectioning. Samples were stained again with uranylacetate and examined with a Hitachi H-7650 transmission electron microscope.
Mapping and cloning of AM1
For mapping AM1, a cross was made between am1 mutant and an indica cultivar PA 64. Firstly, the F2 population of ten individuals with typical am1 mutant phenotype was used for linkage analysis, then 882 F2 plants with am1 mutant phenotypes for fine-mapping. Sequence polymorphisms between Nipponbare (japonica) and 93-11 (indica) were identified and used to develop molecular markers. Primer pairs were designed using the Web server program dCAPS Finder 2.0 (http://helix.wustl.edu/dcaps/dcaps.html). The PCR procedure was as follows: 95 °C for 5 min, followed by 34 cycles of 95 °C for 30 s, 58 °C for 30 s, 72 °C for 40 s, and a final elongation step at 72 °C for 5 min.
Complementation of the am1 mutant
For complementation of the AM1 mutation, a 6.4 kb DNA fragment containing the entire CDS of AM1 and a 2.8 kb upstream sequence was amplified by PCR from Wu B, respectively, and then cloned into the binary vector pCAMBIA1305. The constructs were introduced into am1 mutant by Agrobacterium tumefaciens-mediated transformation as described previously (Hiei et al. 1994).
Sequence and phylogenetic analysis
Gene prediction and structure analysis were performed using the GRAMENE database (http://www.gramene.org/). Homologous sequences of AM1 were identified using the Blastp search program of the National Center for Biotechnology Information (NCBI, http://www.ncbi.nlm.nih.gov/). Multiple sequence alignments were conducted with DNAMAN version 6.0(Lynnon Biosoft). The chloroplast transit peptide (CTP) was predicted by the ChloroP program. PCR primers were designed with the Primer premier 5.0 software. A phylogenetic tree was constructed using MEGA v4.1 software (http://www.megasoftware.net/).
RNA preparation and qRT-PCR analysis
Total rice RNA was extracted with an RNA prep pure kit (Zymo Research, Orange, CA) according to the manufacturer’s instructions. The first-strand cDNA was synthesized from 2 μg of total RNA using the SuperScript II kit (TaKaRa). Primer pairs (suppl Table S3) were designed using Primer Express (Applied Biosystems) and listed in. qRT-PCR was conducted using an ABI7500 fast system with the SYBR Premix Ex Taq (TaKaRa; RR041A). The procedure was as follows: initial polymerase activation for 30 s at 95 °C followed by 40 cycles of 95 °C for 5 s and 60 °C for 34 s. For each sample, qRT-PCR was performed with three technical replicates on three biological replicates. The 2\(^{{ - \vartriangle \vartriangle C_{\text{T}} }}\) method was used to analyze relative transcript levels in gene expression.
Subcellular localization of AM1-GFP proteins in rice protoplasts
The coding sequence of AM1 was amplified and cloned into the N-terminus of GFP under control of the CaMV 35S promoter in the transient expression vector pAN580-GFP, resulting in pAN580-AM1-GFP. The recombinant vector was then transformed into rice protoplasts according to protocols described previously (Chen et al. 2006). GFP fluorescence in the transformed protoplasts was visualized using a confocal laser-scanning microscope (LSM 700; Carl Zeiss, Germany).
Histochemical GUS assay and DAB staining
A putative 2.7 kb promoter genomic fragment upstream of the ATG start codon was amplified by PCR, and the PCR product was cloned into the binary vector pCAMBIA1305 to generate the transformation plasmid pproAM1::GUS. GUS histochemical assays were performed as described previously (Zhou et al. 2013). The stained tissues were photographed using a Canon camera (50D) and enhanced using Photoshop CS6 (Adobe).
H2O2 was detected by DAB staining as described previously (ThordalChristensen et al. 1997). The leaves from wild-type and am1 plants were detached and vacuum-infiltrated with the DAB solution (1 mg of DAB dissolved in 1 ml of distilled water, pH 3.8). The leaves were placed in a growth chamber for 24 h at 25 °C and cleared in boiling ethanol (96 %) for 10 min before photographing.
Protein extraction, SDS–PAGE and western blotting
Leaf material was homogenized in lysis buffer [25 mM Tris–HCl, pH 7.6, 0.15 M NaCl, and 2 % sodium dodecyl sulfate (SDS), 0.01 % 2-mercaptoethanol]. Sample amounts were standardized by fresh weight. The protein samples were separated by 10 % SDS–polyacrylamide gel electrophoresis (PAGE). After electrophoresis, the proteins were transferred onto a PVDF membrane (Millipore) and incubated with specific antibodies. The signals were detected using an ECL Plus Western Blotting Detection Kit (Thermo) and visualized by an imaging system (ChemiDoc™X-RS; Bio-Rad).
