
Abstract
The songs of migratory passerine birds have a key role in mate attraction and territory defence during the breeding season. Many species also sing on their wintering grounds, but the function of this behaviour remains unclear. One possible explanation, proposed by the song improvement hypothesis, is that the birds take advantage of this period to develop their singing skills for the next breeding season. If so, non-breeding songs should reflect features of an early phase in song development, characterized by high vocal plasticity. In our study, we tested this prediction by comparing songs of thrush nightingales (Luscinia luscinia) recorded at two different breeding areas in Europe and one wintering area in Africa. While all songs from European localities had a typical structure characteristic of the study species, 89% of the songs recorded from Africa were highly variable, lacking such typical structure. We conducted further detailed analysis of breeding and winter songs that exhibited species-specific structure. First, we explored plasticity at the syllable level using a cross-correlation analysis, to obtain similarity scores as a measure of consistency. Second, we asked multiple human observers to quantify element variability. Our results showed significant differences in syllable consistency between breeding and wintering grounds, with more consistent delivery of syllables in the breeding areas. Likewise, element variability was substantially lower in the breeding populations. While both results fit the predictions of the song improvement hypothesis, more research is needed to elucidate the roles of singing on the wintering grounds.
Significance statement
Many migratory songbirds sing on their wintering grounds, outside the breeding period. While the role of singing during breeding has been broadly studied, our understanding of the function of winter singing remains limited. We analysed songs of the thrush nightingale, a migratory songbird with highly complex songs, comparing song structures recorded from breeding populations in Europe and an African wintering site. We demonstrate that males recorded at wintering locations sang songs with both significantly lower syllable consistency and higher element variability. Such characteristics are comparable to those observed during the sensorimotor phase of song development, previously described in other species. This pattern supports the song improvement hypothesis, suggesting that males singing on the wintering grounds may practice songs for the next breeding season. This study contributes to the understanding of the functions of songbird vocal behaviour out of the breeding context.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Since Darwin published his book on selection in relation to sex (Darwin 1871), biologists have been intrigued by the functions of sexually selected characters. According to the sexual selection theory, two processes are important for such traits: male-male competition leading to armaments and female choice leading to ornaments (Berglund et al. 1996). Such traits have been intensively studied in the context of reproduction, but we have little knowledge about their functions outside of the breeding context. Birdsong is an example of such a sexually selected character. Males of most songbird species (oscines) that breed in the temperate zone sing intensively during the breeding period to repel rivals and to attract females. The singing performance of male individuals is thus a crucial component in estimating their quality for both competitors and potential partners (e.g. Catchpole and Slater 2008; Gil and Gahr 2002). While this behaviour has been well studied during the breeding period, why birds sing outside the breeding season remains an open question (Sorensen et al. 2016). Long-distance migrants spend a significant part of the year wintering at distant locations. Despite accumulating evidence of songbirds singing on their wintering grounds (e.g. Alexander 1917; Bannerman 1931; Cramp and Perrins 1994), research on this topic is still scarce.
A few studies investigating singing on the wintering grounds suggest that this behaviour may serve for territorial defence (e.g. marsh warbler, Acrocephalus palustris, Kelsey 1989), while others could not confirm such a territorial function (e.g. willow warbler, Phylloscopus trochilus, Sorensen 2014). Sorensen et al. (2016) tested several hypotheses related to the role of winter singing in great reed warblers (A. arundinaceus). First, they showed that seasonal variation in testosterone levels seems not to be responsible for triggering the singing behaviour. Second, no clear territorial behaviour was observed outside the breeding season. As an alternative explanation, the authors thus proposed the “song improvement hypothesis”, suggesting that practicing complex songs during the non-breeding season could improve song performance for the following breeding season, and thus positively impact individual fitness.
Unlike many other sexually selected traits, the song of songbirds is a learned behaviour that is transmitted culturally (Catchpole and Slater 2008; Zeigler and Marler 2008). Young songbirds store model songs heard from conspecifics shortly after hatching during an early sensitive phase. They then gradually adjust their vocal output to what was stored before (Brainard and Doupe 2002). During this sensorimotor phase, the song is first characterized by high vocal plasticity (‘plastic song’), and then gradually develops into more consistent songs (Brenowitz et al. 1997). Eventually, the song crystallizes and reaches its final adult and generally stereotyped form (‘full song’ or ‘crystallized song’; Brainard and Doupe 2002). However, even after crystallization, the song of adult males may retain some plasticity. Such retention has been documented in adult males recapitulating plastic song later in life during an annual sensorimotor phase occurring during the non-breeding season (Nottebohm et al. 1986; Margoliash et al. 1991). The capacity to retain some plasticity may allow the birds to refine, improve and adjust their song motor performance even in adulthood (Sakata and Vehrencamp 2012).
Song improvement via practice thus seems to require adult birds to retain or temporarily regain some plasticity in their song. Consistent with this idea, Blyth’s reed warblers (A. dumetorum) recorded during spring migration produced songs with less organized syntax and less stable syllable combinations compared to populations from breeding areas (Marova et al. 2010). Likewise, common nightingales (Luscinia megarhynchos) exhibited higher degrees of song variability in the wintering population compared to those at breeding localities (Kipper et al. 2016). However, neither of these studies investigated vocal plasticity by directly measuring syllable consistency. Syllable consistency is defined as the ability to replicate the acoustic features of a song from one rendition to the next (Sakata and Vehrencamp 2012). Consistent songs are expected to be adaptive since they send a more efficient signal with a higher-performance value, as high vocal consistency has repeatedly been linked to age, social dominance and reproductive success in several species (reviewed in Sakata and Vehrencamp 2012), and thus should be particularly important during the breeding season. Likewise, the plasticity of songs may be reflected in the variability of elements (the smallest units of a song), i.e. the acoustic structure of individual elements may vary from one rendition to the next. High variability of elements is thus evidence for low vocal consistency.
In our study, we compare the syllable consistency and element variability in thrush nightingales (Luscinia luscinia) recorded during the breeding and wintering period, respectively. We assume that if a song improvement process is at work, it should affect the consistency in the fine structure of the song resulting in less stereotyped songs. We explore the acoustic variation between renditions within each individual’s performance (i.e. in one and the same recording), allowing us to investigate the plasticity of song and the variability of its elements and compare if these individual song patterns differ between populations from breeding and wintering sites. Following the song improvement hypothesis, we predict that individuals recorded at the non-breeding site should display a higher level of vocal plasticity, and thus less stereotyped songs with a lower consistency in syllable performance than those from breeding grounds. Additionally, higher song plasticity might also result in higher element variability, as different reiterations (or rendition) of one element type might lack sufficient consistency in order to be perceived as the same type. To our knowledge, this is the first detailed quantitative analysis of wintering song based on spectral and temporal song characteristics. Such a structural approach may contribute to further understanding of the evolution and functions of non-breeding singing in migrating species.
