
Abstract
The pace-of-life syndrome (POLS) hypothesis posits that life-history characteristics, among individual differences in behavior, and physiological traits have coevolved in response to environmental conditions. This hypothesis has generated much research interest because it provides testable predictions concerning the association between the slow-fast life-history continuum and behavioral and physiological traits. Although humans are among the most well-studied species and similar concepts exist in the human literature, the POLS hypothesis has not yet been directly applied to humans. Therefore, we aimed to (i) test predicted relationships between life history, physiology, and behavior in a human population and (ii) better integrate the POLS hypothesis with other similar concepts. Using data of a representative sample of German adolescents, we extracted maturation status for girls (menarche, n = 791) and boys (voice break, n = 486), and a set of health-related risk-taking behaviors and cardiovascular parameters. Maturation status and health-related risk behavior as well as maturation status and cardiovascular physiology covaried in boys and girls. Fast maturing boys and girls had higher blood pressure and expressed more risk-taking behavior than same-aged slow maturing boys and girls, supporting general predictions of the POLS hypothesis. Only some physiological and behavioral traits were positively correlated, suggesting that behavioral and physiological traits might mediate life-history trade-offs differently. Moreover, some aspects of POLS were sex-specific. Overall, the POLS hypothesis shares many similarities with other conceptual frameworks from the human literature and these concepts should be united more thoroughly to stimulate the study of POLS in humans and other animals.
Significance statement
The pace-of-life syndrome (POLS) hypothesis suggests that life history, behavioral and physiological traits have coevolved in response to environmental conditions. Here, we tested this link in a representative sample of German adolescents, using data from a large health survey (the KIGGs study) containing information on individual age and state of maturity for girls and boys, and a set of health-related risk-taking behaviors and cardiovascular parameters. We found that fast maturing girls and boys had overall higher blood pressure and expressed more risk-taking behavior than same-aged slow maturing girls and boys. Only some behavioral and physiological traits were positively correlated, suggesting that behavioral and physiological traits might mediate life-history trade-offs differently and not necessarily form a syndrome. Our results demonstrate a general link between life history, physiological and behavioral traits in humans, while simultaneously highlighting a more complex and rich set of relationships, since not all relationships followed predictions by the POLS hypothesis.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The pace-of-life syndrome (POLS) hypothesis seeks to characterize phenotypic attributes that mediate the trade-off between current and future reproductive success (Réale et al. 2010; Careau and Garland 2012). As such, it extends one of the core trade-offs in life-history theory (Stearns 1989), to include behavioral and physiological attributes, whereby the assumption is that these attributes have coevolved resulting in a suite of co-varying, functionally relevant traits, depending on the local ecology (Réale et al. 2010). Following the introduction of the POLS hypothesis in animal and behavioral ecology, there has been a rapid accumulation of studies testing the original POLS predictions in a wide range of animal species with conflicting results (Montiglio et al. 2018, topical collection on Pace-of-life syndromes; Royauté et al. 2018, topical collection on Pace-of-life syndromes). Whereas there is an abundance of studies investigating POLS in animals, the POLS hypothesis has not yet been directly applied to the domain of human ecology although many similar concepts have been developed (for more details, see below).
Humans show a relatively slow life history, in comparison to most other mammal species, with late maturation, long life span, senescence, but short inter-birth intervals in comparison to same sized primates (Smith 1992; Hill and Kaplan 1999; Kaplan et al. 2000). Substantial variation in several key human life-history traits (e.g., growth velocity, age of menarche, onset of puberty) has been reported, both on the within and the between population level (Eveleth and Tanner 1990; Worthman 1999; Walker et al. 2006; Pettay et al. 2007). The main focus of the POLS hypothesis is on the trade-off between current and future reproductive success; however, specific characteristics of humans (e.g., longevity, extended parental care, and intergenerational effects) create several more important trade-offs (see “Discussion”).
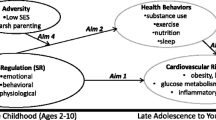
In the human ecology literature, several conceptual frameworks have been introduced, which bear close resemblance to the POLS hypothesis. One of the earliest theories, integrating life-history traits, behavior, and physiology in humans, was proposed by Rushton (Rushton 1985; Figueredo et al. 2013): In his differential K theory, Rushton categorizes humans along a continuous dimension K, with higher K values representing individuals coming from small families with higher parental investments and longer inter-birth intervals and these individuals are predicted to also mature slower and show higher behavioral constraint and altruism (see also Ellis 1988; Figueredo et al. 2004). As such, Rushton’s theory is an application of the “r/K selection” theory introduced by MacArthur (MacArthur 1962; MacArthur and Wilson 2015). Belsky et al. (1991) developed an influential evolutionary-based theory of social organization and lifespan interpersonal development. Their theory postulates that a range of early social contexts, rearing, and physiological and behavioral orientations, predict markers of reproductive strategy (e.g., pubertal timing, sexual activity, and pair bonding) in an adaptive manner. More specifically, the experiences during early life (5–7 years) are predicted to give individuals an estimate of the availability and predictability of resources and trustworthiness of their social environment in the future. Individuals experiencing scarce and/or unpredictable resources and untrustworthy others are expected to advance their sexual maturation and invest less in social bonds. Building on these insights, Ellis et al. (2009) introduced a developmental theory of inter-individual variation in humans proposing a cluster of correlated life-history traits lying on a slow-to-fast continuum, crucially mediated by environmental harshness and unpredictability. Other frameworks have also integrated physiological attributes [e.g., stress response (Del Giudice et al. 2011)] and/or behavioral attributes [e.g., extraversion (Nettle 2005), risk-taking (Ellis et al. 2012)] with life-history traits (see also Belsky 2012). Moreover, numerous empirical studies have investigated potential links between behavior and/or psychological attributes (e.g., Figueredo et al. 2004; Figueredo and Rushton 2009; Jokela et al. 2009, 2011; Ellis et al. 2011; Griskevicius et al. 2011) and life-history traits (see also discussion), though few empirical studies integrate behavior, physiology, and life-history traits within one study. Thus, many concepts similar to POLS are in the extant human literature but they are either specified for humans with their unique life history, differing from most other animals, and their specific socio-cultural environmental conditions and/or use psychosocial instead of biological measures for POL (e.g. Figueredo et al. 2004, 2005, 2007; Figueredo and Rushton 2009). In our opinion, the POLS hypothesis goes beyond these other frameworks by conceptually explaining the evolution of within-species variation in life-history strategies along the current versus future reproduction trade-off by the mediating function of behavioral and physiological traits (Dammhahn et al. 2018, topical collection on Pace-of-life syndromes). Here, we aim to test specific predictions of the POLS hypothesis with an exemplary data set for humans and to contribute to joining these overall similar conceptual frameworks.
Specifically, we investigated several of the key predictions of the POLS hypothesis (Réale et al. 2010) within a human population. To this end, we used data from the German Health Interview and Examination Survey for Children and Adolescents (KiGGS). The KiGGS data set contains information on maturation status, risk-taking behavior, and physiology from a representative sample of German adolescents (Hölling et al. 2007, 2012; Kurth et al. 2008). As an indication of pace-of-life, we used maturation status in same-aged children questioned at an age when half of them were mature and studied how presence and absence of the life-history marker covaried with behavioral and physiological traits. Age of sexual maturation has been used as an indicative measure of pace-of-life in many inter- and intra-specific studies in vertebrates (Stearns 1992; Oli 2004). Menarcheal age is commonly used as a biological marker of female development (Susanne and Bodzsar 1998) and occurs usually between Tanners pubertal stages of breast development B3 and B4 (Tanner 1962; Marshall and Tanner 1969; Harries et al. 1997). Biological characteristics of male sexual maturity are pubic hair development, a characteristic with different stages and high variability over the whole pubertal time, and voice break (a deepening of the voice); a characteristic rarely used as a clinical or biological marker. In contrast to pubic hair development voice break occurs around one year after spermarche (first motile spermatozoa) (Bogin 1999) and between Tanners pubertal stages of genital development G3 and G4 (Harries et al. 1997). Voice break is therefore better comparable (in absence of information of age of spermarche) with menarcheal age in girls and can, as such, be seen as a proxy for the pace-of-life in boys (e.g., Juul et al. 2007).