Results
Phenotypic characterization of the am1 mutant
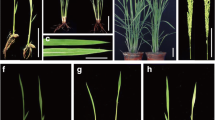
In the paddy filed, the am1 mutant displayed green- and white-variegation in the first few leaves, and the variegation appeared to be randomly distributed on the surface of the mutant leaves and predominantly near the leaf margins (Fig. 1a, b). Approximately from the early tillering stage, the newly-grown leaves showed slightly yellow-green phenotype but in the leaf vein especially in midrib displayed albino (Fig. 1c, d). Consistent with the observations, the contents of chlorophyll and carotenoid in leaves of am1 mutant were much lower than that of WT at the whole developmental stage (Fig. 1e, f). At maturity, the major agronomic traits such as plant height, number of tillers, and grain yield in am1 plants were indistinguishable from those of wild-type plants (Table S1). Additionally, the degree of chlorosis in am1 were strongly influenced by low temperature (such as 20 °C) (Fig. S1).

Phenotypic characteristics of wild-type (WT) and am1 mutant. a 2-week-old wild-type (left) and am1 (right) seedlings. b Enlarged views of the leaves from a. c 8-week-old wild-type (left) and am1 (right) seedlings. d Enlarged views of the leaves from c. e–f The pigment contents in leaves of 2- and 8-week-old am1 mutants are much lower than that in wild-type. Chla chlorophyll a, Chlb chlorophyll b, Chl total chlorophyll, Car carotenoid. Data represents mean ± SD from three independent biological replicates. Asterisks indicate statistically significant differences compared with wild-type (Student’s t test: *P < 0.05;**P < 0.01)
To investigate whether the mutation phenotype of the am1 is associated with ultrastructure changes in the chloroplasts, we compared the ultrastructure of chloroplasts between white sectors of am1 mutant leaves and wild-type normal leaves using transmission electron microscopy (TEM). As shown in Fig. 2, cells in WT leaves contained mature chloroplasts displaying well-organized lamellar structures and were equipped with normally stacked grana and thylakoid membranes. In contrast, cells of am1 mutant contained abnormal plastids which were heretoplastidic without any organized lamellar structures. These observations revealed that the AM1 mutation affected chloroplast development. Chloroplasts are the organelles in plant cells that perform photosynthesis. To test whether the photosynthetic apparatus was affected in am1 mutants, we compared some key parameters between wild-type and am1 plants using a pulse-amplitude-modulation (PAM) chlorophyll fluorometer. These parameters include photochemical efficiency of PSII (ΦPSII), nonphotochemical quenching (NPQ, qN), photochemical quenching (qP, qL) and the maximal efficiency of PSII (Fv/Fm). These values were almost comparable between am1 and wild-type plants (Table S2), indicating the PSII apparatus was not impaired in am1 mutant.

Observation of chloroplasts ultrastructure in mesophyll cells of 2-week-old wild-type and am1 plants. a, b TEM illustrates the normal chloroplast ultrastructure in wild-type leaves by scale-up. c, d the am1 mutant show the altered chloroplast ultrastructure in white tissues compared with wild-type. The black boxes indicate the enlarged sections. Cp chloroplast, Mt mitochondria, T thylakoid lamellar. Bars = 2 μm in a and c, 200 nm in b and d
Map-based cloning of AM1 gene
Genetic analysis from the crosses am1/Jin 32B and am1/Pei’ai64 indicated that the phenotype conferred by am1 was caused by a single, recessive nuclear mutation (Table 1). Using F2 population from the cross between am1 and Pei’ai64, the AM1 locus was mapped on the long arm of chromosome 4, between markers W1 and W11 (Fig. 3a, b). Eleven insertion-deletion polymorphic (InDel) markers were designed between W1 and W11 (Table S3). The AM1 was further narrowed down to an 81 kb region between markers W5 and W7 (Fig. 3c) containing 13 putative ORFs (Fig. 3d). We sequenced all ORFs and only found a 2-bp deletion in the tenth ORF (LOC_Os04g58620) at the position 226 and 228 bp away from the start codon ATG, causing a premature stop codon (Fig. 3d).We then introduced a 6.4-kb genomic fragment containing the transcriptional regulation elements and the full-length coding sequence of AM1 into the am1 mutant. As a result, all of 15 independent transgenic lines transformed with vector pAM1 containing the wild-type LOC_Os04g58620 gene completely rescued the mutant phenotype by comparing the leaf color of transgenic plants with wild-type plants and am1 mutant (Fig. 3e–g). We failed to obtain the RNAi transgenic plants. The T0 generation transgenic seedlings all showed an albinism and were seedling-lethal (data not shown). Nevertheless, these results confirmed that LOC_Os04g58620 corresponded to the AM1 gene.

