
Abstract
Objective
Luteolin, a common dietary flavonoid, induces apoptosis of many types of cancer cells. However, its role in glioblastoma and the potential mechanisms remain unknown. In this research, we studied the molecular mechanisms of the anti-cancer effect of luteolin in glioblastoma cancer cell lines.
Methods
Both U251MG and U87MG human glioblastoma cell lines were tested. Cell growth was assessed by the cell counting kit-8. Cell apoptosis was detected with flow cytometry and caspase-3 immunofluorescence staining. The protein levels of caspase-3/Bax/Bcl-2 and p-PERK/p-eIF2α/ATF4/CHOP/caspase-12 pathway were analyzed using western blots. Reactive oxygen species generation was measured with DCFH-DA staining using flow cytometry. Mitochondrial membrane potential was tested with JC-1 staining. Anti-cancer effect in vivo was measured using tumor xenograft mode in nude mice.
Results
Luteolin induced a lethal endoplasmic reticulum stress response and mitochondrial dysfunction in glioblastoma cells by increasing intracellular reactive oxygen species (ROS) levels. Luteolin induced expression of ER stress-associated proteins, including phosphorylation of PERK, eIF2α, ATF4, CHOP and cleaved-caspase 12. Inhibition of ROS production by anti-oxidant N-acetylcysteine could reverse luteolin-induced ER stress and mitochondrial pathways activation as well as apoptosis. What’s more, we also showed the anticancer effect of luteolin in vivo.
Conclusions
Our results suggest that luteolin induces apoptosis through activating ER stress and mitochondrial dysfunction in glioblastoma cell lines and in vivo, which provides the anti-cancer candidate to treat glioblstoma.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Glioblastoma multiforme (GBM) is the most common and lethal malignant primary brain tumor. Even patients take the most aggressive treatment, they are challenged with poor feedback to surgery, vulnerable relapse and short survival time, due to the high degree of proliferation, invasion and resistance to chemotherapy and radiotherapy. The median survival time is only 3 months without any treatment, and 1–2 years with treatment since the time of diagnosis [1]. Therefore, new therapeutic strategies should be developed to inhibit the recurrence of cancer and to improve the prognosis of GBM patients.
Researches indicate that cancer cells have obviously higher levels of reactive oxygen species (ROS) under oxidative stress because of an imbalanced redox status compared with normal cells [2]. Up-regulated ROS levels can render cancer cells more sensitivity to agents, which increase ROS and oxidative stress [3, 4]. As we all know, apoptosis is one of major mechanisms for erasure of many cancer cells through different pathways, including endoplasmic reticulum (ER), mitochondrial pathway, or a death receptor-medicated apoptotic pathway [5]. Apoptosis is thought to be the principal mechanism of anti-cancer drugs killing cancer cells. Researches indicate that abnormalities in ER function can cause ER stress, resulting in unfolded protein response (UPR), which activated by three key signaling proteins: PKR-like endoplasmic reticulum kinase (PERK), inositol-requiring enzyme 1 (IRE1), and activating transcription factor 6 (ATF6) [6]. Although UPR signaling initially promotes cells survival during cellular stress, sustained and unabated ER stress will contribute to caspase-mediated apoptosis [7].
Flavonoids have a wide spectrum of pharmacological properties, including antioxidant and cancer preventive effect [8]. Among many types of flavonoids, luteolin (3′, 4′, 5, 7-tetrahydroxyflavone) is a common dietary form existing in many vegetables and fruits such as parsley, broccoli and celery [9, 10]. Researches imply that luteolin has anti-cancer effects including pro-apoptosis and cell cycle arrest, inhibition of metastasis and angiogenesis in vitro and vivo [11–13]. What’s more, it has been reported that luteolin could induce apoptosis through activating ER stress in neuroblastoma [7].
In this study, we investigated the molecular mechanisms of anti-cancer effects of luteolin in human glioblastoma cells in vitro and vivo. In the process, we discovered that luteolin could induce ER stress- and mitochondrial dysfunction mediated apoptosis.