Methods
Studied species and its song
The thrush nightingale is a long-distance migrating songbird with complex songs (e.g. repertoire varying from 23 to 42 songs, mean 32.7 per individual, Griessmann and Naguib 2002), characterized by a typical two-part ending phrase consisting of slow syllable repetitions, usually followed by a fast rattling part. This specific composition remains stable over the whole species range (Sorjonen 1983; Griessmann and Naguib 2002; Vokurková et al. 2013; Ivanitskii et al. 2016; cf. Fig. 1a). Closely related to the well-studied common nightingale, thrush nightingales seem to have the same ability to learn new songs even after their early life sensory phase and to adjust their song repertoire according to their neighbours (Sorjonen 1987; Griessmann and Naguib 2002). In secondary contact zones, thrush nightingales sometimes even learn from their sister species, the common nightingale (Vokurková et al. 2013). Learning after their early life sensory phase as well as learning from their sister species can have direct consequences on their vocal territorial interactions during the breeding season (Naguib and Todt 1998; Reif et al. 2015; Souriau et al. 2018). As a result, the mean proportion of song types shared between breeding males can be as high as 47% in a stable population (Ivanitskii et al. 2016). Moreover, syllable sharing seems to be even more pronounced: Griessmann and Naguib (2002) showed that birds from the same population shared up to 80% of their syllable types but only 30% of song types.

a Spectrogram of a typical thrush nightingale song (recorded in Poland) showing the characteristic song organisation of the species. The sub-parts (indicated by Greek characters and highlighting the ending “castanet” and “rattling” parts as introduced by Sorjonen 1983) are characterized by elements that always occur in the same sequence. The α-part is usually the most variable part between and within individual repertoires, while the last parts of the song (solid red arrow lines) are the most stable ones. In this study, song type categories are defined as the same combination of the last β element and the following Ψ1 (Castanet) and Ψ2 (Rattling, fast trill covering a wide frequency range), indicated by solid arrow lines underneath the spectrogram. b–g Spectrograms illustrating syllable renditions selected for the spectral cross-correlation analysis and their sequential position: b, c Poland; d, e Russia; f, g Tanzania. Highlighted syllables illustrate renditions selected for comparison of the same syllable type in two songs of the same type and from the same individual
Every year, thrush nightingales migrate from Northern and Eastern Europe to East and South Africa (Pearson and Backhurst 1976), where they are known to sing actively (Pearson 1984), though their territorial habits and social interactions at their winter grounds remain poorly known. More recent studies investigating migration using geolocators suggest that birds from the same population may migrate to similar wintering regions (Stach et al. 2012; Thorup et al. 2017).
Also, males are highly philopatric to their breeding grounds (e.g. return rates up to 40% in Scandinavia, Sorjonen 1987), implying that the same individuals may continue to engage in vocal interactions in subsequent years.
Studied populations
This study is based on a comparison between three distant populations (i.e. males recorded in the same area) of thrush nightingales sampled over different years. All recordings presented “pure-species” songs, i.e. songs without any heterospecific components (elements from the closely related common nightingale, Vokurková et al. 2013). Populations from breeding grounds in Europe were recorded in Poland, Podlasie (53°12.0′N, 22°24.4′E), during spring 2009, and in Russia, Zvenigorod, Moscow region (55°42.3′N, 36°44.6′E), during spring 2012 and 2013. The wintering individuals were recorded in Tanzania, Mbeya region (8°47.7′S, 34°03.0′E), from January to March 2006, over 6000 km away. The distance between the two breeding localities is 970 km. This ensures a minimal connectivity between the breeding populations and provides different pools of syllables and elements for our analysis, as song repertoire varies with distance in thrush nightingales (Sorjonen 1987).
Populations from the breeding areas were typically found along river banks or woody vegetation patch edges in close neighbouring territories, with a maximum distance of 5 km between sampling sites within a given population. Such males apparently belong to the same sub-population, and thus share a relatively large part of their syllable repertoire, which remains stable and identifiable over several years (AS, unpublished data from Polish and Russian localities). In contrast, the African recordings were made along a 20 km gradient, likely sampling a wider diversity in song repertoire due to the longer distance between individuals, and possibly also their different origin. Although only one population from the wintering ground was analysed in detail due to sample size and recording duration requirements, we also considered several available recordings from Kenya, Malawi, Zambia, Botswana and South Africa (JV unpubl. data, Macaulay Library of Natural Sounds (http://macaulaylibrary.org), Xeno-canto database (http://www.xeno-canto.org)). The use of two well-separated breeding populations, and examination of recordings from other wintering sites, allowed us to reduce the risk that a simple population effect causes the observed differences in syllable consistency and element variability between breeding and wintering grounds. Although we cannot fully rule out such population effects, our approach focussing on syllable consistency within individual repertoires should eliminate the direct effects of geographic variation.
Data collection in the field
Recordings used for this study were made during the morning (0400–1300 h, Russia and Tanzania) or both the morning and evening (0600–0900 and 1900–2000 h, Poland). In the majority of cases, we recorded spontaneously singing individuals. For some recordings from Poland, we used the playback of conspecific songs to elicit vocal behaviour; the potential effect of such playbacks has been controlled for in the analysis (see below).
Our non-invasive recording approach did not involve any marking of individuals. Therefore, particular care was given to the song recording strategies and subsequent selection of recordings to rule out the risk of repeated analysis of the same individual’s song. Only one recording was used per any given recording location, representing confirmed territorial birds in the two breeding populations. At the wintering locality, the birds were usually recorded at greater distances from each other than at the breeding sites, thus minimizing the possibility that one male was recorded twice. Another limitation of studying unmarked birds is the lack of information on the age or pairing status of the subjects. Thus, while individuals of the breeding populations can be considered adults aged 1 year or older, at least some individuals on the wintering ground could have been in their first year (we come back to this issue in the discussion).