Map-based cloning of the AM1 gene. a The AM1 locus was initially localized to the region between W1 and W11 markers in the chromosome 4 (Chr4). b Fine mapping of AM1 which was spanned by BAC clones OSJNBb0017I01, OSJNBa0032F06 and OSJNBa0088H09. c The AM1 locus was narrowed to a 81 kb region between the markers W5 and W7. The number of the recombinants identified from an F2 population of 882 recessive phenotypes is shown below each marker. d The region contained 13 open reading frames (ORFs) and a 2 bp deletion (indicated by red arrow) in the ORF10, resulting in a frame-shift mutation. e–g Complementation of the am1 mutant. The wild-type plants (left), am1 mutants (middle), and mutants transformed with pAM1 vector (right) are shown (color figure online)
Sequence analysis of AM1
AM1 was annotated in public databases (e.g., GRAMENE), as an encoding potassium efflux antiporter protein. AM1 contained twenty exons and nineteen introns. The open reading frame (ORF) of AM1 encodes a polypeptide of 1,155 amino acids with a calculated molecular mass of approximately 123 KD. Analysis of the AM1 protein sequence by ChloroP (Emanuelsson et al. 1999); http://www.cbs.dtu.dk/services/Signa-lP/) suggested a 41 amino acid chloroplast transit peptide (CTP) in the N-terminus (Fig. 4a). By searching the AM1 sequence against the Pfam database (Finn et al. 2010), two conserved domains were identified: one was Na_H_Exchanger located between amino acids 553 and 928, and another TrkA_N domain between amino acids 959 and 1,074 (Fig. 4a). Additionally, AM1 protein possessed ten transmembrane domains (Fig. 4b). Based on these features, the AM1 (LOC_Os04g58620) was grouped into the KEAIa family (Chanroj et al. 2012). In this family, Arabidopsis KEA1 and KEA2 were previously reported. AM1 shares very high peptide identity (65 %, 68 %) and similarity (76 %, 78 %) with both of KEA proteins (Fig. 4c). Phylogenetic tree analysis showed that AM1 homologs were well differentiated and exhibited a clear evolutionary linkage from unicellular green algae to angiosperms. The homologs from algae to mosses formed a subclade, while the proteins from angiosperms formed a large subclade where those from monocots and dicots were clearly divided (Fig. S3). However, most of them were not functionally characterized except AtKEA2, mainly as a K+/H+ antiporter (Aranda-Sicilia et al. 2012). Together, these findings suggested that AM1 functioned at the plastid membrane to control K+/H+ exchange.

Sequence analysis of AM1. a The normal AM1 protein contains three putative domain, including chloroplast transit peptides (CTP) in the N-terminal, Na_H_Exchanger domain and TrkA_N domain, whereas the am1 mutant protein only has CTP domain. b Prediction of ten transmembrane helices of AM1 protein by the program TMHMM available at http://www.cbs.dtu.dk/services/TMHMM/. c Alignment of AM1 with AtKEA 1 and AtKEA2. Sequences are for Oryza sativa LOC_Os04g58620 (AM1), Arabidopsis thaliana AtKEA1 (AT1G01790.1) and AtKEA2 (AT4G00630.2). Conserved amino acids are highlighted in shades of black. The predicted transit peptides, Na_H exchanger domain and KTN domains are labeled above the sequences by different lines. The Rossman fold motif is highlighted in red (color figure online)
Subcellular localization of AM1
Prediction of CTP in the N-terminus of AM1 by ChloroP and the chlorophyll-deficient phenotype observed in the am1 suggested a localization of AM1 in chloroplast. To examine the actual subcellular localization of AM1, an AM1-GFP construct was generated and then introduced into rice protoplasts by A. tumefaciens infiltration. Green fluorescent signals were visualized using confocal microscopy. Green fluorescent signals of AM1-GFP co-localized with the autofluorescent signals of chlorophyll indicating that AM1 targeted the rice chloroplasts (Fig. 5a–d).

Subcellular localization and expression analysis of AM1. Subcellular localization of AM1. Fluorescence signals were detected by confocal microscopy. a Green fluorescence from AM1-GFP fusion protein. b Red fluorescence of chloroplast autofluorescence. c Bright-field image under transmitted light. d Merged image of a, b and c. Bars represent 5 μm. e–i Histochemical staining shows that the p AM1::GUS reporter gene is ubiquitously expressed in roots, stems, leaves, leaf shoots, and panicles. j QRT-PCR analysis of AM1 in roots, stems, leaves, leaf shoots, and panicles of wild-type seedlings. The means and SD were calculated from three independent experiments. k Schematic illustration of a rice seedling with fully expanded third leaf. L1, L2, L3 and L4 indicate the first, second, third and fourth leaves, respectively. Developmental stages (P0–P6) are also indicated. SB (shoot base) corresponds to a 5 mm piece from the bottom of the shoot and contains as yet unmerged leaves at the P0–P3 stages. l QRT-PCR analysis of AM1 in the SB, L2, L3 and L4 tissues of wild-type seedlings. The means and SD were calculated from three independent experiments
Expression analysis of AM1
Data submitted to the microarray transcript profiling database indicated that the AM1 gene was widely expressed in rice, especially the leaves, throughout plant development ((Zimmermann et al. 2004); https://www.genevestigator.ethz.ch). To validate these data, we generated transgenic plants expressing a GUS reporter gene driven by the AM1 promoter (pAM1::GUS). Histochemical assay revealed that AM1 was highly expressed in the leaf, sheath, and panicle, while not in the root (Fig. 5e–i). This was confirmed by qRT-PCR using RNA samples from different parts of wild-type plants (Fig. 5j). We also analyzed AM1 expression in different sections of leaves at various leaf developmental stages (Fig. 5k). We found that AM1 was most highly expressed in the L3 section at the early chloroplast and leaf development stage (Fig. 5l). These results, together with chloroplast localization and the observed am1 phenotype, supported that AM1 played an important role in chloroplast development in rice leaves.