Materials and methods
Cell culture procedures
Human glioblastoma U87MG and U251MG cell lines were obtained from the American Type Culture Collection (ATCC; Manassas, VA, USA) and grown in Dulbecco’s modified Eagle’s medium (DMEM) (Gibco, USA), containing 10% fetal bovine serum (FBS; Gibco, USA), 100 units/ml penicillin and 100 μg/ml streptomycin (HyClone, USA) at 37 °C and 5% CO2 incubator. Primary mouse cerebral vascular endothelial cells were prepared as described previously [14] and cultured in DMEM/F12 (Gibco, USA) containing 20% FBS (Gibco, USA) and 100 units/ml penicillin and 100 μg/ml streptomycin (HyClone, USA), were incubated at 37 °C in a humidified atmosphere of 95% air and 5% CO2.
Reagents and antibodies
Luteolin was purchased from Sigma (St. Louis, MO, USA). Anti-caspase-3, anti-Cleaved-caspase-3, anti-Bax, anti-Bcl-2, anti-eIF2α, anti-phosphorylated-eIF2α, anti-ATF4, anti-β-actin, goat anti-rabbit and anti-mouse IgG (H&L) horseradish peroxidase (HRP) secondary antibodies were purchased from Cell Signaling Technology (Danvers, MA, USA). Antibodies including anti-CHOP, anti-phosphorylated-PERK, anti-PERK, anti-caspase-12 were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Cell viability assay
Cell viability was assessed by Cell Counting Kit-8 (CCK-8) assay (Dojindo, Kumamoto, Japan). Briefly, the single cerebral vascular endothelial cells (CVE), U87MG and U251MG cells were seeded into 96-well plates at 5 × 103 cells per well and allowed to grow overnight to adhere. Luteolin was dissolved in DMSO and diluted with DMEM medium to final concentrations of 5, 10, 20, 40 and 80 μM. 48 h later, 10 μl CCK-8 reagent was added to per well followed by incubation for 3 h at 37 °C. The absorbance (value) at 450 nm wave length (OD450) was measured using Bio-Rad ELISA microplate reader (Bio-Rad Laboratories, CA, USA). The proliferation rate of cells = (1 − the OD450 values of treated groups/the OD450 values of control group) × 100%.
Western blot analysis
U87MG and U251MG cells were plated at a density of 4 × 105 (U87MG) and 3 × 105 (U251MG) cells in six-well plates or 35-mm dishes, respectively, and were allowed to grow overnight in DMEM containing 10% FBS. The cells were treated with different concentrations of luteolin. Then cells were harvested and lysed with solubilization buffer [50 mmol/l Tris–HCl (pH 7.6), 20 mmol/l MgCl2, 200 mmol/l NaCl, 0.5% NP40, 1 mmol/l DTT, and protease inhibitors] and centrifuged at 12,000g for 15 min at 4 °C. Protein concentrations were estimated by Coomassie Plus Protein Assay Reagent (Pierce, Rockford, IL). An equal amount of protein (20–100 μg) was subjected to 8–15% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE) and transferred to polyvinylidene fluoride (PVDF) membranes (Millipore, USA). With blocking with 5% nonfat dried milk for 1 h at room temperature (RT), the immunoreactive membranes were incubated overnight with the appropriate primary antibody as indicated overnight at 4 °C. After incubation with the appropriate secondary antibodies for 2 h at RT, the membranes were visualized with Chemiluminescent HRP Substrate (Millipore, USA). Relative changes in protein expression were estimated from the mean pixel density using Image J software, normalized to β-actin and presented as relative density units.
Immunofluorescence
U87MG and U251MG were seeded on glass coverslips in DMEM containing 10% FBS placed in six-well plates at a density of 1 × 105 per well for 24 h. The medium was then replaced by DMEM containing 10% FBS, and 40 μM luteolin was added. After further incubation for 12 or 24 h, the cells were fixed with 4% paraformaldehyde for 30 min at RT, permeabilized with 0.3% Triton X-100 for 10 min and blocked with goat serum (Boster, Wuhan, China) for 30 min at RT. Then cells were incubated with cytochrome C antibody overnight at 4 °C. Following washes with phosphate-buffered saline (PBS), the coverslips were incubated with a red-labeled secondary antibody (Wuhan Guge Biotechnology Co., Ltd, Wuhan, China) for 2 h at RT. Nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI). Slides were washed, then mounted, imaged under ZEISS fluorescence microscope.