Recordings were generally collected from a cautious distance to minimize disturbance of the bird’s natural behaviour. The mean overall duration (mean duration ±SD) of recordings was Poland = 20.3 ± 2.2 min (n = 15), Russia = 15.6 ± 2.2 min (n = 12), Tanzania = 14.0 ± 4.0 min (n = 7). Recording was performed using Sennheiser ME67 (Poland) or ME66 (Russia, Tanzania) directional microphones, connected to Marantz PMD660 (Poland, Russia) or PMD670 (Tanzania) solid-state digital recorders. It was not possible to record data blind because our study involved focal animals in the field.
Song analyses
Song definition
In this study, we define a syllable type as the smallest invariant unit composed of one or a few elements (i.e. the smallest iteration in the spectrogram) that are always repeated in the same sequential order (Fig. 1a). Syllables are usually organized into larger consistent units called “songs”, typically separated from each other by silent pauses (see examples of three different song types in Fig. 1b–g). Songs of thrush nightingales are generally characterized by the presence of two subsequent ending phrases, referred to as “castanet” and “rattling” by Sorjonen (1983). Given the higher variability of syllables at the beginning of the song, even within the same individual, the definition of a song type for this study is based on a combination of the typical two-part ending and the last phrase preceding it (Fig. 1a) traditionally used for descriptions of thrush nightingale song (Sorjonen 1987).
The songs selected for our quantitative analyses had to pass a two-step evaluation: first, they had to exhibit clearly identifiable thrush nightingale species-specific song organization and characteristic ending phrases (see Fig. 1a), as in previous studies from different breeding sites (Sorjonen 1983, 1987; Naguib and Todt 1998; Marova et al. 2015a; Ivanitskii et al. 2016) (Fig. 2). Songs and/or recordings that were lacking the typical syntax and repetition patterns (similar to ‘sub-songs’) were not considered further. Second, those selected in the first step (either typical songs of the species or variants presenting some degrees of variation in their syntax and sequence of elements, similar to ‘plastic songs’) had to successfully pass an automatic detection procedure necessary to ensure the accurate assignment of syllable types (see below).

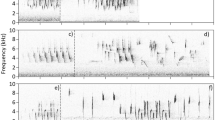
Spectrograms of songs from each population. Note the clear organisation of the thrush nightingale song in a, b Poland and c, d Russia, contrasting with the variability in syllable shapes and repetition patterns visible in various elements of the Tanzanian examples. Song e is representative of the sample successfully used in the cross-correlation analysis, while songs f–h display an increasing level of syllable plasticity, typical of the recordings excluded due to the absence of clear repeatable song patterns
Three syllable categories were defined to cover different aspects of their structural diversity and position in the song, and thus potential associated constraints in vocal production. We aimed to test three syllable types for each individual, one for each category (see Fig. 3, see the full list of tested syllables in Online Resource 1):

Spectrograms (bottom) and oscillograms (top) illustrating examples of selected syllable types used for the cross-correlation analysis in the three different categories (A = disyllable association, B = complex syllables, C = low pitched “castanet”) for each locality
Category A: disyllabic association of two discrete syllables or elements that are always performed one after the other. Those types are usually found among the first stable elements of the recurring sequence defining a song type (see last part of β syllables in red, Fig. 1a).
Category B: this category comprises complex syllables characterized by highly repeated structures, long duration and/or harmonics at either low or high frequency. Similar lower pitched “buzz” structures have been suggested to be costly in the common nightingale (Weiss et al. 2012), due to potential physiological constraints of high-speed production. In thrush nightingales, complex syllables are usually found in the β parts of the songs (see Fig. 1a), except for the Russian population in which they are used as final syllables.
Category C: low pitched “castanet-like” syllables with narrow frequency bandwidth that are highly repeated in the song. They constitute a mandatory part of the characteristic ending of thrush nightingale songs (Sorjonen 1983, 1987).
Populations in Poland and Russia exhibited local syllable sharing between individuals (see Online Resource 1), allowing the selection of comparable syllable types from songs of different individuals for the analysis. In contrast, the absence of such syllable sharing between individuals in Tanzania led to the selection of a larger diversity of syllable types. However, our measure of syllable consistency is not impacted by this difference in the degree of syllable sharing between different populations, as syllables were compared within songs of single individuals.
Syllable consistency
We measured individual syllable consistency by conducting a semi-automatic intra-individual comparison between several occurrences of the same syllable type across different songs. We thus calculated the average spectral cross-correlation (SPCC) of a given syllable rendition in comparison to another rendition of the same syllable type selected from another song of the same type at an analogous serial position (see Fig. 1b–g). For each individual, in the same recording, one syllable rendition was chosen as a reference pattern for each of the three syllable categories, whereby priority was given to renditions with high signal to noise ratios and without masking sounds.
The cross-correlation analysis was performed with Avisoft-SASlab Pro 5.2 (www.avisoft.com, Specht 2007) as follows: first, the syllable repertoire of birds was visually explored to assess the quality of potential candidate syllables. Based on this exploration, appropriate syllable types were preselected according to the criteria of recording quality and whether there was a sufficient number of recurrences at analogous serial positions inside the same song type. The selection of renditions on analogous serial positions was necessary to rule out any potential effects of the serial position (see Fig. 1b–g). Next, we applied the “scan for template spectrogram patterns” function in Avisoft-SASlab for automatic recognition of syllable types. To this end, we used the frequency band cut-off function to compute only those frequency bands covered by the given syllable type, minimizing the effects of potential background noises. Settings were set to exclude any frequency deviation (0 Hz) or masking interval (0 s). To improve this automatic procedure, the recognition of syllables was visually confirmed (false positive = wrong syllable detected) and guided (false negative = matching syllable undetected at the selected threshold) by one human observer (AS). The same observer also carried out the preselection and was not blind as to the population origin of the recordings. Given this semi-automatic approach, the risk of a potential observer-expectancy bias (e.g. Brumm et al. 2017) is expected to be low.
For the recording of each individual that passed the two-step selection procedure, we analysed three syllable types, each representing one of the three categories previously introduced (see examples in Fig. 3), except for one Tanzanian individual for which we could not find an appropriate syllable type for category A and two other Tanzanian individuals with no appropriate syllable types for category B. The mean number (±SD) of renditions of the same syllable type in the same individual compared per category was, for each population: Poland A = 21.1 ± 5.7, B = 11.2 ± 3.8, C = 10.5 ± 2.2; Russia A = 18.8 ± 6.5, B = 12.3 ± 5.5, C = 10.8 ± 4.8; Tanzania A = 9.43 ± 4.1, B = 10.20 ± 2.5, C = 10.67 ± 3.5.