We next examined the transcription levels of the genes associated with chlorophyll biosynthesis, chloroplast development, or photosynthesis in the am1 mutant. Nine genes were selected, including genes that encode Rubisco large subunit (rbcL; Hwang and Tabita 1989), glutamyl-tRNA reductase (HEMA1; Kumar and Soll 2000), NADPH: protochlorophyllide oxidoreductase A (PORA; Holtorf and Apel 1996), light-harvesting Chl a/b-binding protein of PSII (CAB1R and CAB2R; Matsuoka 1990), YGL1 (Wu et al. 2007) and Mg-chelatase subunits (CHLD, CHLI and CHLH; Zhang et al. 2006). Most of these genes were all significantly down-regulated in the am1 mutant except YGL1 which was up-regulated in the mutant (Fig. 6a). These indicated that AM1 played an important role in regulating the expression of genes associated with chlorophyll biosynthesis, chloroplast development, and photosynthesis in the first few seedling leaves.

Analysis of gene transcription and protein accumulation associated with chlorophyll biosynthesis and photosynthesis in wild-type and am1. a Expression analysis of genes involved in chlorophyll biosynthesis, chloroplast development, or photosynthesis by QRT-PCR. Data represent mean ± SD from three independent biological replicates. Asterisks indicate statistically significant differences compared with wild-type (Student’s t test: **P < 0.01). b Western blot analysis of photosynthetic protein accumulation. The same filter was stained with Coomassie Brilliant Blue to visualize total proteins and the large subunit of Rubisco (RbcL) is indicated on the stained gel
As the mutation affects chloroplast ultrastructure and chloroplast-associated gene expression, we examined the accumulation of photosynthetic proteins in the mutant. These proteins included photosystem-I subunits (PsaA), photosystem-II subunits (D1), the ATP synthase subunits (AtpA and AtpB), RNA polymerase subunits (RopB) and the large subunit of soluble rubisco proteins (RbcL). The photosynthetic proteins AtpA and AtpB subunits were not detected in the am1 mutants. The levels of psaA and RbcL were reduced in am1 mutants compared with that of the wild-type. Meanwhile, the levels of psbA and RpoB were similar to those of the wild-type (Fig. 6b), indicating that the accumulation of most photosynthetic proteins was affected in am1 mutants due to defective chloroplast development.
Am1 show enhanced sensitivity to KCL at germination
Expression of AM1 under various abiotic stresses and treatments was examined. As shown in Fig. 7a, AM1 was induced under three treatments, including Nacl, Kcl and drought (PEG treatment). PEG treatment led to earlier expression peak of AM1 than salt treatments. Water and ABA treatments did not significantly alter AM1 expression. Since AM1 was predicted to be a potassium efflux antiporter (Fig. 4), we wondered whether am1 altered responses to Kcl. We examined the responses of am1 mutant to added Kcl at germination stage. Under the normal condition, the am1 mutant showed the similar germination rate to WT (Fig. 7b) but initiated germination with a 12 h delay relative to WT on MS medium supplemented with 100 mM Kcl (Fig. 7c), and 24 h delay on medium with 200 mM Kcl (Fig. 7d). These results suggested that AM1 played an important role in potassium transport.

Am1 show enhanced sensitivity to Kcl at germination. a Expression levels of AM1 seedlings in response to Nacl, Kcl, ABA and PEG using qRT-PCR. Five-week-old wild-type seedlings were immersed with their roots in Nacl (200 mM), Kcl (200 mM), ABA (100 μM), PEG (20 %) or H2O for the indicated times. After treatment, the aerial parts of seedlings were harvested and used for total RNA isolation. Germination rates of wild-type and am1 seeds in the absence (b), and presence of 100 mM (c), 200 mM (d) of Kcl. The germination was defined as the appearance of the white tip of the coleoptiles at least 5 mm long. Data represent mean ± SD from three independent biological replicates. Asterisks indicate statistically significant differences compared with the control (Student’s t test: *P < 0.05; **P < 0.01)
Enhanced tolerance to drought in am1 mutant
Induction by PEG treatments suggested that AM1 was likely involved in the drought tolerance. We investigated the effect of drought stress on the performance of am1 plants. Four-week-old plants were analyzed. Under the normal conditions, am1 plants grew normally and no significant differences were observed except for chlorosis. When water was withheld from these plants for 10 days, phenotypic changes were observed. WT plants exhibited severe symptoms of drought stress that the leaves became dry green, rolled and even withered; by contrast, these symptoms were substantially delayed in am1 plants (Fig. 8a). We also summarized the survival rate of the WT and am1 plants after 12 days of watering; more than 67.5 % of am1 plants recovered and grew normally whereas only about 12.5 % of WT plants recovered (Fig. 8b). These results indicated that am1 could increase the tolerance to drought at the seeding stage.