Analysis of apoptosis
To detect and quantify apoptosis cells, TUNEL staining and subsequent flow cytometric were performed. As we all know, nuclei with fragmented DNA is characteristic of apoptosis. We performed a TUNEL assay using the ApopTag Plus Fluorescein In Situ Apoptosis Detection Kit (Promega, WI, USA) according to the manufacturer’s instructions. After treatment, cells were trypsinized, washed with PBS and suspended with 500 μl of binding buffer containing 5 μl of Annexin V-FITC (AV) and 5 μl of propidium iodide (PI) (BD Biosciences, CA, USA). Cells were subjected to flow cytometry assay with incubated for 15 min in the dark. Flow cytometry was performed using a FACSCanto 6-color flow cytometer (BD Biosciences, CA, USA).
Measurement of reactive oxygen species generation
Cellular reactive oxygen species (ROS) contents were measured by flow cytometry as described previously [15]. In short, 4 × 105 cells per well were plated on six-well plates. After overnight cells being attached to the plates, U251MG and U87MG cells were then exposed to luteolin for 6 h or pretreated with 5 mM NAC for 2 h. Then cells were stained with 10 μM DCFH-DA (Beyotime Biotech, China) at 37 °C for 30 min. U251MG and U87MG cells were collected and fluorescence was analyzed by FACSCalibur flow cytometer (BD Biosciences, USA).
Measurement of mitochondrial membrane potential
We used JC-1 (Beyotime Biotech, China) as specific probe to test the effects of luteolin on cell mitochondrial membrane potential (△ψ m). U87MG cells were treated with 40 μM luteolin for 12 h or pretreated with 5 mM NAC for 2 h. Then cells were stained with JC-1 at 37 °C cell incubator for 30 min. Images taken from monomer and aggregate were merged and viewed under ZEISS fluorescence microscope (40× amplification, Japan).
Tumor xenograft study
All procedures in animals were approved by the Animal Care and Use Committee of Nanjing University and conformed to Guide for the Care and Use of Laboratory Animals from National Institutes of Health. U87MG cells (5.0 × 106) were suspended in 100 μl PBS and then injected subcutaneously into right side of the anterior flank of the male BALB/c athymic nude mice (Charles River Breeding Laboratories, Wilmington, MA) (n = 5 for each group) at 4–6 weeks of age. Tumor growth was measured every 2 days using Vernier calipers and tumor volumes were determined by external measurements and calculated using the formula V = π/6 × larger diameter × (smaller diameter)2, as described by Lopez-lazaro [16]. The mean and standard error were then calculated for each experimental group for each time point. When the tumors reaching 70–100 mm3 in volume, intraperitoneal injections of DMSO or luteolin (10 mg/kg) were administered thrice per week for 35 days (15 times in total). Body weight was used to monitor the health of animals. At the end of the experiments, the animals were sacrificed and tumors were dissected and weighted.
Statistical analysis
Statistical analysis was performed using the SPSS19.0 software package. Comparisons between two groups were performed using the Student’s t test and ANOVA analysis was used between multiple groups. The results were expressed as the mean ± SEM. p value less than 0.05 was considered statistically significant.
Results
Luteolin induced apoptosis in human glioblastoma cancer cells
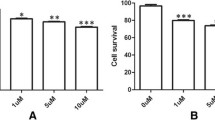
To study the effect of luteolin on the viability of human glioblastoma cancer cells, U251MG, U87MG cells and normal mouse vascular endothelial cells (VECs) were treated with different concentrations of luteolin (0, 5, 10, 20, 40, 80 μM) in vitro for 48 h and cells viability was tested by the CCK-8 assay. The results were showed in Fig. 1a. Luteolin decreased the cell viability in U251MG and U87MG dose-dependently. Importantly, luteolin did not obviously affect the cell viability of normal mouse vascular endothelial cells (VECs) at the dose up-to 40 μM.