Element variability
We performed an additional analysis to further investigate song characteristics at the element level. Due to the subjectivity of defining limits for an element category when songs presented continuous variability, we used ten different human observers. The observers, experienced with spectrograms of birdsong but not of thrush nightingales and blind to the population origin of the given individuals or tested hypotheses (to avoid any potential bias), were provided with the same set of spectrograms. Each of our three populations was represented by five individuals selected from the same sample as used for the analysis of syllable consistency, and each individual by five songs (resulting in a total of 75 songs). The observers were then instructed to visually assess the number of element types as well as the total number of countable elements present in each song. Given the structural specificity of highly repeated long trills in the typical songs of our species and the technical difficulty in counting the number of elements in the fastest ones, all the broadband clicks repeated more than 3 times in a row (e.g. final trills in Fig. 1) were scored as one “serial element”.
Element variability within each evaluated song was expressed as the ratio of different element types to the total number of elements per song (equivalent to “song versatility” in Järvi 1983; Gil and Slater 2000; Gil et al. 2001). Lastly, a mean index value for an individual bird was calculated based on each observer’s mean score from the five songs of a given individual. We expected element variability to be reflected in the spectrograms by a larger number of different element types sung.
Statistical analysis
Our hypothesis that birds from the wintering grounds sing less stereotyped songs with a lower consistency in syllable performance was tested by a comparison of (1) the syllables’ SPCC for each of our three syllable categories and (2) the element variability index between our three populations with a focus on the contrast between breeding and non-breeding grounds.
We used LMMs (linear mixed-effect models) implemented in R software 3.2.2 (R Core Team 2016). To test for a global difference in the syllable correlation scores between populations, we first fitted full models (nlme package, Pinheiro et al. 2017) with the syllable correlation score as a dependent variable, with the presence or absence of playback used in order to elicit song and population origin as fixed-effects predictor variables and bird’s identity as a random factor. The models were then corrected by either including or not including the potential playback effect based on the output of the previous models for each category separately.
A similar approach was used to test for an overall difference in element variability, with the element variability index as a dependent variable, the population origin as a fixed-effects predictor variable and the bird’s identity as a random factor. A potential influence of playback on the element variability index was not tested by the model, as all the individuals from Poland used for this analysis except one were stimulated, and such an effect would then be confounded into the Polish population category.
To correct for multiple testing, we adjusted the α values of global tests using the sequential Bonferroni procedure. For LMMs that revealed a significant effect of any predictor variables, we used post hoc Tukey’s contrasts for multiple comparisons (multcomp package, Bretz et al. 2010), which provided P values adjusted by the single-step method.
Ethical note
Samples used in this study are based on a non-invasive recording technique from a distance. Birds were not caught, but for few individuals, playback stimulation was performed to trigger an active singing behaviour. This stimulation was kept as short as possible and we are not aware of any consequences for subjects’ breeding or welfare. Authorizations for fieldwork were granted from legal authorities in each different sampling country when needed. For the wintering population, recordings were made within the framework of another research project (Geberzahn et al. 2009, 2010).
Results
While all songs in the recordings from both breeding populations had the typical structure of thrush nightingale song, this was far from being the case in the wintering population: when pooling all recordings across different recording locations, 59% of all Tanzanian recordings (39 out of 66) did not pass the first step of the selection process, as their songs were highly disorganised (i.e. most or all of the typical sub-parts α, β, Ψ1, Ψ2 and ω could not be identified, cf. Fig. 1a) and/or missed silent pauses in between. The recordings available from other known wintering grounds (Kenya, Malawi, Zambia, Botswana and South Africa) showed the same patterns of plastic and disorganized song structure as described above for Tanzanian recordings.
Recordings that did not pass the first step of the selection process could not be considered further as our analysis relied on the ability to assign syllables to syllable types. In contrast, we did not have to exclude any recordings from Poland and Russia (Fig. 2). Of the remaining 27 recordings from Tanzania, only 7 successfully passed the automatic detection procedure (each of them corresponding to a different recording location). Thus, in total, 89% of the Tanzanian recordings (59 out of 66) did not enter the subsequent analysis due to extremely low structural consistency, while all recordings from the breeding sites (15 from Poland and 12 from Russia) passed both steps and were suitable for the quantitative analysis of intra-individual syllable consistency.
Intra-individual syllable consistency
Our measure for syllable consistency (SPCC) showed a significant overall difference between populations for all three categories (Table 1a). In all of them, birds from Tanzania displayed significantly less consistent syllables than both Polish and Russian ones (Table 2, Fig. 4).

Syllable correlation score (SPCC) as a measure of syllable consistency in three populations of thrush nightingales. Shown are three syllable categories: a disyllabic associations (category A), b complex syllables (category B), and c castanet-like syllables (category C). From left to right: Poland (PL), Russia (RU), Tanzania (TZ). Box-and-whisker plots depict the median, interquartile range and non-outlier range (whisker length up to 1.5 times the interquartile range). Numbers underneath the plots represent the total number of observations (n) and the number of subjects (N). Note that the individual identity of birds was entered in the corresponding LMM as “random = (~ 1|ID)” in order to avoid pseudoreplication. Post hoc Tukey tests from the linear models were used to conduct pairwise comparisons of populations for each syllable category (NS = p > 0.05, *p < 0.05, ***p < 0.001)
Significant differences were also found between breeding populations in categories A and B. While birds from the Russian population produced more consistent category A syllables than birds from the Polish population (Table 2, Fig. 4a), the patterns were reversed for the category B syllables, sung less consistently by Russian birds than by Polish ones (Table 2, Fig. 4b).
Element variability
The analysis based on the element variability index obtained by assessments of ten human observers showed a significant difference in element variability between populations (Table 3a). As predicted, songs produced by males wintering in Tanzania showed a higher element variability index than songs from both Polish and Russian populations (Table 3b, Fig. 5). This clearly indicates that syllables in songs from the wintering population were more variable than those from breeding grounds, between which no significant differences in element variability were observed (Table 3b, Fig. 5).