As behavioral traits, we focused on risk-taking behavior as part of the possible syndromes provided by the POLS hypothesis and because of the characterization of the adolescent transition through increased rates of risky and reckless behavior with potentially severe consequences including disability and death due to, for example, violence, suicide, accident, eating disorders, and drug and alcohol use (Steinberg 2008; National Research Council 2011; Ellis et al. 2012). As physiological traits, we focused due to limitations of the KiGGS data set on a set of cardiovascular parameters. We used these parameters as a proxy for the sympathetic/parasympathetic nervous system reactivity, which has been suggested to align with pace-of-life (Réale et al. 2010). For humans, the Adaptive Calibration Model (ACM; Del Giudice et al. 2011, Ellis et al. 2017) would allow to make more nuanced predictions regarding physiological regulation of life histories, which cannot be tested, however, with our limited data set (see also “Discussion”).
Based on the POLS hypothesis (Réale et al. 2010; Dammhahn et al. 2018, topical collection on Pace-of-life syndromes), we expect covariation between life history, risk-taking, and cardiovascular physiology. Specifically, based on the verbal framework presented in Fig. 1 of Réale et al. (2010), we expect that individuals who mature faster show (i) increased risk-taking behavior and (ii) score higher on cardiovascular parameters, compared to individuals who mature slower. Moreover, if risk-taking behaviors and physiology mediate the trade-off between current and future reproduction similarly, we expect (iii) a positive correlation between risk-taking behaviors and cardiovascular parameters.

Standardized mean difference (Hedges’ g) and 95% CI between same-aged fast maturing and slow maturing girls and boys. Positive values indicate that the fast life-history group showed higher values for this variable (either higher values of cardiovascular parameters, or higher risk-taking behavior). Negative values indicate that the slow life history group showed higher values for this variable. Overall effect strengths within trait domains (gray diamonds) and over all traits (black diamonds) were calculated using random-effects meta-analysis models
Methods
Study population
The KiGGS study is a health survey of 17,641 children and adolescents of both sexes, aged between 0 and 18 years, and was performed between 2003 and 2006 in Germany (Hölling et al. 2007, 2012; Kamtsiuris et al. 2007; Kurth et al. 2008; Rosario et al. 2010). This cross-sectional study gathered representative data through medical health examinations and questionnaire-based assessments and it uniquely encompasses data on physiology, risk-taking behavior, and life history. All information for one person was collected within 1 day (Hölling et al. 2007). Data collection (detailed explained in Hölling et al. 2007, 2012, Kamtsiuris et al. 2007 and Kurth et al. 2008) and data analysis by the authors were performed independently of each other which minimize observer bias. For more information about study design and quality management within the KiGGS study, please see the literature mentioned above.
Life history
The KiGGS study provides data on the exact age of the adolescents, as well as information about the occurrence of menstruation of girls and voice break of boys, both quantified as yes or no at the time of questioning. Because the exact timing of the onset of either maturation events was not available, we selected one homogeneous age group for each sex and compared individuals who did mature with those that did not mature within this age group. We decided against the opposite approach: grouping children of the same state but different ages (young mature versus old mature) since we assume that the social environment and behavior of contemporaries is strongly associated and behavior, especially to risk-taking behaviors, may overlay differences due to differential POL. In addition, we first removed data of children with migration background from the data set. Because the developmental tempo (age of maturation) between different human populations can differ because of various genetic as well as socio-economic conditions (Susanne and Bodzsar 1998). We further removed age groups below 11 years because they were not included in the questionnaire-based assessment to assess risk taking (see below).
In the remaining data set (2804 girls and 2205 boys between 11 and 18 years), we searched for the age with a 50% probability of having reached maturity, using logistic regression models with binomial residual distribution and a “logit” link function. Girls reached the 50% probability of having menarche with 12.89 years (± standard error (SE) = 0.04). Boys reached the 50% probability of voice break experience at an age of 14.35 years (± SE = 0.05 years). Next, we formed homogeneous age groups by including all individuals that were up to 1 year younger or older than the age at which 50% of all individuals reached maturity. This procedure allowed us to compare slower (absence of maturity sign) with faster (presence of maturity sign) pace-of-life groups while controlling for age. Our final sample comprised 791 girls (378 without menarche (slow maturing) and 413 with menarche (fast maturing), age range 11.89–13.89) and 486 boys (226 without voice break (slow maturing) and 260 with voice break (fast maturing), age range 13.35–15.35). By categorizing girls and boys into these age-homogenous groups depending on their maturational status, we largely controlled for age in all further analysis.
Risk-taking behavior
For quantifying different categories of adolescents’ risk-taking behavior, we used the behavioral categories described by the Youth Risk Behavior Surveillance System (YRBSS) of the Centers for Disease Control and Prevention (CDC) of the United States (Brener et al. 2002; Kann et al. 2016). This program distinguishes six broad categories of health-risk behaviors: (a) tobacco use, (b) alcohol and other drug use, (c) inadequate physical activity, (d) unhealthy dietary behaviors, (e) behaviors related to unintentional injuries and violence, and (f) risky sexual behaviors.
The parental and child questionnaire of the KiGGS study contains detailed information on a range of health-risk behaviors, including tobacco use, alcohol and drug use, diet, violence, and quality of life of German adolescents of both sexes, aged between 11.00 and 18.00 years (Hölling et al. 2007, 2012; Kamtsiuris et al. 2007; Kurth et al. 2008; Rosario et al. 2010). To the best of our knowledge, only Höpker et al. (2014) used the KiGGS data set to analyze adolescent risk behavior. Since no validated measures of behaviors were available yet, we screened the KiGGS questionnaires and identified questionnaire items comprising information falling into the six categories of health-risk behaviors of the YRBSS. In total, we identified 85 (continuous or categorical) such questionnaire items distributed as follows: (a) tobacco use (n = 5), (b) alcohol and other drug use (n = 13), (c) inadequate physical activity (n = 9), (d) unhealthy dietary behaviors (n = 6), and (e) behaviors related to unintentional injuries and violence (n = 52) (more details in supplement Table S1). There were no questions related to risky sexual behaviors in the survey.
To reduce the amount of heterogeneous questionnaire items to a smaller set of interpretable components, we used a categorical principal component analyses (catPCA) (Linting and van der Kooij 2012). We used this data reduction step instead of directly applying confirmatory analyses because many questionnaire items were ordinal or nominal with very heterogeneous distributions, which hampered fitting complex models directly (for further confirmatory steps of the analysis see below). The catPCA is similar to a linear principal component analysis but allows modeling non-linear relationships and including numeric, ordinal, and nominal variables. In a first step, we sorted all risk behavior-related questionnaire items according to the degree of risk, whereby a small value encoded low risk and vice versa, except for inadequate physical activity behaviors. For using catPCA, we followed the guidelines of Linting and van der Kooij (2012), which recommend an initial analysis to evaluate analysis level, variable selection, and the number of components. We ran a two-dimensional initial analysis treating all variables as spline nominal level, except dichotomous variables which were set as nominal. Based on transformation plots, we chose the appropriate analysis level of each variable; for ordinal and nominal spline level, we used the default settings of splines with two interior knots and two degrees of spline. Questionnaire items were considered meaningful variables if they accounted for more than 10% of the total variance (VAF > 0.1). To determine the final number of dimensions (components), we used the following recommended criteria: (i) elbow criterion in a scree plot (Fabrigar et al. 1999), (ii) Eigenvalue larger than one (Kaiser Criterion), and (iii) interpretability of the components. We also compared the results of different dimensionalities for evaluation of the chosen number of components (Linting et al. 2007; Linting and van der Kooij 2012). Because catPCA requires only integer variables, we chose [following Linting and van der Kooij 2012] appropriate discretizing procedures. Ranking procedure was used for continuous variables which were treated as ordinal (spline) or nominal (spline) in the analysis level. Variables, treated as numeric, were discretized with a multiplying procedure. In case of non-responding participants for particular questionnaire items, we followed the missing value strategy of excluding those ones for each variable separately. Because rotation methods in catPCA are not available, we re-analyzed all transformed variables in a linear PCA to perform oblique (oblimin with δ = 0) rotation. Pattern matrix and structure matrix (results of oblique rotation) were only used for increasing interpretability. For each combination of sex and risk-taking category, we conducted a separate catPCA using the outcomes to define health-related risk behavior components (Table 1).