Luteolin induced apoptosis in human glioblastoma cancer cells. a U251MG, U87MG cells and normal mouse vascular endothelial cells (VECs) were treated with luteolin at the indicated concentrations for 48 h, and cell viability was analyzed by CCK-8 assay. U251MG (b, c) and U87MG (d, e) cells were treated with luteolin at the indicated concentrations (0, 5, 10, 20, 40, 80 µM) for 48 h, and were evaluated by flow cytometry after staining with Annexin V-FITC and PI. All values represented the means ± SEM of three independent experiments. f, g Induction of apoptosis in human glioblastoma cells was tested by TUNEL staining after treatment with luteolin (40 μM) for 48 h. Green fluorescence mean positive results. h Luteolin increased the expression of cleaved caspase-3 in glioblastoma cells. The U251MG and U87MG cells were treated with luteolin for 48 h. The protein expression levels of cleaved caspase-3 was analyzed using western blot. β-Actin was used as an internal control. *,# p < 0.05, **,## p < 0.01 vs. control cells
To determine whether the inhibition of cell viability was due to apoptosis, we further examined the apoptotic effect of luteolin on human glioblastoma cells using Annexin V/propidium iodide (PI) staining assay. As shown in Fig. 1b–e, the U251MG and U87MG cells lines had shown a concentration-dependent apoptosis after 48 h treatment with luteolin. When luteolin was up to a concentration of 40 μM, the flow cytometry assays showed an apparent increase in apoptosis of both of U251MG and U87MG. In addition, TUNEL staining results also indicated that luteolin could induce apoptosis of U251MG and U87MG glioblastoma cancer cells (Fig. 1f, g). Then we determined the levels of apoptosis-related proteins in U251MG and U87MG cells after exposure to luteolin. The Fig. 1h showed that treatment with luteolin for 48 h increased the level of cleaved-caspase-3. We thought that lueolin could induce apoptosis of glioblastoma cancer cell lines. Hence, we should research the possible reasons.
ROS generation was the regulator of luteolin-induced apoptosis
It was believed that luteolin could induce apoptosis resulting from ROS-mediated late endoplasmic reticulum stress and mitochondrial dysfunction in Neuro-2a mouse neruoblastoma cells [7]. Therefore, we tested the role of ROS in luteolin-induced apoptosis in human glioblastoma cancer cells. As shown in Fig. 2a–f, treatment with 40 μM luteolin in U251MG and U87MG cancer cells for 6 h could significantly increase DCF-reactive ROS level. What’s more, pretreatment with the antioxidant 5 mM NAC (N-acetyl cysteine) for 2 h would partly inhibited luteolin up-regulated ROS level. In addition, the results showed that NAC pretreatment for 2 h could also abolish 40 μM luteolin induced-apoptosis (Fig. 2g, h).

ROS generation was the regulator of luteolin-induced apoptosis. Intracellular ROS generation induced by luteolin was measured in U251MG (a) or U87MG (b) cells by staining with DCFH-DA (10 µM) for 30 min. Fluorescence images were acquired by fluorescence microscopy. Intracellular ROS levels were measured by flow cytometry. c U251MG; d U87MG. e, f Flow cytometry results from c and d were calculated and represented as the percent of control. g, h Cell apoptosis was measured by flow cytometry by staining with Annexin V-FITC and PI. a–h NAC partly blocked ROS generation induced by luteolin. Cells were pretreated with or without 5 mM NAC for 2 h before exposure to luteolin for 6 h. Data presented were representative of three independent experiments. All values represented the means ± SEM. *p < 0.05, **p < 0.01
Luteolin-induced ROS increases ER stress and mitochondrial dysfunction, which contributes to luteolin lethality in glioblastoma cells
Researches showed that increased ROS level in the intracellular redox status would increase the levels of unfolded proteins in the ER and resulted in ER stress response [15, 17, 18]. ER stress could phosphorylate protein kinase R-like ER kinase (PERK), then PERK medicated the phosphorylation of eukaryotic initiation factor-2α(p-eIF2α), which would block cap-dependent protein translation but allow preferential translation of ATF4. Then, ATF4 could induce the prodeath transcriptional regulator CHOP [6, 19]. So, we examined the protein expressions level of ER stress-related proteins, such as p-PERK, p-eIF2α, ATF4, CHOP and caspase-12 in U251MG and U87MG cancer cells after the treatment of luteolin. The time-course results indicated that 40 μM luteolin could significantly activate ER stress. The protein levels of p-PERK, p-eIF2α and ATF4 reached the peak after 6 h of treatment, while CHOP, cleaved-caspase-12 and cleaved-caspase-3 did at 24 h after treatment in U251MG glioblastoma cells (Fig. 3a). Dose results indicated that different concentrations of luteolin also increased the expression of p-PERK, p-eIF2α, ATF4 and CHOP in U251MG and U87MG cancer cells (Fig. 3b, c). In addition, we found that pretreatment of NAC could block the up-regulated expression of p-eIF2α, ATF4 and CHOP in U251MG glioblastoma cells (Fig. 3d). These results showed that luteolin-induced ROS accumulation could result in induction of ER stress, with the up-regulated level of p-PERK, p-eIF2α, ATF4, CHOP and cleaved-caspase-12 in glioblastoma cancer cells.