Element variability index in three populations of thrush nightingales assessed at the element level. From left to right: Poland (PL), Russia (RU), Tanzania (TZ). Box-and-whisker plots depict the median, interquartile range and non-outlier range. Numbers underneath the plots represent the total number of songs (n) and the number of individuals (N) tested. Note that the individual identity of birds was entered in the corresponding LMM as “random = (~ 1|ID)” in order to avoid pseudoreplication. Post hoc Tukey tests from the linear model were used to conduct pairwise comparisons of populations for each syllable category (NS = p > 0.05, **p < 0.01)
Discussion
Thrush nightingales recorded on their wintering grounds in Tanzania sang with significantly lower consistency than individuals recorded at two different breeding locations. Our measure of syllable consistency was lower for wintering birds in all three selected syllable categories. Likewise, wintering birds performed songs with more variable elements than did birds from the two breeding populations. Our multiple-population approach allowed us to reduce the potential influence of a simple population effect as the two breeding populations displayed similar patterns, both differing from the wintering one, and additional recordings from other sites in Africa were comparable to those from Tanzania. Below, we consider our findings in light of the song improvement hypothesis proposed by Sorensen et al. (2016). Furthermore, we discuss other potential causes for singing on the wintering ground, based on both field and developmental studies.
Higher vocal plasticity in the wintering population
Songs performed by thrush nightingales on their Tanzanian wintering ground exhibited a strikingly lower syllable consistency and higher element variability than the songs produced by individuals of two different populations from breeding grounds. Concerning the lower syllable consistency quantified by cross-correlation analyses our results are highly conservative, given that more than half of the Tanzanian recordings did not pass the first step of the selection, and about 3/4 of the remaining songs had to be excluded in the second step because of the high variability between different syllable renditions, making assignments to the same syllable type impossible. Therefore, taken as a whole, songs of thrush nightingales were substantially less consistent in Tanzania than estimated by our quantitative analysis.
Such vocal plasticity is usually associated with early sub-song (soft, rambling sounds) and plastic song produced during the sensorimotor phase of the song learning process, in contrast to the more consistent song produced during the breeding season (e.g. Marler and Peters 1982; Hultsch and Todt 2004; Catchpole and Slater 2008). Although we cannot rule out that our Tanzanian sample contains songs of first-year males, by excluding recordings from our analysis that showed structures highly reminiscent of developmental sub-song and plastic song, our data should rather represent birds past their first year of life. Therefore, such vocal plasticity likely reflects older males engaging in a later plastic phase (Margoliash et al. 1991), although studies with marked birds of known age would be necessary to confirm this hypothesis. In any case, we consider that the lower syllable consistency in the wintering population revealed by our analysis supports the song improvement hypothesis, as defined in Sorensen et al. (2016). Higher element variability in the wintering population also likely results from higher vocal plasticity in winter singing. The more variable performance of syllables might have hindered human observers in their assigning of aberrant syllable renditions to the corresponding syllable type. To some extent, however, higher element variability in the wintering populations could be indeed caused by males producing a larger number of different syllable types per song.
While the consistency for all three syllable type categories was always lowest in recordings from Tanzania, the differences between Russian and Polish population were not consistent (Fig. 4). Additionally, the results for the category C (castanet-like syllables, characterized by long repeated sequences and simple structure) suggest an overall higher consistency compared to the categories A and B within each population. This pattern might be explained by a potential learning facilitation for syllables of category C, which are usually exhibited in long repetitive phrases found across the species range (see Sorjonen 1987).
Singing in wintering birds
Our results show that thrush nightingales predominantly produce plastic song when singing on their wintering ground in Tanzania. Similar contrasts have been observed between recordings of thrush nightingales from other known breeding grounds (e.g. Sorjonen 1983; Griessmann and Naguib 2002; Vokurková et al. 2013; Ivanitskii et al. 2016) and from other wintering grounds, including Kenya, Malawi, Zambia, Botswana and South Africa, which display songs with a lack of element consistency comparable to the one described in our analysis (AS personal observation, Online Resource 3). This suggests that plastic song on the wintering grounds is widespread at the species level rather than a rare behaviour. Moreover, our findings are concordant with the results of a study on the common nightingale singing at its wintering grounds in Gambia (Kipper et al. 2016). While their analysis did not assess song consistency quantitatively, the authors noted that the element accuracy and stereotypy of elements seemed lower for most recordings from the non-breeding period. Comparable results were also found in the great reed warbler (Acrocephalus arundinaceus), another species with high song complexity and a similar migration strategy. Great reed warblers were found to use longer songs and switched more often between different syllable types at their winter grounds (Sorensen et al. 2016). In the Palearctic region, Blyth reed warbler (Acrocephalus dumetorum) individuals from the most southern areas of their distribution, most likely migrating individuals, were also found to sing songs with “chaotic syntax and instable phonetic structure” (Marova et al. 2010; Samotskaya et al. 2016). To support and extend these examples, a recent comparative meta-analysis from Sorensen et al. (2016) found a significant link between the occurrence of song in migratory bird species wintering in Africa, and the complexity of their produced song. We may speculate that the occurrence of a singing phase on wintering grounds is widespread, and actually essential, for those species in which the development of complex song extends beyond an early sensory phase of song learning on their breeding grounds (Table 3).
Proximate aspects of plasticity in wintering song
The occurrence of plastic song in wintering birds may be related to the fact that the acquisition of new song characteristics correlates with seasonal changes of the song control regions in the brain over consecutive years, as has been shown in some species (e.g. Nottebohm 1981; Tramontin and Brenowitz 2000). However, whether thrush nightingales reduce their song control nuclei seasonally is currently unknown. In other species that do not exhibit such pronounced seasonal changes, it has been shown that the variability in song structure seems to be actively generated by the song system, as lesions in certain song nuclei can cause a reduction of song variability (e.g. Brainard and Doupe 2000; Kao and Brainard 2006). Song plasticity may thus be functional and allow continuous modification of the song through rehearsal and adjustment to the social environment even in adulthood (e.g. Sakata et al. 2008; Sakata and Vehrencamp 2012; Woolley and Doupe 2008; Tumer and Brainard 2007).