Applying this procedure to the completed 85 questionnaire items of 2802 girls and 2896 boys aged between 11 and 18 years, we extracted 11 risk-taking components for girls and 10 for boys (Table 1, supplement Table S2 for girls and Table S3 for boys). Of those, we excluded “drug use behaviors” for further analyses due to an unrepresentative amount of positive drug use statements (within our subsample less than 10% of the participants reported to have used drugs at least once). Further, we excluded one component representing the psychosocial state of an individual (e.g., fear, peer pressure) because it fell outside of the scope of the six categories of health-risk behaviors of the YRBSS. Finally, we excluded health-risk behaviors that were not actively performed (e.g., exposure to smoking “passive tobacco use”).
Our final data set for health-related risk behaviors thus encompassed only actively performed behavior and included the following variables: active tobacco use, alcohol use, inadequate physical activity, media consumption, unhealthy dietary behaviors, physical violence, and two components of neglecting protection (i.e. not using a helmet when riding a bike) (details in supplement Tables S2 and S3). Using Pearson correlations of components with original questionnaire items, we checked for interpretability of components and if necessary recoded variables so that each component increased in value with increasing risk-taking. This was achieved by multiplying the following risk-taking components by − 1: inadequate physical activity and media consumption in both sexes and neglecting protection in boys only (see supplement Tables S2 and S3). For example, a high value of active tobacco use indicates that the individual smokes a lot and started to use tobacco at an early age; a high value in physical violence behavior signaled a higher willingness to be physically violent.
Physiological parameters
The KiGGS data set comprises five cardiovascular parameters: systolic and diastolic blood pressure, mean arterial pressure, pulse pressure, and heart rate. Full details on the methods employed to measure these parameters are provided in Neuhauser and Thamm (2007) and Neuhauser et al. (2011). All parameters, except heart rate, were measured twice at the same day; we used the mean value of these two measurements.
Statistical analyses
To test whether fast and slow maturing boys and girls differ in a set of physiological and risk-taking behavioral traits, we calculated the standardized mean difference Hedges’ g to compare both life-history groups for each trait for girls and boys, respectively. A positive value of Hedges’ g implies a higher mean estimate for the fast life-history group than for the slow life-history group, and vice versa. Confidence intervals (95% CIs) of the standardized mean difference are based on the sampling variance. We calculated (for visual representations) overall effects using random-effects meta-analyses models, running separate analyses including (i) all physiological parameters, (ii) all health-related risk-taking behaviors, and (iii) all variables.
To test whether physiological and behavioral traits were correlated at the phenotypic level, we used Pearson correlations for each pair of physiological and behavioral traits within the selected subsample of 791 girls and 486 boys. We considered correlations as significant, if 95% CIs excluded 0; all 95% CIs are given in square brackets.
To examine the relationships between physiological, behavioral, and life-history traits under the POLS hypothesis, we used confirmatory factor analysis (CFA). CFA is a structural equation modeling technique that allows us to measure latent variables (Hoyle 1995; Kline 2011) and to test our hypothesized structure of POLS. For our analysis, we followed the guidelines for constructing structural equation models by Ullman (2006) and Fan et al. (2016). We constructed a model including three latent variables representing behavior, physiology, and life history in accordance with POLS theory. In our model, the latent behavior variable represents health-related risk behavior. For the CFA, we limited the set of behavioral indicator variables to those which most strongly represent health-related risk behavior; these four indicator variables were alcohol use, active tobacco use, inadequate physical activity, and physical violence behavior. Due to our data set structure (only one variable expressing life history), highly correlated cardiovascular characteristics (systolic blood pressure and diastolic blood pressure (r > 0.67, p < 0.001 in girls; r > 0.65, p < 0.001 in boys)) and the once measured variable heart rate the latent variables expressing life history and physiology were formed by only one indicator variable each. Physiology was expressed by systolic blood pressure and life history by the occurrence of maturation (girls menarche and boys voice break, for more explanation please see above) (see also Fig. 3).
To perform a CFA, we prepared and re-structured the data as follows: as indicator variables expressing health-related risk behavior, we used the factor scores generated by catPCA of alcohol use, active tobacco use, physical violence behavior, and inadequate physical activity (for more details please see above) which were log-transformed, except for active tobacco use. Active tobacco use was used in the CFA model as a binary variable (active smoking: yes or no). Furthermore, maturational status was used as a binary indicator variable and systolic blood pressure used as a z-scored continuous variable. We checked our data set for multivariate normality, one important assumption when modeling with maximum likelihood procedures (Fan et al. 2016), with Mardia’s Multivariate Normality Test (Mardia 1970; Korkmaz et al. 2014). Due to the non-multivariate normality, neither in girls (pskewness < 0.001 and pkurtosis = 0.79) nor in boys (pskewness < 0.001 and pkurtosis < 0.001), and the complex structure of the data set (a mixture of continuous and binary variables) the appropriated estimation method “WLSMV” was used to get robust standard errors (SE) and adjusted test statistics (Rosseel 2012). In accordance with SEM guidelines (Ullman 2006; Fan et al. 2016), we used the Χ2 test and the recommended indices RMSEA (root mean square error of approximation), CFI (comparative fit index), and SRMR (standardized root mean square residual) for assessing model fit. We conducted separate CFAs for boys and girls.
All analyses were calculated using R 3.1.2 (R Development Core Team 2014). For the calculation of Hedges’ g, we used the “Metafor” package (version 1.9–7 + Viechtbauer (2010)). CFA models were built using the “lavaan” package (version 0.5–20 + Rosseel (2012)), while the analysis of multivariate normality was done within the “MVN” package (0.1–2 + Korkmaz et al. (2014)). Descriptive statistics (mean and standard deviation (SD)) of cardiovascular characteristics and risk-taking components for girls and boys, catPCA modeling and correlation analyses were done with IBM SPSS Statistics for Windows (version 22). CIs were estimated with bootstrapping using percentile-method with 1000 bootstrap samples within SPSS. For all traits in boys and girls, sample sizes differed due to missing information within questionnaires (for full information on sample sizes for each analyses see supplement Tables S4 and S5).
Results
Mean age of 791 girls was 12.89 years (SD = 0.59); girls of the fast life-history group (n = 413) had a mean age of 13.09 years (SD = 0.55), whereas girls of the slow life-history group (n = 378) showed a mean age of 12.68 years (SD = 0.56). Boys (n = 486) were on average 14.38 years old (SD = 0.61), whereby “fast” boys (n = 260) had a mean age of 14.67 years (SD = 0.54) and “slow” boys (n = 226) of 14.04 years (SD = 0.51). Further descriptive statistics for all physiological parameters and all risk behavioral components separated by sex and life-history group are presented in supplement Table S4.
Comparison between fast and slow life-history groups
Overall, fast maturing girls and boys had higher cardiovascular parameter values than slow maturing girls and boys (Fig. 1, Table S4). Particularly, blood pressure was higher in fast than in slow life-history groups; except for diastolic blood pressure, which only tended to differ between fast and slow maturing girls. In contrast, fast maturing individuals of both sexes had lower heart rates.
Overall, boys and girls who matured faster tended to express higher scores of health-related risk-taking behaviors than those that matured slower (Fig. 1, Table S4). In girls, fast maturing individuals showed higher levels of substance use (active tobacco use and alcohol use), inadequate physical activity, unhealthy dietary behaviors, physical violence, and neglecting protection 2. Girls belonging to the slow life-history group showed higher media consumption behavior. There were no differences in neglecting protection between both life-history groups in girls (Fig. 1, Table S4). Boys with a fast life history showed more risk-taking concerning active tobacco use, alcohol use, media consumption, and neglecting protection 2 and a slight tendency to have more inadequate physical activity behaviors. In contrast, “slow” boys neglect protection to a higher degree, and indicated more unhealthy dietary behaviors and tended to show more physical violence (Fig. 1, Table S4).