Enhanced tolerance to drought in am1 mutant. a Drought treatment of wild-type and am1 mutant plants. Twenty-day-old plants were subjected to drought stress for 12 days. b Survival rate of the plants shown in a after 12 days of recovery from drought stress. All non-treated plants (CK) show 100 % survival. Data represent mean ± SD from three replicates. Asterisks indicate statistically significant differences compared with the wild-type at P < 0.01. c Environmental scanning electron microscopy images of three levels of stomatal opening. Bar = 5 μm. d The percentage of three levels of stomatal opening in wild-type and am1 mutant (n = 78 stomata for wild-type; n = 89 stomata for am1). e DAB staining of the seedling leaves from wild-type and am1 mutant
Drought stress is known to be closely related to stomatal movements. We next examined the stomatal status in am1 and WT plants. The 4-week-old WT and am1 plants were exposed to drought stress for 1 week before they were used for stomatal analysis. As a result, 39.2 % of stomata were completely closed in the am1 mutant plants, while only 19.8 % in the wild-type plants. At the mean time, only 11.3 % of stomata were completely open in the am1 mutants, but 31.5 % in wild-type plants (Fig. 8c, d). The percentage of partially open stomata was similar in am1 mutant and wild-type plants (48.6–50.2 %). These results showed that stomatal movement was greatly affected in am1 mutants.
Since H2O2 induces leaf stomatal closure (Huang et al. 2009; Bright et al. 2006; Apel and Hirt 2004; Hamilton et al. 2000; McAinsh et al. 1996), we examined H2O2 level in the wild-type and am1 leaves by diaminobenzidine (DAB) staining and found a higher accumulation in am1 mutant plants (Fig. 8e). These results suggested that accumulation of H2O2 in am1 mutants probably underlied the increased stomatal closure conferring the drought tolerance.
Discussion
KEA genes encode putative K+ efflux antiporters that are predominantly found in algae and plants but rarely in metazoan. In the current study, we isolated the first KEA mutant am1 through map-based cloning in rice. The mutant allele displayed chlorophyll-deficient phenotype in the leaves suggesting an essential role in normal chloroplast development. In addition, am1 showed enhanced drought tolerance as compared to WT. Thus, KEAs played important role in abiotic stress tolerance.
KEA proteins constitute an ancient group of transporters homologous with bacterial transporters harboring a C-terminal KTN domain, the presence of which is a feature shared by most of bacterial K+ channels and transporters such as Kch, TrkA,Ybal, and KefB or KefC (Choe 2002). Its minimal functional unit is a dimeric molecule connected by a flexible hinge. Hinge move can be physically coupled with transmembrane loops controlling K+ flux (Choe 2002). Additionally, this domain contains the typical Rossman fold motif (G–X–G–X–X–G…D) involved in NAD binding, indicating metabolic control of transporter function (Roosild et al. 2002; Jiang et al. 2001). KEA family remained relatively constant at 3–7 genes from unicellular algae to flowering plants suggesting conserved roles and its functions are largely unknown (Chanroj et al. 2012). The sole functional information of KEA protein in plants comes from the ATKEA2 which functions at chloroplast as a cation/proton exchanger with preference for K+ and maintains the pH homeostasis (Aranda-Sicilia et al. 2012). In this present study, AM1 was classified into the same clades (KEAIa) as the ATKEA2 (Chanroj et al. 2012), suggesting their similar functions, i.e., the similar targeting to the chloroplast and mainly leaf expression in AM1 to ATKEA2 (Fig. 5). More recently, A two-pore K+ channel which controls photosynthetic light utilization has been found to be located in the thylakoid membrane of Arabidopsis (Carraretto et al. 2013), and the more precise localization of AM1 need to be further investigated. Although we failed to test the cation transport activity of AM1, the up-regulation of AM1 expression levels by salt induction as well as the enhanced sensitivity to Kcl at seed germination suggests that AM1 plays an important role in regulation of monovalent cation efflux to maintain the ion homeostasis in plant cells (Fig. 7).
Although bioinformatic analysis and proteomic studies suggested that KEA transporters in eukaryotes served functions mainly in the chloroplast or plastid of plant cells: firstly, up to date, all plants contained three subtypes of KEA from lower green algae to higher flowering plants (Chanroj et al. 2012); secondly, almost all the KEAs are found in plastid-containing organisms, ranging from cyanobacteria, green plants, red algae to secondary endosymbionts, but not in fungi; thirdly, some KEAIa proteins were previously identified in proteomic studies of the Arabidopsis chloroplast inner envelope (Ferro et al. 2010; Brautigam and Weber 2009; Zybailov et al. 2008). More recently, ATKEA2 was reported to target to chloroplasts in cotyledons (Aranda-Sicilia et al. 2012). Yet no genetic evidence and the phenotypic description of KEA mutants were provided to support this thesis. In our study, we isolated a KEA mutant designated am1 by fine mapping. Am1 mutant allele had a 2-bp deletion in 1th ORF and caused a premature stop codon which directly led to the loss of two main functional domains (Figs. 3e, 4a). Thus, am1 could be considered to lose most of the functions. Am1 mutant allele displayed a chlorophyll-deficient phenotype and abnormal chloroplast ultrastructure (Figs. 1, 2), In addition, am1 showed abnormal chloroplast gene expression and protein accumulation (Fig. 6). These findings provided the genetic evidence for KEA proteins involving in the chloroplast development. In the am1 mutants, the accumulation of photosystem-II subunits and the values of key photosynthetic parameters were similar to the wild-type (Fig. 6b; Table S2), indicating that photosystem II apparatus was not impaired in am1 mutant. Monovalent cation/proton antiporters controlling chloroplast development has been previously reported. The Arabidopsis CHX23 gene encodes a putative Na+ (K+)/H+ exchanger and functions in the adjustment of pH in the chloroplast and cytoplasm (Song et al. 2004). Leaves of chx23 mutant displayed pale-yellow with higher cytosolic pH than wild-type. Although the mechanism for AM1 affecting the chloroplast development might differ from that in CHX23, it provided a cue for further investigating the roles of AM1 in chloroplast development.