Luteolin-induced ROS increases ER stress in human glioblatoma cancer cells. a Cells were exposed to 40 μM luteolin for indicated times. b, c U251MG and U87MG cancer cells were treated with different concentration of luteolin for 24 h. d NAC partly reversed the activation of ER stress induced by 40 μM luteolin. U251MG cells were pretreated with or without 5 mM NAC for 2 h before exposure to luteolin for 6 h. a–d The expressions of phosphorylated-PERK, PERK, phosphorylated-eIF2α, eIF2α, ATF4, CHOP, cleaved-caspase-12 and cleaved-caspase-3 were detected by western blot. Data presented were representative of three independent experiments
We also know that mitochondria plays a central role in regulation of cell apoptosis [20]. Excessive generation and accumulation of ROS results the cell oxidatively stressed and impairs membrane proteins, which result in mitochondrial dysfunction. Researches indicate that loss of mitochondrial membrane potential (△ψ m) is disastrous for cells and results in the release of cytochrome C to the cytosol [21], which leads to apoptosis. So, we examined the mitochondrial membrane potential after treatment of 40 μM luteolin for 12 h using JC-1 as a molecular probe by fluorescence microscope analysis. The integrity of mitochondrial membranes potential decreased in U87MG cells after treatment of 40 μM luteolin as we observed the elevation of green fluorescence from the red to green (Fig. 4a). On top of that, the release of cytochrome C to cytoplasm was elevated in U251MG cells by immunofluorescence (Fig. 4b). The imbalance between anti-apoptotic and pro-apoptotic Bcl-2 family proteins would lead to the decrease of mitochondrial membrane potential and cause apoptosis. So we tested the protein expression of Bcl-2 and Bax. As shown in Fig. 4c, treatment with luteolin for 24 h significantly decreased the expression of anti-apoptotic protein Bcl-2 and increased the expression of pro-apoptotic protein Bax in U251MG and U87MG glioblastoms cells. Additionally, pretreatment with 5 mM NAC for 2 h could decrease the up-regulated expression of Bax and increase the expression of inhibited-Bcl-2 (Fig. 4d). These results indicated that luteolin treatment induced ROS-dependent mitochondrial apoptosis in glioblastoma cancer cells.

Luteolin activated mitochondrial apoptotic pathway. a U251MG cells were treated with luteolin, obviously decreasing the mitochondrial membrane potential (△ψ m). U87MG cells after treatment with luteolin (40 μM) for 12 h were stained with JC-1 and analyzed by fluorescence microscope. b U251MG cells after treatment of luteolin (40 μM) for 12 h were stained with anti-cytochrome C and DAPI for detecting the expression of cytochrome C. c, d Western blot analysis effects of luteolin on expression of Bcl-2 and Bax in U251MG (c, d) and U87MG (c) cancer cells. All images shown here were representative of three independent experiments with similar results
Luteolin inhibited tumor growth in a xenograft mouse model
To evaluate the anti-glioblastoma effect of luteolin in vivo, immune-deficient nude mice bearing U87MG tumor xenografts were employed. When tumors reaching 70–100 mm3, 0.1% DMSO or luteolin (10 mg/kg) was administered by intraperitoneal injection thrice per week for 35 days (15 times in total). As showed in Fig. 5a, b treatment with luteolin inhibited U87MG tumor xenografts growth. Moreover, luteolin did not affect body weight, alanine aminotransferase (ALT) or aspartate transaminase (AST) obviously (Fig. 5c, d). The in vivo mechanistic studies revealed that luteolin treatment inhibited tumor xenografts by induction of ER stress-associated proteins, ATF4, CHOP and cleaved-caspase-12 and caspase-3 activation (Fig. 5e). In addition, the TUNEL staining of tumor xenografts tissue also showed that luteolin resulted in apoptosis (Fig. 5f). These data suggested that ER stress and apoptosis were involved in luteolin-induced suppression of tumor growth.