Complementary insights from developmental studies
Song ontogeny has not been directly investigated under controlled laboratory conditions in thrush nightingales. However, in the closely related common nightingale, young males kept in the laboratory spend several months singing plastic song during the winter and early spring (thus whilst their wild conspecifics are on the wintering ground) and crystallize their song at the beginning of the first breeding season (Hultsch and Todt 2004). Furthermore, captive common nightingales go through additional phases of plastic singing before their song crystallizes again in subsequent breeding seasons (Henrike Hultsch, personal communication). This additional phase of plastic singing seems to be a prerequisite for males to integrate song types into their motor program that they have newly acquired during their first season (Todt and Geberzahn 2003), highlighting the potential role of plastic singing as a form of motor exploration in adulthood. Moreover, in common nightingales, hand-reared yearlings not only delayed the use of some early tutored songs to the 2nd year of life but also produced imitations more often in their 2nd year if the same tutored songs were broadcasted again when the individuals were about 1 year old (Geberzahn et al. 2002; Geberzahn and Hultsch 2003). In that regard, singing during the winter season could have a role in activating pre-existent memory as well as increasing and adjusting a bird’s song repertoire to its social environment.
Such adjustments might be particularly important in species that use vocal matching to address each other in vocal territorial disputes, such as common nightingales (Todt and Naguib 2000) or song sparrows (Melospiza melodia, Beecher 2008). In that context, sharing parts of one’s repertoire with territorial neighbours may convey advantages, and such vocal sharing has been reported in many species (see Handley and Nelson 2005 for review). This change in repertoire would thus allow better song matching and conflict regulation between neighbours of the same population over years in highly philopatric species. Thrush nightingales seem to be able to add new elements to their song repertoire over successive breeding seasons, as documented by Sorjonen (1987) and suggested by Griessmann and Naguib (2002). Thus, singing during the wintering period could not only benefit young males developing their singing skills, but also older males adjusting their repertoire to new songs heard during the previous breeding seasons.
Complementary and alternative hypotheses for wintering song
Our results are in accordance with the “market place” hypothesis (Kipper et al. 2016), which proposes that singing outside of the breeding season could be an opportunity for older birds to learn and share new songs with nearby wintering individuals. Wintering grounds would then constitute a song “market place” where males would go through a new sensory phase of song learning, allowing better territorial communication with neighbouring conspecifics over time. From this point of view, the “market place” hypothesis can be regarded as a part of the song improvement hypothesis.
Regardless of the potential delayed benefits for a migrating bird to sing outside of its breeding season, further investigations will be needed to substantiate that wintering song serves as song improvement. Likewise, we cannot rule out other possible functions. The strong territorial behaviour of thrush nightingale males during the breeding season does not seem to be widespread on the wintering grounds, and particular individuals singing highly variable songs could sometimes be observed singing quite close to each other (NG personal observation). Nonetheless, a competition over feeding resources might still occur. In order to determine whether winter singing serves a territorial function in thrush nightingales, future playback studies in Africa would be needed to test whether variable songs recorded from the wintering grounds are potent enough to elicit a territorial response.
Conclusion
This study reveals conspicuous differences in song performance between breeding and wintering grounds in thrush nightingales. Our results improve the understanding of how a culturally transmitted sexually selected trait like birdsong, playing a central role in reproductive success, might be influenced by a complex learning process with functional implications over different seasons. Future studies should aim to track individual birds over their full life cycle in order to provide direct evidence for song improvement. Additionally, further investigations exploring the strategies of non-breeding singing behaviour and its possible functions are needed to achieve a clear understanding of songbirds’ singing behaviour throughout their full life cycle.
References
Alexander CJ (1917) Observations on birds singing in their winter quarters and on migration. Brit Birds 11:98
Bannerman DA (1931) Some evidence of the nightingale, Luscinia megarhyncha megarhyncha, singing in tropical West Africa. Ibis 73:71–74. https://doi.org/10.1111/j.1474-919X.1931.tb01505.x
Beecher MD (2008) Function and mechanisms of song learning in song sparrows. Adv Stud Behav 38:167–225. https://doi.org/10.1016/S0065-3454(08)00004-1
Berglund A, Bisazza A, Pilastro A (1996) Armaments and ornaments: an evolutionary explanation of traits of dual utility. Biol J Linn Soc 58:385–399. https://doi.org/10.1111/j.1095-8312.1996.tb01442.x
Brainard MS, Doupe AJ (2000) Interruption of a basal ganglia–forebrain circuit prevents plasticity of learned vocalizations. Nature 404(6779):762–766. https://doi.org/10.1038/35008083
Brainard MS, Doupe AJ (2002) What songbirds teach us about learning. Nature 417:351–358. https://doi.org/10.1038/417351a
Brenowitz EA, Margoliash D, Nordeen KW (1997) An introduction to birdsong and the avian song system. J Neurobiol 33:495–500. https://doi.org/10.1002/(SICI)1097-4695(19971105)33:5<495::aid-neu1>3.0.CO;2-%23
Bretz F, Hothorn T, Westfall P (2010) Multiple Comparisons Using R. CRC Press, New York
Brumm H, Zollinger SA, Niemelä PT, Sprau P (2017) Measurement artefacts lead to false positives in the study of birdsong in noise. Methods Ecol Evol 8:1617–1625. https://doi.org/10.1111/2041-210X.12766
Catchpole CK, Slater P (2008) Bird song: biological themes and variations, 2nd edn. Cambridge University Press, New York
Cramp S, Perrins CM (1994) The birds of the Western Palearctic. Oxford University Press, Oxford
Darwin C (1871) The descent of man, and selection in relation to sex, vol 2. John Murray, London
Geberzahn N, Hultsch H (2003) Long–time storage of song types in birds: evidence from interactive playbacks. Proc R Soc Lond B 270:1085–1090. https://doi.org/10.1098/rspb.2003.2340