Correlations between health-related risk-taking behaviors and physiology
Overall, pair-wise correlations between physiological and behavioral traits revealed a mixed pattern (Fig. 2). For girls, three components of risk-taking were positively correlated phenotypically with most cardiovascular parameters, namely inadequate physical activity (all p < 0.05, except pulse pressure p > 0.1), media consumption (all p < 0.05, except mean arterial pressure p < 0.1, systolic blood pressure and pulse pressure p > 0.1), and unhealthy dietary behavior (all p < 0.05, except diastolic blood pressure p > 0.1 and heart rate p > 0.1). In addition, physical violence was positively correlated with pulse pressure (p < 0.05). For alcohol use, we found no significant correlations with cardiovascular parameters. Neglecting protection was negatively correlated with most cardiovascular parameters (all p < 0.05, except pulse pressure p > 0.1). For active tobacco use, we found negative correlations with diastolic blood pressure and mean arterial pressure (p < 0.05). Physical violence was also negatively correlated with diastolic blood pressure (p < 0.05). Neglecting protection 2 was negatively correlated with heart rate (p < 0.05) (Fig. 2, Table S5).

Correlation coefficients and 95% CI of all possible pair-wise combinations of physiological parameters (a–e) and health-related risk-taking behaviors for girls (aged 11.89–13.89 years) and boys (aged 13.35–15.35 years)
In boys, inadequate physical activity was positively correlated phenotypically with most cardiovascular parameters (all p < 0.05, except pulse pressure p > 0.1). Four components of risk-taking were negatively correlated with at least one cardiovascular characteristic, namely physical violence (all p < 0.05, except pulse pressure p > 0.1), alcohol use (heart rate p < 0.05), neglecting protection (all p < 0.05, except systolic blood pressure p < 0.1 and pulse pressure p > 0.1), and media consumption (heart rate p < 0.05). For the remaining risk-taking behaviors (neglecting protection 2, active tobacco use and unhealthy dietary behaviors), no significant correlations were found (Fig. 2, Table S5).
Confirmatory factor analysis for girls and boys
The fit of the CFA model of the girls sample (n = 524) revealed a slight deviation of the implicit covariance matrix of the indicators of our chosen model structure from the real one (Χ2 = 16.58, df = 8, p = 0.04). Further CFA model fit indices for the girls’ sample were quite good; RMSEA = 0.045 (p = 0.553; 90% CI = 0.012–0.076) and SRMR = 0.03. Only the CFI of 0.93 indicated a poor fit. Overall, model fit for the boys sample (n = 328) was poor (Χ2 = 32.55, df = 8, p < 0.001; RMSEA = 0.097, p = 0.012; 90% CI = 0.06–0.133; CFI = 0.799, SRMR = 0.056). Therefore, all results of the CFA, particularly for the boys’ sample, have to be interpreted with caution.
The latent factor solutions for our CFA models for girls and boys are shown in Fig. 3 and Table 2. For girls, standardized structure coefficients (SSC) for indicator variables of health-related risk behavior ranged between 0.28 (for active tobacco use) and 0.43 (for alcohol use). The latent variable life history was correlated with health-related risk behavior (cov = 0.44) and to a lower degree with physiology (cov = 0.15). In girls, physiology and health-related risk behavior were not correlated (Fig. 3, Table 2). In boys, alcohol use (SSC = 0.77), active tobacco use (SSC = 0.60), and physical violence behavior (SSC = 0.37) support the latent variable health-related risk behavior. Only physical activity seemed to have no influence. In boys, life history was correlated with health-related risk behavior (cov = 0.30) and with physiology (cov = 0.23). We found no support for the relationship between physiology and health-related risk behavior (Fig. 3, Table 2).

Structure of confirmatory factor analysis (CFA) and strength of associations (based on standardized structure coefficients) in the model of life history (maturity), health-related risk-taking behaviors and physiology in two separate models, one for girls (left) and one for boys (right). Indicator variables (observed variables) were illustrated with rectangles and latent variables with ellipses. Single arrowheads express a direct effect of the latent variable on the indicator variables; two-headed arrows indicate covariance between latent variables. Solid arrows illustrate significant relationships, dashed arrows non-significant relationships
Discussion
The POLS hypothesis posits that phenotypic attributes mediate the trade-off between current and future reproduction (Réale et al. 2010; Dammhahn et al. 2018, topical collection on Pace-of-life syndromes). This hypothesis has generated much research interest in animal and behavioral ecology (Montiglio et al. 2018, topical collection on Pace-of-life syndromes; Royauté et al. 2018, topical collection on Pace-of-life syndromes) because it provides testable predictions concerning the association between the slow-fast life-history continuum and behavioral and physiological traits. Here, we tested several of these predictions using data of a contemporary human population. We found covariation between maturation status and health-related risk behavior as well as between maturation status and cardiovascular physiology for both boys and girls, supporting a general POLS. Further, we found that fast maturing boys and girls had overall higher blood pressure and tended to express more risk-taking behavior than same-aged slow maturing boys and girls. Thus, general predictions of the POLS hypothesis are partly supported for a human population. Only some of the single physiological and behavioral traits were positively correlated (Fig. 2, Table S5) and the CFA did not support covariation between cardiovascular physiology and health-related risk behavior (Fig. 3, Table 2). Hence, behavioral and physiological traits might mediate life-history trade-offs differently and not necessarily form a syndrome. Moreover, we found commonalities but also interesting differences between the sexes, suggesting that some aspects of POLS are sex-specific (Hämäläinen et al. 2018, topical collection on Pace-of-life syndromes), and that some sex-specific feedbacks between life history and behaviors exist. Below, we discuss our findings in more detail and attempt to integrate the POLS hypothesis with similar conceptual frameworks from the human literature.
Life history and physiology
As predicted by the POLS hypothesis, individuals on a slow life-history trajectory had lower blood pressure (except girls for diastolic blood pressure) than those expressing a faster life history. Moreover, the present CFA results indicate a, albeit weak, relationship between life history and physiology (expressed by cardiovascular characteristics). Cardiovascular parameters, such as blood pressure, can serve as a proxy for the sympathetic/parasympathetic nervous system reactivity (Réale et al. 2010). Furthermore, high systolic blood pressure is positively linked to basal metabolic rate (Snodgrass et al. 2008), a physiological trait assumed to covary with POL (Réale et al. 2010). In the animal ecology literature, most studies investigating the link between physiology and POL have focused on metabolic parameters (Montiglio et al. 2018, topical collection on Pace-of-life syndromes; Royauté et al. 2018, topical collection on Pace-of-life syndromes), and to the best of our knowledge, none of these studies used cardiovascular parameters. In human medicine, the determinants of blood pressure are well known and include age, sex, testosterone levels (Huisman et al. 2006), parent health-risk behavior, birth weight, and obesity (Huisman et al. 2006; Simonetti et al. 2011; Patel and Walker 2016; van der Steen and Hokken-Koelega 2016). By creating homogeneous age groups and treating the sexes separately, we attempted to control for key components of variation in blood pressure. Moreover, blood pressure during childhood and adolescence appears to be a good predictor of blood pressure later in life (Chen et al. 2007). We found higher blood pressures in fast maturing girls and boys than in slow maturing ones. Sexual maturity had an influence on cardiovascular characteristics during adolescence. Our results are in line with Cho et al. (2001), they found a relevant contribution of sexual maturity status on systolic blood pressure (when controlling for height and body mass index) within the Heartfelt study, but also pointed out that the literature is not clear about contribution of sexual maturity on variation in cardiovascular characteristics. In addition, systolic and diastolic blood pressure values are higher in adult women with earlier menarcheal age (Dreyfus et al. 2015; Jelenkovic and Rebato 2016).
In contrast to blood pressure, heart rate was lower in boys and girls maturing fast, as compared to those maturing slower and, thus, the pattern for heart rate was against our prediction. Heart rate variation is an indicator of the stress response in animals and humans (e.g. Koolhaas et al. 1999, 2010; Thayer et al. 2012). Unfortunately, the KiGGS data set contained only one measurement of heart rate, preventing us from assessing heart rate variation in response to stress. Heart rate is under regulation of the sympathetic- and parasympathetic antagonists’ system. The parasympathetic control increases from infancy to adulthood. Hence, earlier puberty could be associated to increased parasympathetic tone; consequently, heart rate is lower (Graziano and Derefinko 2013; Ellis et al. 2017) and reflects acceleration towards a slower maturation schedule. Similarly, under the adaptive calibration model of stress responsivity (ACM: Del Giudice et al. 2011; Ellis et al. 2017) low basal heart rate could be linked to a faster life history under certain local environmental condition. Without further data on early environmental conditions and repeated physiological measures of the subjects testing, these more nuanced predictions remain outside the scope of our study.