Luteolin inhibited tumor growth in a xenograft mouse model. Luteolin treatment inhibited tumor volume (a, b) of U87MG human glioblastoma cancer xenografts in nude mice, but did not affect body weight (c) and alanine aminotransferase (ALT) or aspartate transaminase (AST) (d) of mice. e Western blot analysis on the expression of ATF4, CHOP, cleaved-caspase-12, Bax and cleaved-caspase-3 from respective tumor tissue lysates. β-Actin was used as protein loading control. f TUNEL staining from respective tumor tissue. All values represented the means ± SEM. *p < 0.05, **p < 0.01
Discussion
Glioblastoma multiforme are the most common and lethal intracranial tumors, which exhibit a relentless malignant progression characterized by widespread invasion throughout the brain, resistance to traditional and newer targeted therapeutic approaches, and certain death. Temozolomide is the most commonly employed in clinic either alone or in combination with radiotherapy. However, some GBM patients develop resistance and relapse after a few months of treatment. Therefore, there is an urgent need for additional preventive and therapeutic drugs against GBM.
Das et al. showed that treatment of glioblastoma cells (T98G and U87MG) with flavonoids, such as apigenin, epigallocatechin, epigallocatechin-3-gallate and genistein triggered ROS production that in turn induced apoptosis with phosphorylation of p38 MAPK and activation of the redox-sensitive JNK1 pathway [22]. In this study, we found for the first time that luteolin could induce glioblastoma cancer cells apoptosis in vitro and in vivo through ER stress and mitochondrial dysfunction. It has been shown that luteolin is capable of inhibiting cell proliferation and inducing apoptosis in many types of cancer cells, for instance prostate cancer, breast cancer, lung cancer and hepatoma cells [10, 23]. Literature has reported that luteolin exert chemosensitizing effects on apoptosis in cancer cells [24]. However, the effect of luteolin on glioblastoma cancer cells and its action mechanisms for the anti-cancer effect have not been well elucidated.
Researches indicate that ROS plays an important role in the process of cancer genesis and metastasis. Cancer cells always have higher levels of ROS and higher antioxidant activities in an uncontrolled status compared with normal cells [25], which result in the lack of ability to cope with additional oxidative stress and vulnerability to ROS [26]. Therefore, targeting ROS is an important therapeutic strategy for cancer drugs such as cisplatin, paclitaxel [27, 28]. Exploring the mechanisms of ROS-based treatment is required for further improving the efficacy and specificity of cancer drugs.
In response to oxidative stress medicated by ROS, cells adapted to accumulation of unfolded and misfolded proteins by ER stress. ER stress protects cell by shutting-down the general protein synthesis and increasing the expression of molecular chaperones such as ER resident hsp70 homologue and glucose-regulated protein 78 (GRP78) [19]. When ER stress is prolonged, the pro-death ER stress pathways mediated by CHOP (CAAT/enhancer binding protein homologous protein) and caspase-12 activated [29]. ER stress-induced apoptosis is a key pathologic event of anti-tumor effects in many cancers [30, 31]. Researches have reported activated caspase-12 localized in the ER exerting the pro-apoptotic effect of ER stress [32–34]. Our results showed that p-PERK, p-eIF2α and ATF4 were up-regulated in a time-dependent manner after treatment of luteolin, indicating that ER stress was activated at 3–6 h. What’s more, the ER stress associated apoptotic pathway protein CHOP and cleaved-caspase 12 increased at 6–24 h after luteolin treatment in U251MG cells. In addition, p-PERK, p-eIF2α, ATF4 and CHOP was also up-regulated in a dose-dependent manner. Furthermore, we found that NAC could block the luteolin-induced activation of ER stress proteins, such as p-eIF2α, ATF4 and CHOP, which revealed that accumulated ROS level was the upstream regulator of luteolin-induced ER stress in glioblastoma cells. We suggest that generation of reactive oxygen species is an early event, which initiates ER stress-induced apoptotic process and late mitochondrial dysfunction in luteolin-treated glioblastoma cancer cell lines. Imperfectly we did not further explore the role of ER stress in luteolin-induced cell apoptosis in GBM cells. What’s more, in our study, both ER stress and mitochondrial were implicated in luteolin-induced cell apoptosis, however, it was not well understood the cross-talking between ER stress and mitochondrial dysfunction in luteolin-induced apoptosis pathways.