Drought is one of the prime abiotic stresses in the world and poses a serious threat to plant growth and seed production.To survive, plants have to respond to and adapt to the stresses. One of strategies is the reduction of stomatal opening to prevent water from losing. Reactive oxygen species (ROS) including hydrogen peroxide (H2O2) that were widely generated under various stress have been proposed to function as second messengers in abscisic acid (ABA) signaling in guard cells (McAinsh et al. 1996; Hamilton et al. 2000; Kohler et al. 2003; Bright et al. 2006; Zhang et al. 2001). H2O2 can also induce stomata closure independent of ABA signaling pathway (Huang et al. 2009). In plants, lots of cation/proton antiporters have been reported to regulate salt tolerance (Chanroj et al. 2012). However, none was showed to participate in drought tolerance. In this study, we showed that AM1 was induced by PEG and the am1 mutant plants could enhance the drought tolerance (Figs. 7a, 8a, b). Further analysis indicated that AM1 mutation resulted in a smaller stomatal opening and more accumulation of H2O2 (Fig. 8c–e). Previous studies showed that chloroplasts were the site of ABA biosynthesis (Seo and Koshiba 2002), and pigment-deficient mutants reduced endogenous level of ABA (Fambrini et al. 1993). Thus, increased stomatal closure in the am1 mutant probably was not due to endogenous ABA accumulation. Over-accumulation of H2O2 in am1 mutant most likely acted as an initiator of increased stomatal closure to confer the drought tolerance.
In summary, we isolated and characterized the first KEA mutant in rice. Our results not only provided a genetic evidence that KEA involved in the chloroplast development, but also revealed their novel function in the regulation of drought stress. However, it remains a challenge to identify their roles in ion transport and interpret molecular mechanism, as to how the AM1 affects the chloroplast development and drought stress.
Abbreviations
- Car:
-
Carotenoid
- Chl:
-
Chlorophyll
- KEA:
-
Potassium (K+) efflux antiporter
- ORF:
-
Open reading frame
- DAB:
-
Diaminobenzidine
- GFP:
-
Green fluorescent protein
References
Apel K, Hirt H (2004) Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol 55:373–399. doi:10.1146/annurev.arplant.55.031903.141701
Aranda-Sicilia MN, Cagnac O, Chanroj S, Sze H, Rodriguez-Rosales MP, Venema K (2012) Arabidopsis KEA2, a homolog of bacterial KefC, encodes a K(+)/H(+) antiporter with a chloroplast transit peptide. Biochim Biophys Acta 1818(9):2362–2371. doi:10.1016/j.bbamem.2012.04.011
Archer EK, Bonnett HT (1987) Characterization of a virescent chloroplast mutant of tobacco. Plant Physiol 83(4):920–925
Arnon DI (1949) Copper enzymes in isolated chloroplasts polyphenoloxidase in beta vulgaris. Plant Physiol 24(1):1–15
Booth IR (2003) Bacterial ion channels. Genet Eng (N Y) 25:91–111
Brautigam A, Weber AP (2009) Proteomic analysis of the proplastid envelope membrane provides novel insights into small molecule and protein transport across proplastid membranes. Mol Plant 2(6):1247–1261. doi:10.1093/mp/ssp070
Bright J, Desikan R, Hancock JT, Weir IS, Neill SJ (2006) ABA-induced NO generation and stomatal closure in Arabidopsis are dependent on H2O2 synthesis. Plant J 45(1):113–122. doi:10.1111/j.1365-313X.2005.02615.x
Carraretto L, Formentin E, Teardo E, Checchetto V, Tomizioli M, Morosinotto T, Giacometti GM, Finazzi G, Szabo I (2013) A thylakoid-located two-pore K+ channel controls photosynthetic light utilization in plants. Science 342(6154):114–118. doi:10.1126/science.1242113
Chanroj S, Wang G, Venema K, Zhang MW, Delwiche CF, Sze H (2012) Conserved and diversified gene families of monovalent cation/H(+) antiporters from algae to flowering plants. Front Plant Sci 3:25. doi:10.3389/fpls.2012.00025
Chen S, Tao L, Zeng L, Vega-Sanchez ME, Umemura K, Wang GL (2006) A highly efficient transient protoplast system for analyzing defence gene expression and protein–protein interactions in rice. Mol Plant Pathol 7(5):417–427. doi:10.1111/j.1364-3703.2006.00346.x
Choe S (2002) Potassium channel structures. Nat Rev Neurosci 3(2):115–121. doi:10.1038/nrn727