Overall, our results bear some similarity to the findings of Figueredo et al. (2004): Using data of the National Survey for Midlife Development in the USA, they found that a set of correlated psychosocial traits (K), their measure for POL was highly correlated with a general factor for overall physical and mental health (see also below). Though this study did not focus on age of maturation per se, it lends support for a tight association between POL and physiological parameters. Taken together, in humans, age of maturation, a key life-history trait, appears to covary with non-metabolic physiological traits, such as cardiovascular parameters, that are known to affect physical activity and are part of animal coping styles.
Life history and behavior
Generally, fast maturing boys and girls tended to display higher levels of health-related risk-taking behaviors than their same-age peers. Particularly, tobacco and alcohol use and inadequate physical activity were substantially higher in fast maturing girls and boys. In girls, fast maturing individuals also displayed higher levels of unhealthy dietary behavior and physical violence. Fast maturing boys had higher levels of media consumption than their slower maturing peers. Proximately, these differences are likely due to elevated sex hormones in matured individuals as compared to immature peers, driving both POL and the expression of health-related risk-taking behavior. Still, it remains to be explained, why hormones are upregulated earlier in some individuals than others.
Among our set of risk-taking components, some represent health-risk behavior more directly than others. Particularly for active and direct risk-taking behaviors, such as smoking and drinking alcohol, we found support for our prediction of higher risk taking in faster life histories. Thus, following the predictions from the POLS hypothesis, same-aged individuals with a faster life history showed increased risk-taking behavior in terms of unhealthy substance use, reduced physical activity and increased physical violence risk. Similar to the POLS hypothesis, (Ellis et al. 2012) proposed in their “evolutionary model of risky behavior in adolescence” that engaging in risky behavior can be beneficial in concert with a fast life history and under certain environmental conditions. Human psychological literature provides a wealth of studies demonstrating various links between personality and life history (summarized, e.g., in Réale and Dingemanse 2010; Figueredo et al. 2013). Particularly, fast maturing boys and girls display higher levels of aggressive and delinquent behavior than slow maturing (Najman et al. 2009) and sensation seeking and drug use during adolescence compared to same-aged peers (references in Ellis et al. 2012). Some of these differences between slow and fast maturing adolescents can persist through adulthood, such as initial differences in physical strength, body height and dominance in boys carrying over to manhood (references in Ellis et al. 2012). In sum, girls and boys pursuing a fast life-history trajectory score higher in various risk-taking behaviors during a critical transition phase in life, a phase characterized by a substantial increase in morbidity and mortality (Ellis et al. 2012).
Given that much is at stake and that several characteristics expressed in adolescence can have long-lasting consequences, risk-taking behavior might be one mediator of the trade-off between current and future reproduction in humans. However, the above-described differences in physiological and behavioral traits between slow and fast life-history groups could also be the result of state-dependent plasticity based on the developmental status of an individual. For instance, cardiovascular characteristics or the amount of risk taking could change when individuals advance from pre-puberty to puberty and adolescence. A longitudinal study with several consecutive measurements of risk-taking behavior, cardiovascular characteristics and life-history traits is required to rule out this possibility.
Behavior and physiology
We also explored potential links between health-related risk-taking behavior and physiology at the phenotypic level, assuming that positive correlations would indicate that both sets of traits mediate the trade-off between current and future reproduction in a similar way (direction) and, thus, would form suits of integrated characteristics along a slow-fast POL continuum. We found mixed support for this prediction. Within our CFA analysis neither in girls nor in boys’ health-related risk-taking behavior was linked to physiology at the phenotypic level. But among all single risk-taking behaviors, only inadequate physical activity correlated positively at the phenotypic level with all (except pulse pressure) cardiovascular parameters in girls and boys. Other single risk-taking parameters (unhealthy dietary behavior, media consumption, alcohol use or physical violence) correlated positively with at least one single cardiovascular characteristic in girls or in boys. In addition, physical violence, active tobacco use, alcohol use, neglecting protection, neglecting protection 2, and media consumption showed negative correlations with at least one cardiovascular characteristic. Notably, the direction and strength of these phenotypic correlations varied between the sexes.
Although not directly predicted by the POLS hypothesis, behavioral and physiological traits are often forming syndromes. Within the framework of POLS, correlations between aspects of energy metabolism and behaviors related to energy acquisition (e.g., foraging) or energy expenditure (e.g., mating) have received much attention (Careau et al. 2008; Mathot and Dingemanse 2015). Another well-known syndrome in animal behavior are coping styles, which are correlated behavioral and physiological traits characterizing the stress response of an individual, jointly regulated by sympathetic and parasympathetic reactivity (Koolhaas et al. 1999; Koolhaas et al. 2010). For example, risk-taking (“proactive”) individuals are assumed to have a different coping style than less risk-taking (“reactive”) individuals. Proximately, a high sympathetic reactivity and low parasympathetic reactivity lead to a high but consistent heart rate under stressors in proactive individuals (Koolhaas et al. 1999; Ellis et al. 2006), allowing them to cope with the source of stress through a “flight-or-fight” response. Reactive individuals are assumed to show the opposite physiological patterns and tend to react by freezing. In animals, cardiovascular parameters are considered a part of coping styles. For example, proactive individuals develop hypertension more often than reactive animals due to increased sympathetic reactivity (Koolhaas et al. 2010 and references therein). In humans, cardiovascular parameters have been found to correlate with personality; for example, the amplitude patterns of electrocardiogram (ECG) correlated with various personality scores in healthy individuals (Koelsch et al. 2012). Thus, although personality and physiology covary in many species, there is only mixed support for cardiovascular physiology traits and risk-taking mediating the life-history trade-off between current and future reproduction in the same way, at least based on the set of traits used in our study.
Sex-specific POLS in humans
Although the overall link between age of maturation and physiology and behavior was largely similar in both sexes, we found several sex differences: for physiology, POLS was better supported in boys than in girls, whereas for risk-taking behavior POLS was better supported for girls than boys. In addition, CFA results revealed slight differences between girls and boys in terms of health-related risk behavior (inadequate physical activity, active tobacco use and alcohol use) and the strength of the association between physiology with life history (weaker in girls than in boys) and between health-related risk-taking behavior and life history (weaker in boys than in girls) (see Fig. 3, Table 2). Moreover, differences between fast and slow maturing individuals were sex-specific for a number of single risk-taking behaviors. Against the POLS hypothesis and in contrast to boys, fast maturing girls had lower scores in media consumption than slow maturing peers. In our data set, boys had much higher values for this variable. For example, 6.4% of boys reported playing video games for more than 2 h/day, compared to only 1.1% in girls. Hence, this variable might reflect unhealthy behavior better for boys than girls. Conversely, unhealthy dietary behavior was expressed more in fast maturing girls than slow maturing girls, but in boys, the pattern was reversed. For this behavior, sex differences are well known (Croll et al. 2002; Swanson et al. 2011) and, in agreement with previous studies, we also found a lower propensity of eating disorders in boys.
Apart from sex-specific importance of certain risk-taking behaviors, the differences between the sexes might be due to two, non-mutually excluding, causes. First, the differences between life-history groups might increase with age and boys are on average 1 year older than girls in our life-history groups. Although this might explain why the differences between fast and slow maturing boys are stronger than between fast and slow maturing girls in physiological parameters, it is unlikely to explain differences between the sexes for health-related risk-taking behaviors. Second, sex differences in POLS might have an adaptive value (Hämäläinen et al. 2018, topical collection on Pace-of-life syndromes). In humans and many other species, males and females differ in life history, physiological and behavioral traits (summarized in Hämäläinen et al. 2018, topical collection on Pace-of-life syndromes; Immonen et al. 2018, topical collection on Pace-of-life syndromes, see also Del Giudice and Belsky 2011) and sex-specific selection pressures might generate differences in mean trait expressions, strength of trait correlations and covariance structures (Hämäläinen et al. 2018, topical collection on Pace-of-life syndromes). Exploring sex differences in POLS in humans in more detail and from an evolutionary perspective (see, e.g., Ellis et al. 2011), will be a fascinating area of future research.