In conclusion, we studied the anti-proliferative and pro-apoptotive effects and mechanisms of luteolin in glioblastoma cancer cell lines. We found that treatment of luteolin resulted in severe ROS accumulation, which activated the ER stress and mitochondrial apoptotic pathways. We also demonstrated the effect of luteolin on suppression of tumor growth in vivo by a xenograft tumor model.
References
Wen PY, Kesari S (2008) Malignant gliomas in adults. N Engl J Med 359:492–507
Trachootham D, Alexandre J, Huang P (2009) Targeting cancer cells by ROS-mediated mechanisms: a radical therapeutic approach? Nat Rev Drug Discov 8:579–591
Yang J, Lu M, Lee C, Chen G, Lin Y, Chang F et al (2011) Selective targeting of breast cancer cells through ROS-mediated mechanisms potentiates the lethality of paclitaxel by a novel diterpene, gelomulide K. Free Radic Biol Med 51:641–657
Nogueira V, Hay N (2013) Molecular pathways: reactive oxygen species homeostasis in cancer cells and implications for cancer therapy. Clin Cancer Res 19:4309–4314
Ghobrial IM, Witzig TE, Adjei AA (2005) Targeting apoptosis pathways in cancer therapy. CA Cancer J Clin 55:178–194
Tabas I, Ron D (2011) Integrating the mechanisms of apoptosis induced by endoplasmic reticulum stress. Nat Cell Biol 13:184–190
Choi AY, Choi JH, Yoon H, Hwang K, Noh MH, Choe W et al (2011) Luteolin induces apoptosis through endoplasmic reticulum stress and mitochondrial dysfunction in Neuro-2a mouse neuroblastoma cells. Eur J Pharmacol 668:115–126
Neuhouser ML (2004) Dietary flavonoids and cancer risk: evidence from human population studies. Nutr Cancer 50:1–7
Mencherini T, Picerno P, Scesa C, Aquino R (2007) Triterpene, antioxidant, and antimicrobial compounds from Melissaofficinalis. J Nat Prod 70:1889–1894
Lin Y, Shi R, Wang X, Shen HM (2008) Luteolin, a flavonoid with potential for cancer prevention and therapy. Curr Cancer Drug Targets 8:634–646
Cai X, Ye T, Liu C, Lu W, Lu M, Zhang J et al (2011) Luteolin induced G2 phase cell cycle arrest and apoptosis on non-small cell lung cancer cells. Toxicol In Vitro 25:1385–1391
Wu B, Zhang Q, Shen W, Zhu J (2008) Anti-proliferative and chemosensitizing effects of luteolin on human gastric cancer AGS cell line. Mol Cell Biochem 313:125–132
Chian S, Thapa R, Chi Z, Wang XJ, Tang X (2014) Luteolin inhibits the Nrf2 signaling pathway and tumor growth in vivo. Biochem Biophys Res Co 447:602–608
Nakagawa S, Deli MA, Kawaguchi H, Shimizudani T, Shimono T, Kittel A et al (2009) A new blood-brain barrier model using primary rat brain endothelial cells, pericytes and astrocytes. Neurochem Int 54:253–263
Zou P, Zhang J, Xia Y, Kanchana K, Guo G, Chen W et al (2015) ROS generation mediates the anti-cancer effects of WZ35 via activating JNK and ER stress apoptotic pathways in gastric cancer. Oncotarget 6:5860–5876
Zhang L, Wang H, Ding K, Xu J (2015) FTY720 induces autophagy-related apoptosis and necroptosis in human glioblastoma cells. Toxicol Lett 236:43–59
Zhang K, Kaufman RJ (2008) From endoplasmic-reticulum stress to the inflammatory response. Nature 454:455–462
Hotamisligil GKS (2010) Endoplasmic reticulum stress and the inflammatory basis of metabolic disease. Cell 140:900–917
Walter P, Ron D (2011) The unfolded protein response: from stress pathway to homeostatic regulation. Science 334:1081–1086