Geberzahn N, Hultsch H, Todt D (2002) Latent song type memories are accessible through auditory stimulation in a hand-reared songbird. Anim Behav 64:783–790. https://doi.org/10.1006/anbe.2002.3099
Geberzahn N, Goymann W, Muck C, ten Cate C (2009) Females alter their song when challenged in a sex-role reversed bird species. Behav Ecol Sociobiol 64:193–204. https://doi.org/10.1007/s00265-009-0836-0
Geberzahn N, Goymann W, ten Cate C (2010) Threat signaling in female song—evidence from playbacks in a sex-role reversed bird species. Behav Ecol 21:1147–1155. https://doi.org/10.1093/beheco/arq122
Gil D, Gahr M (2002) The honesty of bird song: multiple constraints for multiple traits. Trends Ecol Evol 17:133–141. https://doi.org/10.1016/S0169-5347(02)02410-2
Gil D, Slater PJB (2000) Song organisation and singing patterns of the willow warbler, Phylloscopus trochilus. Behaviour 137:759–782. https://doi.org/10.1163/156853900502330
Gil D, Cobb JLS, Slater PJB (2001) Song characteristics are age dependent in the willow warbler, Phylloscopus trochilus. Anim Behav 62:689–694. https://doi.org/10.1006/anbe.2001.1812
Griessmann B, Naguib M (2002) Song sharing in neighboring and non-neighboring thrush nightingales (Luscinia luscinia) and its implications for communication. Ethology 108:377–387. https://doi.org/10.1046/j.1439-0310.2002.00781.x
Handley HG, Nelson DA (2005) Ecological and phylogenetic effects on song sharing in songbirds. Ethology 111:221–238. https://doi.org/10.1111/j.1439-0310.2004.01043.x
Hultsch H, Todt D (2004) Learning to sing. In: Marler PR, Slabbekoorn H (eds) Nature’s Music: The Science of Birdsong. Elsevier Academic Press, London, pp 80–106
Ivanitskii VV, Marova IM, Antipov VA (2016) Sequential organization in the song of thrush nightingale (Luscinia luscinia): clustering and sequential order of the song types. Bioacoustics 26:199–215. https://doi.org/10.1080/09524622.2016.1239132
Järvi T (1983) The evolution of song versatility in the willow warbler Phylloscopus trochilus: a case of evolution by intersexual selection explained by the “female’s choice of the best mate”. Ornis Scand 14:123–128 https://10.2307/3676015
Kao MH, Brainard MS (2006) Lesions of an avian basal ganglia circuit prevent context-dependent changes to song variability. J Neurophysiol 96:1441–1455. https://doi.org/10.1152/jn.01138.2005
Kelsey MG (1989) A comparison of the song and territorial behavior of a long-distance migrant, the marsh warbler Acrocephalus palustris, in Summer and Winter. Ibis 131:403–414. https://doi.org/10.1111/j.1474-919X.1989.tb02788.x
Kipper S, Sellar P, Barlow CR (2016) A comparison of the diurnal song of the common nightingale (Luscinia megarhynchos) between the non-breeding season in the Gambia, West Africa and the breeding season in Europe. J Ornithol 157:1–9. https://doi.org/10.1007/s10336-016-1364-0
Margoliash D, Staicer CA, Inoue SA (1991) Stereotyped and plastic song in adult indigo buntings, Passerina cyanea. Anim Behav 42:367–388. https://doi.org/10.1016/S0003-3472(05)80036-3
Marler P, Peters S (1982) Structural changes in song ontogeny in the swamp sparrow Melospiza georgiana. Auk 3:446–458. https://doi.org/10.1093/auk/99.3.446
Marova IM, Ivanitskii VV, Veprintseva OD (2010) Individual, population, and geographic differentiation in advertising song of the Blyth's reed warbler, Acrocephalus dumetorum (Sylvidae). Biol Bull 37:846–860. https://doi.org/10.1134/S106235901008008X
Marova IM, Ivlyeva AL, Veprintzeva OD, Ivanitskii VV (2015a) A comparative analysis of song differentiation in thrush nightingale (Luscinia luscinia) and common nightingale (Luscinia megarhynchos) within their ranges. Zool Zh 94:701–710. https://doi.org/10.7868/S0044513415060124
Naguib M, Todt D (1998) Recognition of neighbors’ song in a species with large and complex song repertoires: the thrush nightingale. J Avian Biol 29:155–160. https://doi.org/10.2307/3677193
Nottebohm F (1981) A brain for all seasons: cyclical anatomical changes in song control nuclei of the canary brain. Science 214:1368–1370. https://doi.org/10.1126/science.7313697
Nottebohm F, Nottebohm ME, Crane L (1986) Developmental and seasonal changes in canary song and their relation to changes in the anatomy of song-control nuclei. Behav Neural Biol 46:445–471. https://doi.org/10.1016/S0163-1047(86)90485-1
Pearson DJ (1984) The nightingale, sprosser and irania in Kenya. Scopus 8:18–23
Pearson DJ, Backhurst GC (1976) The Southward migration of Palearctic birds over Ngulia, Kenya. Ibis 118:78–105. https://doi.org/10.1111/j.1474-919x.1976.tb02012.x
Pinheiro J, Bates D, DebRoy S, Sarkar D, Core Team R (2017) nlme: Linear and nonlinear mixed effects models. R package version 3:1–131 http://CRAN.R-project.org/prackage=nlme
R Core Team (2016) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria http://www.R-project.org
Reif J, Jiran M, Reifová R, Vokurková J, Dolata PT, Petrusek A, Petrusková T (2015) Interspecific territoriality in two songbird species: potential role of song convergence in male aggressive interactions. Anim Behav 104:131–136. https://doi.org/10.1016/j.anbehav.2015.03.016
Sakata JT, Vehrencamp SL (2012) Integrating perspectives on vocal performance and consistency. J Exp Biol 215:201–209. https://doi.org/10.1242/jeb.056911
Sakata JT, Hampton CM, Brainard MS (2008) Social modulation of sequence and syllable variability in adult birdsong. J Neurophysiol 99:1700–1711. https://doi.org/10.1152/jn.01296.2007
Samotskaya V, Marova IM, Kvartalnov P, Arkhipov VY, Ivanitskii VV (2016) Song in two cryptic species: comparative analysis of large-billed reed warblers Acrocephalus orinus and Blyth’s reed warblers Acrocephalus dumetorum. Bird Study 63:479–489. https://doi.org/10.1080/00063657.2016.1220489
Sorensen MC (2014) Singing in Africa: no evidence for a long supposed function of winter song in a migratory songbird. Behav Ecol 25:909–915. https://doi.org/10.1093/beheco/aru058
Sorensen MC, Jenni-Eiermann S, Spottiswoode CN (2016) Why do migratory birds sing on their tropical wintering grounds? Am Nat 187:E65–E76. https://doi.org/10.1086/684681
Sorjonen J (1983) Transmission of the two most characteristic phrases of the song of the thrush nightingale Luscinia luscinia in different environmental conditions. Ornis Scand 14:278–288. https://doi.org/10.2307/3676320
Sorjonen J (1987) Temporal and spatial differences in traditions and repertoires in the song of the thrush nightingale (Luscinia luscinia). Behaviour 102:196–211. https://doi.org/10.1163/156853986X00126