A human pace-of-life syndrome?
Several conceptual frameworks exist that are similar to the POLS hypothesis (Réale et al. 2010). For example, Rushton’s (1985) differential K theory posits that among-individual variation in behavioral and physiological traits is related to slow-fast life-history strategies. This framework has stimulated much empirical work (summarized in Figueredo et al. 2013), revealing that (i) suites of psychosocial measures form a higher order life-history factor (termed K), representing a slow-fast continuum; (ii) an integrated, general factor of personality (termed GFP) phenotypcially correlates with K; (iii) an integrated, general factor of overall physical and mental health (termed covitality) phenotypcially correlates with K; and (iv) these correlations have a significant genetic component (e.g., Figueredo et al. 2004, 2005, 2007; Figueredo and Rushton 2009). Further, Ellis et al. (2009) proposed a developmental theory of inter-individual variation in humans, which posits that correlated life-history traits form a slow-fast continuum and evolved in response to environmental harshness and unpredictability. Subsequently, risk-taking behavior, particularly during adolescence, was incorporated into this view of human life-history syndromes (Ellis et al. 2012). Brumbach et al. (2009) provided support for this framework by showing that frequent changes or ongoing inconsistency in childhood environments negatively affected health and risk taking (their proxies for life-history strategies) in a representative sample of adolescents and young adults in the USA. Moreover, resource availability during childhood determined how mortality cues affected gambling decisions: individuals who grew up relatively poor favored risky but immediate rewards when confronted with mortality cues, whereas individuals who grew up relatively wealthy favored future rewards (Griskevicius et al. 2011). Other researchers also highlighted that physiological characteristics, particularly stress response, should covary with life-history strategies because they evolved in response to the same environmental conditions that shaped life-history variation (e.g., Del Giudice and Belsky 2011; Del Giudice et al. 2011). Thus, many ideas present in the human literature bear close resemblance to the POLS hypothesis, but they also differ in some key details. All of these frameworks are based on life-history theory (Pianka 1970; Stearns 1982) and view life-history strategies as evolved adaptive responses to certain environmental conditions (e.g. Ellis et al. 2009; Del Giudice and Belsky 2011). However, all concepts in the human literature are either specified for humans with their unique life history, differing from most other animals, and their specific socio-cultural environmental conditions and/or use psychosocial instead of biological measures for POL (e.g., Figueredo et al. 2004, 2005, 2007; Figueredo and Rushton 2009). In our opinion, the POLS hypothesis goes beyond these other frameworks by conceptually explaining the evolution of within-species variation in life-history strategies along the current versus future reproduction trade-off by the mediating function of behavioral and physiological traits (Dammhahn et al. 2018, topical collection on Pace-of-life syndromes).
Our results of links between a key life-history parameter, physiology and health-related risk-taking behavior in a contemporary German population, are plausible under the POLS hypothesis. However, formally testing POLS in humans and other animals is difficult because it requires experimental manipulation of environmental conditions and disentangling within- from between-individual correlations (Montiglio et al. 2018, topical collection on Pace-of-life syndromes). In general, raw phenotypic correlations (or simple group differences, as used in this study) are not well suited to reveal syndromes between traits because other sources of variation (e.g., within-individual variation) are not controlled for. Ideally, behavior, physiology, and life history should all be measured repeatedly and simultaneously over the course of a life time to estimate pure between-individual correlations among traits (Dingemanse et al. 2012). Only these correlations will reveal additive genetic, permanent environment or maternal effects, which can be shaped by evolution (see e.g. Figueredo and Rushton 2009). However, many life-history traits (namely age at maturity, age at first reproduction) occur only once in a lifetime. Moreover, repeated measures of complex traits, such as personality in humans, in concert with physiological measures are often not available, partly because they are studied in very different research areas (e.g., psychology and medicine), and partly because of logistic challenges of following humans over the course of their lifetime. Several governmental organizations do collect large representative data sets, such as KiGGS and MIDUS, which—although not perfect in sampling design for testing POLS theory—might provide valuable data of a large sample. Including crucial life-history traits in these surveys and applying a repeated measures design for subgroups, would allow testing POLS predictions more rigorously in the future.
Humans are special in their life history and their cultural environment (Bogin 1999), but despite human universals, we find support for the POLS hypothesis in humans. Better multivariate data sets of repeatedly measured traits, in concert with fitness data (e.g., Alvergne et al. 2010; Jokela et al. 2011), would allow ultimately testing whether behavior and physiology mediate the trade-off between current and future reproduction in humans. Combining data sets of human populations living under environmental conditions that vary in harshness and predictability (e.g., Brumbach et al. 2009) would additionally help testing the environmental drivers of POLS evolution. Previous conceptual frameworks of the human literature are more anthropocentric but can be well integrated with the POLS hypothesis. Further, uniting these concepts more thoroughly would be fruitful for the study of POLS in humans and other animals, akin to the integration of behavioral ecological approaches to animal personality and approaches and tools of personality psychology (e.g., Gosling 2001; Nettle and Penke 2010; Uher 2011).
Conclusions
Using data on life history, health-related risk-taking behavior and cardiovascular physiology of contemporary human adolescents, we found some support for key predictions of the POLS hypothesis. We are aware that our comparisons between life-history groups have some limitations and cannot replace multi-trait (co)variance analyses, but repeated measures of physiological traits in concert with behavioral traits and biological measures of life history are rare, even in the rich human literature. We hope that our work can familiarize animal ecologists with the developments taking place in human ecology; for instance, by showing that the trade-off between current and future reproductive success is only one out of several key processes that could underlie covariance between life history and behavior/physiology (Mathot and Frankenhuis 2018, topical collection on Pace-of-life syndromes).
For human ecologists, we hope this introduction to the POLS can stimulate future research exploiting other long-term data sets. Throughout, we have highlighted similarities and differences between the key approaches in animal and human ecology. By pointing out parallels and distinctions, we hope to stimulate cross-fertilization between these two research domains, which have evolved largely independent (see also Nettle 2006; Nettle and Penke 2010).