Desagher S, Martinou J (2000) Mitochondria as the central control point of apoptosis. Trends Cell Biol 10:369–377
Olichon A, Baricault L, Gas N, Guillou E, Valette A, Belenguer P et al (2003) Loss of OPA1 perturbates the mitochondrial inner membrane structure and integrity, leading to cytochrome c release and apoptosis. J Biol Chem 278:7743–7746
Das A, Banik NL, Ray SK (2010) Flavonoids activated caspases for apoptosis in human glioblastoma T98G and U87MG cells but not in human normal astrocytes. Cancer Am Cancer Soc 116:164–176
Selvendiran K (2006) Luteolin promotes degradation in signal transducer and activator of transcription 3 in human hepatoma cells: an implication for the antitumor potential of flavonoids. Cancer Res 66:4826–4834
Shi RX, Ong CN, Shen HM (2004) Luteolin sensitizes tumor necrosis factor-alpha-induced apoptosis in human tumor cells. Oncogene 23:7712–7721
Szatrowski TP, Nathan CF (1991) Production of large amounts of hydrogen peroxide by human tumor cells. Cancer Res 51:794–798
Gorrini C, Harris IS, Mak TW (2013) Modulation of oxidative stress as an anticancer strategy. Nat Rev Drug Discov 12:931–947
Bragado P, Armesilla A, Silva A, Porras A (2007) Apoptosis by cisplatin requires p53 mediated p38α MAPK activation through ROS generation. Apoptosis 12:1733–1742
Alexandre J, Hu Y, Lu W, Pelicano H, Huang P(2007) Novel action of paclitaxel against cancer cells: bystander effect mediated by reactive oxygen species. Cancer Res 67:3512–3517
Rao R, Nalluri S, Fiskus W, Savoie A, Buckley KM, Ha K, et al (2010) Role of CAAT/enhancer binding protein homologous protein in Panobinostat-mediated potentiation of Bortezomib-induced lethal endoplasmic reticulum stress in mantle cell lymphoma cells. Clin Cancer Res 16:4742–4754
Oakes SA (2017) Endoplasmic reticulum proteostasis: a key checkpoint in cancer. Am J Physiol Cell Physiol 312:C93–C102
Kim AD, Madduma HS, Piao MJ, Kang KA, Cho SJ, Hyun JW (2015) Esculetin induces apoptosis in human colon cancer cells by inducing endoplasmic reticulum stress. Cell Biochem Funct 33:487–494
Boyce M, Yuan J (2006) Cellular response to endoplasmic reticulum stress: a matter of life or death. Cell Death Differ 13:363–373
Ge W, Yin Q, Xian H (2015) Wogonin induced mitochondrial dysfunction and endoplasmic reticulum stress in human malignant neuroblastoma cells via IRE1alpha-dependent pathway. J Mol Neurosci 56:652–662
Wu CT, Weng TI, Chen LP, Chiang CK, Liu SH (2013) Involvement of caspase-12-dependent apoptotic pathway in ionic radiocontrast urografin-induced renal tubular cell injury. Toxicol Appl Pharmacol 266:167–175
Acknowledgements
The authors thank Dr Han Yanling for the technical assistance. This work was supported by Grants from the National Natural Science Foundation of China (No. 81371357) and China Postdoctoral Science Foundation funded project under Grant (No. 2014M562665).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors disclose no potential conflicts of interest.
Rights and permissions
About this article

Cite this article
Wang, Q., Wang, H., Jia, Y. et al. Luteolin induces apoptosis by ROS/ER stress and mitochondrial dysfunction in gliomablastoma. Cancer Chemother Pharmacol 79, 1031–1041 (2017). https://doi.org/10.1007/s00280-017-3299-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00280-017-3299-4