Dong H, Fei GL, Wu CY, Wu FQ, Sun YY, Chen MJ, Ren YL, Zhou KN, Cheng ZJ, Wang JL, Jiang L, Zhang X, Guo XP, Lei CL, Su N, Wang H, Wan JM (2013) A rice virescent-yellow leaf mutant reveals new insights into the role and assembly of plastid caseinolytic protease in higher plants. Plant Physiol 162(4):1867–1880. doi:10.1104/pp.113.217604
Emanuelsson O, Nielsen H, von Heijne G (1999) ChloroP, a neural network-based method for predicting chloroplast transit peptides and their cleavage sites. Protein Sci 8(5):978–984. doi:10.1110/ps.8.5.978
Fambrini M, Pugliesi C, Vernieri P, Giuliano G, Baroncelli S (1993) Characterization of a sunflower (Helianthus annuus L.) mutant, deficient in carotenoid synthesis and abscisic-acid content, induced by in vitro tissue culture. Theor Appl Genet 87(1–2):65–69. doi:10.1007/BF00223746
Ferro M, Brugiere S, Salvi D, Seigneurin-Berny D, Court M, Moyet L, Ramus C, Miras S, Mellal M, Le Gall S, Kieffer-Jaquinod S, Bruley C, Garin J, Joyard J, Masselon C, Rolland N (2010) AT_CHLORO, a comprehensive chloroplast proteome database with subplastidial localization and curated information on envelope proteins. Mol Cell Proteomics 9(6):1063–1084. doi:10.1074/mcp.M900325-MCP200
Finn RD, Mistry J, Tate J, Coggill P, Heger A, Pollington JE, Gavin OL, Gunasekaran P, Ceric G, Forslund K, Holm L, Sonnhammer EL, Eddy SR, Bateman A (2010) The Pfam protein families database. Nucleic Acids Res 38:D211–D222. doi:10.1093/nar/gkp985
Hamilton DW, Hills A, Kohler B, Blatt MR (2000) Ca2+ channels at the plasma membrane of stomatal guard cells are activated by hyperpolarization and abscisic acid. Proc Natl Acad Sci USA 97(9):4967–4972. doi:10.1073/pnas.080068897
Hiei Y, Ohta S, Komari T, Kumashiro T (1994) Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J 6(2):271–282
Holtorf H, Apel K (1996) Transcripts of the two NADPH protochlorophyllide oxidereductase genes PorA and PorB are differentially degraded in etiolated barley seedlings. Plant Mol Biol 31(2):387–392
Huang XY, Chao DY, Gao JP, Zhu MZ, Shi M, Lin HX (2009) A previously unknown zinc finger protein, DST, regulates drought and salt tolerance in rice via stomatal aperture control. Genes Dev 23(15):1805–1817. doi:10.1101/gad.1812409
Hwang SR, Tabita FR (1989) Cloning and expression of the chloroplast-encoded rbcL and rbcS genes from the marine diatom Cylindrotheca sp. strain N1. Plant Mol Biol 13(1):69–79
Jiang Y, Pico A, Cadene M, Chait BT, MacKinnon R (2001) Structure of the RCK domain from the E. coli K+ channel and demonstration of its presence in the human BK channel. Neuron 29(3):593–601
Jung KH, Hur J, Ryu CH, Choi Y, Chung YY, Miyao A, Hirochika H, An G (2003) Characterization of a rice chlorophyll-deficient mutant using the T-DNA gene-trap system. Plant Cell Physiol 44(5):463–472
Kohler B, Hills A, Blatt MR (2003) Control of guard cell ion channels by hydrogen peroxide and abscisic acid indicates their action through alternate signaling pathways. Plant Physiol 131(2):385–388. doi:10.1104/pp.016014
Kumar AM, Soll D (2000) Antisense HEMA1 RNA expression inhibits heme and chlorophyll biosynthesis in Arabidopsis. Plant Physiol 122(1):49–56
Kusumi K, Sakata C, Nakamura T, Kawasaki S, Yoshimura A, Iba K (2011) A plastid protein NUS1 is essential for build-up of the genetic system for early chloroplast development under cold stress conditions. Plant J 68(6):1039–1050. doi:10.1111/j.1365-313X.2011.04755.x
Maser P, Thomine S, Schroeder JI, Ward JM, Hirschi K, Sze H, Talke IN, Amtmann A, Maathuis FJ, Sanders D, Harper JF, Tchieu J, Gribskov M, Persans MW, Salt DE, Kim SA, Guerinot ML (2001) Phylogenetic relationships within cation transporter families of Arabidopsis. Plant Physiol 126(4):1646–1667
Matsuoka M (1990) Classification and characterization of cDNA that encodes the light-harvesting chlorophyll a/b binding protein of photosystem II from rice. Plant Cell Physiol 31(4):519–526
McAinsh MR, Clayton H, Mansfield TA, Hetherington AM (1996) Changes in stomatal behavior and guard cell cytosolic free calcium in response to oxidative stress. Plant Physiol 111(4):1031–1042
Rodermel S (2001) Pathways of plastid-to-nucleus signaling. Trends Plant Sci 6(10):471–478