References
Alvergne A, Jokela M, Lummaa V (2010) Personality and reproductive success in a high-fertility human population. Proc Natl Acad Sci USA 107:11745–11750
Belsky J (2012) The development of human reproductive strategies: progress and prospects. Curr Dir Psychol Sci 21:310–316
Belsky J, Steinberg L, Draper P (1991) Childhood experience, interpersonal development, and reproductive strategy: an evolutionary theory of socialization. Child Dev 62:647–670
Bogin B (1999) Patterns of human growth. Cambridge University Press, Cambridge
Brener ND, Kann L, McManus T, Kinchen SA, Sundberg EC, Ross JG (2002) Reliability of the 1999 youth risk behavior survey questionnaire. J Adolesc Health 31:336–342
Brumbach BH, Figueredo AJ, Ellis BJ (2009) Effects of harsh and unpredictable environments in adolescence on development of life history strategies. Hum Nat 20:25–51
Careau V, Garland T (2012) Performance, personality, and energetics: correlation, causation, and mechanism. Physiol Biochem Zool 85:543–571
Careau V, Thomas D, Humphries M, Réale D (2008) Energy metabolism and animal personality. Oikos 117:641–653
Chen X, Wang Y, Mi J (2007) Tracking of blood pressure from childhood to adulthood: a systematic review and meta-analysis. FASEB J 21:A1363
Cho SD, Mueller WH, Meininger JC, Liehr P, Chan W (2001) Blood pressure and sexual maturity in adolescents: the heartfelt study. Am J Hum Biol 13:227–234
Croll J, Neumark-Sztainer D, Story M, Ireland M (2002) Prevalence and risk and protective factors related to disordered eating behaviors among adolescents: relationship to gender and ethnicity. J Adolescent Health 31:166–175
Dammhahn M, Dingemanse NJ, Niemelä P, Réale D (2018) Pace-of-life syndromes: a framework for the adaptive integration of behaviour, physiology and life-history. Behav Ecol Sociobiol. (in press)
Del Giudice M, Belsky J (2011) The development of life history strategies: toward a multi-stage theory. In: Buss DM, Hawley PH (eds) The evolution of personality and individual differences. Oxford University Press, Oxford, pp 154–176
Del Giudice M, Ellis BJ, Shirtcliff EA (2011) The adaptive calibration model of stress responsivity. Neurosci Biobehav Rev 35:1562–1592
Development Core Team R (2014) R: a language and environment for statistical computing. The R Foundation for Statistical Computing. Austria, Vienna http://www.R-project.org
Dingemanse NJ, Bouwman KM, van de Pol M, van Overveld T, Patrick SC, Matthysen E, Quinn JL (2012) Variation in personality and behavioural plasticity across four populations of the great tit Parus major. J Anim Ecol 81:116–126
Dreyfus J, Jacobs DR, Mueller N, Schreiner PJ, Moran A, Carnethon MR, Demerath EW (2015) Age at menarche and cardiometabolic risk in adulthood: the coronary artery risk development in young adults study. J Pediatr 167:344–352
Ellis L (1988) Criminal behavior and r/K selection: an extension of gene-based evolutionary theory. Pers Individ Differ 9:697–708
Ellis BJ, Jackson JJ, Boyce WT (2006) The stress response systems: universality and adaptive individual differences. Dev Rev 26:175–212
Ellis BJ, Figueredo AJ, Brumbach BH, Schlomer GL (2009) Fundamental dimensions of environmental risk. Hum Nat 20:204–268
Ellis BJ, Shirtcliff EA, Boyce WT, Deardorff J, Essex MJ (2011) Quality of early family relationships and the timing and tempo of puberty: effects depend on biological sensitivity to context. Dev Psychopathol 23:85–99
Ellis BJ, Del Giudice M, Dishion TJ et al (2012) The evolutionary basis of risky adolescent behavior: implications for science, policy, and practice. Dev Psychol 48:598–623
Ellis BJ, Del Giudice M, Shirtcliff EA (2017) The adaptive calibration model of stress responsitivity: concepts, findings, and implications for developmental psychopathology. In: Beauchaine TP, Hinshaw SP (eds) Child and adolescent psychopathology, 3rd edn. Wiley & Sons, New York, pp 237–276
Eveleth PB, Tanner JM (1990) Worldwide variation in human growth. Cambridge University Press, Cambridge
Fabrigar LR, Wegener DT, MacCallum RC, Strahan EJ (1999) Evaluating the use of exploratory factor analysis in psychological research. Psychol Methods 4:272–299
Fan Y, Chen J, Shirkey G, John R, Wu SR, Park H, Shao C (2016) Applications of structural equation modeling (SEM) in ecological studies: an updated review. Ecol Process 5:19–31
Figueredo AJ, Rushton JP (2009) Evidence for shared genetic dominance between the general factor of personality, mental and physical health, and life history traits. Twin Res Hum Genet 12:555–563
Figueredo AJ, Vasquez G, Brumbach BH, Schneider SM (2004) The heritability of life history strategy: the k-factor, covitality, and personality. Soc Biol 51:121–143
Figueredo AJ, Sefcek JA, Vasquez G, Brumbach BH, King JE, Jacobs WJ (2005) Evolutionary personality psychology. In: Buss DM (ed) The handbook of evolutionary psychology. John Wiley, Hoboken, pp 851–877
Figueredo AJ, Vásquez G, Brumbach BH, Stephanie M, Schneider R (2007) The k-factor, covitality, and personality. Hum Nat 18:47–73
Figueredo AJ, de Baca TC, Woodley MA (2013) The measurement of human life history strategy. Pers Individ Differ 55:251–255
Gosling SD (2001) From mice to men: what can we learn about personality from animal research? Psychol Bull 127:45–86
Graziano P, Derefinko K (2013) Cardiac vagal control and children's adaptive functioning: a meta-analysis. Biol Psychol 94:22–37
Griskevicius V, Tybur JM, Delton AW, Robertson TE (2011) The influence of mortality and socioeconomic status on risk and delayed rewards: a life history theory approach. J Pers Soc Psychol 100:1015–1026
Hämäläinen A, Immonen E, Tarka M, Schütt W (2018) Evolution of sex-specific pace-of-life syndromes: causes and consequences. Behav Ecol Sociobiol. https://doi.org/10.1007/s00265-018-2466-x
Harries M, Walker JM, Williams DM, Hawkins S, Hughes I (1997) Changes in the male voice at puberty. Arch Dis Child 77:445–447
Hill K, Kaplan H (1999) Life history traits in humans: theory and empirical studies. Annu Rev Anthropol 28:397–430
Hölling H, Kamtsiuris P, Lange M, Thierfelder W, Thamm M, Schlack R (2007) The German health interview and examination survey for children and adolescents (KiGGS): study management and conduct of fieldwork. Bundesgesundheitsbla 50:557–566
Hölling H, Schlack R, Kamtsiuris P, Butschalowsky H, Schlaud M, Kurth BM (2012) The KiGGS study. Nationwide representative longitudinal and cross-sectional study on the health of children and adolescents within the framework of health monitoring at the Robert Koch Institute. Bundesgesundheitsbla 55:836–842
Höpker T, Lampert T, Spallek J (2014) Identification and characterisation of health behaviours in 11- to 17-year-old adolescents: a cluster analysis based on the German health interview and examination survey for children and adolescents. Gesundheitswesen 76:453–461
Hoyle RH (1995) Structural equation modeling: concepts, issues, and applications. Sage, Thousand Oaks
Huisman H, Schutte A, Van Rooyen J, Malan N, Malan L, Schutte R, Kruger A (2006) The influence of testosterone on blood pressure and risk factors for cardiovascular disease in a black South African population. Ethnic Dis 16:693–698
Immonen E, Hämäläinen A, Schütt W, Tarka M (2018) Evolution of sex-specific pace-of-life syndromes: genetic architecture and physiological mechanisms. Behav Ecol Sociobiol. https://doi.org/10.1007/s00265-018-2462-1
Jelenkovic A, Rebato E (2016) Association of maternal menarcheal age with anthropometric dimensions and blood pressure in children from Greater Bilbao. Ann Hum Biol 43:430–437
Jokela M, Kivimäki M, Elovainio M, Keltikangas-Järvinen L (2009) Personality and having children: a two-way relationship. J Pers Soc Psychol 96:218–230
Jokela M, Alvergne A, Pollet TV, Lummaa V (2011) Reproductive behavior and personality traits of the five factor model. Eur J Pers 25:487–500
Juul A, Magnusdottir S, Scheike T, Prytz S, Skakkebæk NE (2007) Age at voice break in Danish boys: effects of pre-pubertal body mass index and secular trend. Int J Androl 30:537–542
Kamtsiuris P, Lange M, Rosario AS (2007) Der Kinder- und Jugendgesundheitssurvey (KiGGS): Stichprobendesign, Response und Nonresponse-Analyse. Bundesgesundheitsbla 50:547–556
Kann L, McManus T, Harris WA et al (2016) Youth risk behavior surveillance—United States, 2015. MMWR Surveill Summ 65:1–174
Kaplan H, Hill K, Lancaster J, Hurtado AM (2000) A theory of human life history evolution: diet, intelligence, and longevity. Evol Anthropol 9:156–185
Kline RB (2011) Principles and practice of structural equation modeling. The Guilford Press, New York