Souriau A, Kohoutová H, Reif J, Vokurková J, Petrusek A, Reifová R, Petrusková T (2018) Can mixed singing facilitate coexistence of closely related nightingale species? Behav Ecol 29:925–932. https://doi.org/10.1093/beheco/ary053
Souriau A, Geberzahn N, Ivanitskii VV, Marova IM, Vokurková J, Reifová R, Reif J, Petrusková T (2019) Data from: Singing behind the stage: thrush nightingales produce more variable songs on their wintering grounds. Dryad Digital Repository. https://doi.org/10.5061/dryad.tqjq2bvtd
Specht R (2007) Avisoft-SASLab Pro ver. 4.5, 5. Sound analysis and synthesis software. Berlin: Avisoft Bioacoustics. Available at www.avisoft.com
Stach R, Jakobsson S, Kullberg C, Fransson T (2012) Geolocators reveal three consecutive wintering areas in the thrush nightingale. Anim Migr 1:1–7. https://doi.org/10.2478/ami-2012-0001
Thorup K, Tøttrup AP, Willemoes M, Klaassen RHG, Strandberg R, Vega ML, Rahbek C (2017) Resource tracking within and across continents in long-distance bird migrants. Sci Adv 3:e1601360. https://doi.org/10.1126/sciadv.1601360
Todt D, Geberzahn N (2003) Age-dependent effects of song exposure: song organisation sets a boundary between fast and delayed vocal imitation. Anim Behav 65:971–979. https://doi.org/10.1006/anbe.2003.2127
Todt D, Naguib M (2000) Vocal interactions in birds: the use of song as a model in communication. Adv Stud Behav 29:247–296. https://doi.org/10.1016/S0065-3454(08)60107-2
Tramontin AD, Brenowitz EA (2000) Seasonal plasticity in the adult brain. Trends Neurosci 23:251–258. https://doi.org/10.1016/S0166-2236(00)01558-7
Tumer EC, Brainard MS (2007) Performance variability enables adaptive plasticity of ‘crystallized’ adult birdsong. Nature 450:1240–1244. https://doi.org/10.1038/nature06390
Vokurková J, Petrusková T, Reifová R, Kozman A, Mořkovský L, Kipper S, Weiss M, Reif J, Dolata PT, Petrusek A (2013) The causes and evolutionary consequences of mixed singing in two hybridizing songbird species (Luscinia spp.). PLoS ONE 8:e60172. https://doi.org/10.1371/journal.pone.0060172
Weiss M, Kiefer S, Kipper S (2012) Buzzwords in females’ ears? The use of buzz songs in the communication of nightingales (Luscinia megarhynchos). PLoS ONE 7:e45057. https://doi.org/10.1371/journal.pone.0045057
Woolley SC, Doupe AJ (2008) Social context–induced song variation affects female behavior and gene expression. PLoS Biol 6:e62. https://doi.org/10.1371/journal.pbio.0060062
Zeigler P, Marler P (2008) The Neuroscience of Birdsong. Cambridge University Press, New York
Acknowledgments
We want to thank all the colleagues involved in the assessment of element variability for their precious help: Ondřej Belfín, Paolo Biella, Sharina van Boheemen, Hana Kahounová, Jan Mertens, Alexandra Průchová, Lucie Jiránková and especially Adam Petrusek for his overall support and help with the manuscript. NG would like to thank Wolfgang Goymann, Sonja and Andreas Kümmerle and Liz and Neil Baker for logistic support as well as Mackubi Joseph for help with fieldwork in Tanzania. VI and IM are grateful to Vladislav Antipov for the recording of nightingales in Russia. We also thank David Hardekopf for language corrections. Additional thanks go to Tony Archer, James Bradley, Josh Engel, Niall Perrins and Derek Solomon for kindly consenting to the use of their recordings. Lastly, we are grateful to two anonymous reviewers and Jeff Podos for their valuable comments on a previous version of the manuscript.
Funding
This work was supported by the Association for the Studies of Animal Behaviour (ASAB) to NG; the Deutsche Forschungsgemeinschaft (grant numbers GE 1253/3-1 and GE 1253/3-2 to NG); the Russian Fund for Basic Research (grant number 16-04-01721 to VI) and the Russian Science Foundation (grant number 14-50-00029 to VI).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical approval
All applicable international, national, and institutional guidelines for research on animals were followed. Samples used in this study are based on a non-invasive recording technique from a distance. Birds were not caught, but for few individuals, playback stimulation was performed to trigger an active singing behaviour. This stimulation was kept as short as possible and we are not aware of any consequences for subjects’ breeding or welfare. Passive recording did not require supervision from institutional or regulatory bodies, but authorizations were granted from legal authorities in each different sampling country when needed. In the wintering population, recordings were made within the framework of another research project (for more details see Geberzahn et al. 2009, 2010).
Conflict of interest
The authors declare that they have no conflict of interest.
Data accessibility
Analyses reported in this article can be reproduced using the data provided in Souriau et al. 2019 (data Tables with recordings information and spectrograms prints used for the human-based assessment of element variability).
Additional information
Communicated by J. Podos
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Online resource 1
Spectrograms of syllable types used in the cross-correlation analysis for each population and each category. The letters above the spectrograms indicates the population (PL: Poland, RU: Russia, TA: Tanzania), the syllable category and the identity code of the subject (PDF 1118 kb)
Online resource 2
Contrast in syllable consistency scores (SPCC) between categories for each population. Table a shows the overall contrast in the element variability index between all three categories (A = disyllabic associations, B = complex syllables, C: castanet-like syllables). Table b shows all pairwise comparisons from the modelling of the effect of syllable category on the syllable consistency score, for each population (PDF 185 kb)
Online resource 3
Spectrograms of thrush nightingale song bouts from several different wintering sites in Africa. All wintering populations showed various degrees of song plasticity, reflecting similar observations in our focus population in Tanzania. Recording references are as following (www.xeno-canto.org): (a) Kenya (XC396432), (b) Malawi (XC82082) lacking typical song organization of thrush nightingale, (c) Zambia (XC41337) and (d) Botswana (XC46020) plastic songs with high syllable and element variability, (e) and (f) South Africa (XC400388 and XC184009) song resembling the typical species songs with some level of plasticity (PDF 1960 kb)
Rights and permissions
About this article

Cite this article
Souriau, A., Geberzahn, N., Ivanitskii, V.V. et al. Singing behind the stage: thrush nightingales produce more variable songs on their wintering grounds. Behav Ecol Sociobiol 73, 150 (2019). https://doi.org/10.1007/s00265-019-2765-x
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-019-2765-x