Roosild TP, Miller S, Booth IR, Choe S (2002) A mechanism of regulating transmembrane potassium flux through a ligand-mediated conformational switch. Cell 109(6):781–791
Seo M, Koshiba T (2002) Complex regulation of ABA biosynthesis in plants. Trends Plant Sci 7(1):41–48
Song CP, Guo Y, Qiu Q, Lambert G, Galbraith DW, Jagendorf A, Zhu JK (2004) A probable Na+ (K+)/H+ exchanger on the chloroplast envelope functions in pH homeostasis and chloroplast development in Arabidopsis thaliana. Proc Natl Acad Sci USA 101(27):10211–10216. doi:10.1073/pnas.0403709101
Su N, Hu ML, Wu DX, Wu FQ, Fei GL, Lan Y, Chen XL, Shu XL, Zhang X, Guo XP, Cheng ZJ, Lei CL, Qi CK, Jiang L, Wang H, Wan JM (2012) Disruption of a rice pentatricopeptide repeat protein causes a seedling-specific albino phenotype and its utilization to enhance seed purity in hybrid rice production. Plant Physiol 159(1):227–238. doi:10.1104/pp.112.195081
Sugimoto H, Kusumi K, Tozawa Y, Yazaki J, Kishimoto N, Kikuchi S, Iba K (2004) The virescent-2 mutation inhibits translation of plastid transcripts for the plastid genetic system at an early stage of chloroplast differentiation. Plant Cell Physiol 45(8):985–996. doi:10.1093/pcp/pch111
Sugimoto H, Kusumi K, Noguchi K, Yano M, Yoshimura A, Iba K (2007) The rice nuclear gene, VIRESCENT 2, is essential for chloroplast development and encodes a novel type of guanylate kinase targeted to plastids and mitochondria. Plant J 52(3):512–527. doi:10.1111/j.1365-313X.2007.03251.x
ThordalChristensen H, Zhang Z, Wei Y, Collinge D (1997) Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley–powdery mildew interaction. Plant J 11(6):1187–1194
Wu Z, Zhang X, He B, Diao L, Sheng S, Wang J, Guo X, Su N, Wang L, Jiang L, Wang C, Zhai H, Wan J (2007) A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol 145(1):29–40. doi:10.1104/pp.107.100321
Yoo SC, Cho SH, Sugimoto H, Li J, Kusumi K, Koh HJ, Iba K, Paek NC (2009) Rice virescent3 and stripe1 encoding the large and small subunits of ribonucleotide reductase are required for chloroplast biogenesis during early leaf development. Plant Physiol 150(1):388–401. doi:10.1104/pp.109.136648
Zhang X, Zhang L, Dong F, Gao J, Galbraith DW, Song CP (2001) Hydrogen peroxide is involved in abscisic acid-induced stomatal closure in Vicia faba. Plant Physiol 126(4):1438–1448
Zhang H, Li J, Yoo JH, Yoo SC, Cho SH, Koh HJ, Seo HS, Paek NC (2006) Rice Chlorina-1 and Chlorina-9 encode ChlD and ChlI subunits of Mg-chelatase, a key enzyme for chlorophyll synthesis and chloroplast development. Plant Mol Biol 62(3):325–337. doi:10.1007/s11103-006-9024-z
Zhou K, Ren Y, Lv J, Wang Y, Liu F, Zhou F, Zhao S, Chen S, Peng C, Zhang X, Guo X, Cheng Z, Wang J, Wu F, Jiang L, Wan J (2013) Young leaf chlorosis 1, a chloroplast-localized gene required for chlorophyll and lutein accumulation during early leaf development in rice. Planta 237(1):279–292. doi:10.1007/s00425-012-1756-1
Zimmermann P, Hirsch-Hoffmann M, Hennig L, Gruissem W (2004) Genevestigator. Arabidopsis microarray database and analysis toolbox. Plant Physiol 136(1):2621–2632. doi:10.1104/pp.104.046367
Zybailov B, Rutschow H, Friso G, Rudella A, Emanuelsson O, Sun Q, van Wijk KJ (2008) Sorting signals, N-terminal modifications and abundance of the chloroplast proteome. PLoS One 3(4):e1994. doi:10.1371/journal.pone.0001994
Acknowledgments
This work was supported by the National Natural Science Foundation of China (31000667), National Transgenic Science and Technology Program (2011ZX08009 003), and the Major State Basic Research Development Program of China (973 Program, 2009CB8500).
Conflict of interest
The authors have declared that no conflict of interests exist.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Jeong Sheop Shin.
P. Sheng and J. Tan contributed equally to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Sheng, P., Tan, J., Jin, M. et al. Albino midrib 1, encoding a putative potassium efflux antiporter, affects chloroplast development and drought tolerance in rice. Plant Cell Rep 33, 1581–1594 (2014). https://doi.org/10.1007/s00299-014-1639-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00299-014-1639-y