Koelsch S, Enge J, Jentschke S (2012) Cardiac signatures of personality. PLoS One 7:e31441
Koolhaas J, Korte S, de Boer S, van der Vegt B, van Reenen C, Hopster H, de Jong I, Ruis M, Blokhuis H (1999) Coping styles in animals: current status in behavior and stress-physiology. Neurosci Biobehav Rev 23:925–935
Koolhaas J, de Boer S, Coppens C, Buwalda B (2010) Neuroendocrinology of coping styles: towards understanding the biology of individual variation. Front Neuroendocrinol 31:307–321
Korkmaz S, Goksuluk D, Zararsiz G (2014) MVN: an R package for assessing multivariate normality. R J 6:151–162
Kurth B-M, Kamtsiuris P, Hölling H, Schlaud M, Dölle R, Ellert U, Kahl H, Knopf H, Lange M, Mensink GB (2008) The challenge of comprehensively mapping children’s health in a nation-wide health survey: design of the German KiGGS-study. BMC Public Health 8:196
Linting M, van der Kooij A (2012) Nonlinear principal components analysis with CATPCA: a tutorial. J Pers Assess 94:12–25
Linting M, Meulman JJ, Groenen PJ, van der Koojj AJ (2007) Nonlinear principal components analysis: introduction and application. Psychol Methods 12:336–358
MacArthur RH (1962) Some generalized theorems of natural selection. Proc Natl Acad Sci USA 48:1893–1897
MacArthur RH, Wilson EO (2015) Theory of island biogeography (MPB-1), vol. 1. Princeton University Press, Princeton
Mardia KV (1970) Measures of multivariate skewnees and kurtosis with applications. Biometrika 57:519–530
Marshall WA, Tanner JM (1969) Variations in pattern of pubertal changes in girls. Arch Dis Child 44:291–303
Mathot KJ, Dingemanse NJ (2015) Energetics and behavior: unrequited needs and new directions. Trends Ecol Evol 30:199–206
Mathot KJ, Frankenhuis WE (2018) Models of pace-of-life syndromes (POLS): a systematic review. Behav Ecol Sociobiol. https://doi.org/10.1007/s00265-018-2459-9
Montiglio P-O, Dammhahn M, Dubuc-Messier G, Réale D (2018) The pace-of-life syndrome revisited: the role of ecological conditions and natural history on the slow-fast continuum. Behav Ecol Sociobiol. (in press)
Najman JM, Hayatbakhsh MR, McGee TR, Bor W, O'Callaghan MJ, Williams GM (2009) The impact of puberty on aggression/delinquency: adolescence to young adulthood. Aust N Z J Criminol 42:369–386
National Research Council (2011) The science of adolescent risk-taking: workshop report. National Academies Press, Washington, DC
Nettle D (2005) An evolutionary approach to the extraversion continuum. Evol Hum Behav 26:363–373
Nettle D (2006) The evolution of personality variation in humans and other animals. Am Psychol 61:622–631
Nettle D, Penke L (2010) Personality: bridging the literatures from human psychology and behavioural ecology. Philos Trans R Soc B 365:4043–4050
Neuhauser H, Thamm M (2007) Blutdruckmessung im Kinder- und Jugendgesundheitssurvey (KiGGS). Bundesgesundheitsbla 50:728–735
Neuhauser HK, Thamm M, Ellert U, Hense HW, Rosario AS (2011) Blood pressure percentiles by age and height from nonoverweight children and adolescents in Germany. Pediatrics 127:e978-e988
Oli MK (2004) The fast–slow continuum and mammalian life-history patterns: an empirical evaluation. Basic Appl Ecol 5:449–463
Patel N, Walker N (2016) Clinical assessment of hypertension in children. Clin Hypertens 22:15–18
Pettay JE, Helle S, Jokela J, Lummaa V (2007) Natural selection on female life-history traits in relation to socio-economic class in pre-industrial human populations. PLoS One 2:e606
Pianka ER (1970) On r-and K-selection. Am Nat 104:592–597
Réale D, Dingemanse NJ (2010) Personality and individual social specialisation. In: Szekely T, Moore AJ, Komdeur J (eds) Social behaviour: genes, ecology and evolution. Cambridge University Press, New York, pp 417–441
Réale D, Garant D, Humphries MM, Bergeron P, Careau V, Montiglio PO (2010) Personality and the emergence of the pace-of-life syndrome concept at the population level. Philos Trans R Soc B 365:4051–4063
Rosario AS, Kurth B-M, Stolzenberg H, Ellert U, Neuhauser H (2010) Body mass index percentiles for children and adolescents in Germany based on a nationally representative sample (KiGGS 2003–2006). Eur J Clin Nutr 64:341–349
Rosseel Y (2012) lavaan: an R package for structural equation modeling. J Stat Softw 48:1–36
Royauté R, Berdal MA, Garrison CR, Dochtermann NA (2018) Paceless life? A meta-analysis of the pace-of- life syndrome hypothesis. Behav Ecol Sociobiol. https://doi.org/10.1007/s00265-018-2472-z
Rushton JP (1985) Differential K theory: the sociobiology of individual and group differences. Pers Indiv Differ 6:441–452
Simonetti GD, Schwertz R, Klett M, Hoffmann GF, Schaefer F, Wühl E (2011) Determinants of blood pressure in preschool children. The role of parental smoking. Circulation 123:292–298
Smith BH (1992) Life history and the evolution of human maturation. Evol Anthropol 1:134–142
Snodgrass JJ, Leonard WR, Sorensen MV, Tarskaia LA, Mosher MJ (2008) The influence of basal metabolic rate on blood pressure among indigenous Siberians. Am J Phys Anthropol 137:145–155
Stearns S (1982) The role of development in the evolution of life histories. In: Bonner JT (ed) Evolution and development. Dahlem workshop reports (life sciences research report), vol. 22. Springer, Berlin, pp 237–258
Stearns SC (1989) Trade-offs in life-history evolution. Funct Ecol 3:259–268
Stearns SC (1992) The evolution of life histories. Oxford University Press, Oxford
Steinberg L (2008) A social neuroscience perspective on adolescent risk-taking. Dev Rev 28:78–106
Susanne C, Bodzsar E (1998) Patterns of secular change of growth and development. In: Bodzsar EB, Susanne C (eds) Secular growth changes in Europe, 1st edn. Eötvös University Press, Budapest, pp 5–26
Swanson SA, Crow SJ, Le Grange D, Swendsen J, Merikangas KR (2011) Prevalence and correlates of eating disorders in adolescents: results from the national comorbidity survey replication adolescent supplement. Arch Gen Psychiat 68:714–723
Tanner J (1962) Growth at adolescence. Blackwell Scientific Publications, Oxford
Thayer JF, Åhs F, Fredrikson M, Sollers JJ, Wager TD (2012) A meta-analysis of heart rate variability and neuroimaging studies: implications for heart rate variability as a marker of stress and health. Neurosci Biobehav Rev 36:747–756
Uher J (2011) Personality in nonhuman primates: what can we learn from human personality psychology? In: Weiss A, King J, Murray L (eds) Personality and temperament in nonhuman primates. Springer, New York, pp 41–76
Ullman JB (2006) Structural equation modeling: reviewing the basics and moving forward. J Pers Assess 87:35–50
van der Steen M, Hokken-Koelega AC (2016) Growth and metabolism in children born small for gestational age. Endocrin Metab Clin 45:283–294
Viechtbauer W (2010) Conducting meta-analyses in R with the metafor package. J Stat Softw 36:1–48
Walker R, Gurven M, Hill K, Migliano A, Chagnon N, De Souza R, Djurovic G, Hames R, Hurtado AM, Kaplan H (2006) Growth rates and life histories in twenty-two small-scale societies. Am J Hum Biol 18:295–311
Worthman CM (1999) Evolutionary perspectives on the onset of puberty. Evol Med:135–163
Acknowledgments
We thank all participants of the two workshops Towards a general theory of the pace-of-life syndrome, held in Hannover in 2015 and 2016, for inspiring discussions as well as the Volkswagen Stiftung (Az. 89905) for funding these workshops. We thank Marco Del Giudice, Denis Réale, Willem Frankenhuis and one anonymous reviewer as well as members of the Animal Ecology group at the University of Potsdam for providing constructive comments on earlier versions of the manuscript.
Funding
This study and two workshops Towards a general theory of the pace of life syndrome were funded by Volkswagen Stiftung (Az. 89905).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
This article does not contain any studies with human participants performed by any of the authors. The KiGGS survey was reviewed and approved by the responsible ethics committee at the University Hospital of the Charité of the Humboldt University in Berlin.
Additional information
Communicated by D. Réale
This article is a contribution to the Topical Collection Pace-of-life syndromes: a framework for the adaptive integration of behavior, physiology and life-history - Guest Editors: Melanie Dammhahn, Niels J. Dingemanse, Petri T. Niemelä, Denis Réale.
Rights and permissions
About this article

Cite this article
Lehmann, A., Eccard, J.A., Scheffler, C. et al. Under pressure: human adolescents express a pace-of-life syndrome. Behav Ecol Sociobiol 72, 57 (2018). https://doi.org/10.1007/s00265-018-2465-y
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-018-2465-y