
Abstract
Behavioral ecologists have assumed that animals will optimize behavior to match their current situation, yet studies reveal limited behavioral flexibility and extensive variation among individuals in their response to a similar stimulus. This phenomenon is referred to as “animal personality,” and recent efforts seek to determine the implications of personality for interactions with the biotic and abiotic environment. Doing so promises to illuminate how selection is operating on behavioral traits and, consequently, how stable, among-individual variation in behavioral types is maintained evolutionarily. We examined a natural population of forest-dwelling eastern box turtles (Terrapene carolina) to understand the effects of differences in levels of boldness on thermoregulatory behavior, injury history, and movement rates. Repeated behavioral trials revealed that individual turtles varied consistently and dramatically in boldness (latency to emerge from the shell or begin moving after handling and confinement). Behavioral differences were independent of phenotypic state or environmental conditions, suggesting the existence of personality in these turtles. Moreover, bolder turtles maintained higher body temperatures in the field and tended to have more damage to their shells, possibly indicative of more frequent predation attempts. Little work has been done on personality in terrestrial ectotherms, and this study enhances our understanding of this phenomenon by highlighting the potential trade-off between mortality and thermoregulation.
Significance statement
Animals often exhibit markedly different personality traits from each other. We sought to understand how personality traits affect interactions with the environment, and thus how natural selection may be acting upon and maintaining behavioral variation. We studied this in terrestrial ectotherms, box turtles, for whom the trade-off between basking and predation risk may be a significant factor. We found that individual turtles differed consistently in boldness (time spent hiding and immobile after handling) and that bolder turtles maintained higher body temperatures but tended to have more injuries on their shells. This suggests that being bold is beneficial for thermoregulation, which may enhance growth and fecundity, but could expose turtles to greater risk of predation. Variation in the expression of boldness may, therefore, serve to maintain behavioral variation in the population.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Behavioral ecologists have historically assumed that individuals will optimize their behavior to best fit their biotic and abiotic environment. Consequently, variation in behavior among conspecific individuals occupying the same environment was regarded as statistical noise and received little attention (Sih et al. 2004). Recently, there has been increasing recognition of the importance of, and degree of, behavioral variation within populations (Sih and Bell 2008). The occurrence of systematic, stable, and within-population differences in behavioral tendencies has been described as “animal personality” (or, variably, “temperament” or “behavioral syndrome”; MacKay and Haskell 2015). Evidence for personality is widespread in animals and is typically identified as significantly repeatable individual differences in behavior that are not attributable to other clear phenotypic differences (e.g., age or sex; Bell et al. 2009). These findings have raised the critical questions of how such variation is generated and maintained; that is, why has selection against suboptimal behaviors not eroded personality differences? To address this challenge, various models have been proposed such as frequency-dependent selection (Wolf and McNamara 2012), individual niche specialization (Bergmüller and Taborsky 2010), or covariation with stable differences in state (Dall et al. 2004), such as physiology (Adriaenssens et al. 2016) or morphology (Thomas et al. 2016), or with life history strategy (Biro and Stamps 2008). Furthermore, better understanding of the evolution of animal personalities is increasingly important as personality may have profound consequences for evolutionary processes (e.g., speciation; Wolf and Weissing 2012), ecological interactions (Sih et al. 2012), and conservation (e.g., reintroduction efforts; Bremner-Harrison et al. 2004).
To understand both how personality evolved and the broader consequences of personality differences, it is necessary to characterize how personality differences within populations affect individual interactions with the environment, which ultimately determines fitness. Behavioral differences must be consequential in the context of an animal’s lifestyle to be accounted for by adaptive evolutionary models of personality differences or to be of any significance for ecological processes or applied concerns. To this end, it is generally true that individual differences in personality traits alter interactions with the biotic and abiotic environment and often impact fitness (Smith and Blumstein 2008). For example, Debeffe et al. (2014) found that roe deer fawns with lower levels of neophobia were more likely to disperse, and Boyer et al. (2010) linked the level of parasite infections to exploratory behaviors in a population of Siberian chipmunks. A personality trait of particular interest is boldness (the propensity to engage in risky versus cautious behavior; Wilson et al. 1994), as it has been particularly well studied in many taxa, and frequently affects the survival of individuals and their interactions with the environment (Smith and Blumstein 2008). Studies of fish (e.g., Hirsch et al. 2017), birds (e.g., Myers and Hyman 2016), reptiles (e.g., Carter et al. 2010), and mammals (e.g., Kanda et al. 2012) have all indicated that boldness can be easily measured, is highly repeatable within individuals and variable among individuals, and can have significant effects on ecologically important variables. Therefore, boldness is an excellent focal personality trait.
Boldness, along with other personality traits, has been implicated in the generation of growth-mortality trade-offs (Stamps 2007), wherein bolder phenotypes are associated with reduced survival (due to risky behavior; Ballew et al. 2017) but enhanced growth or fecundity (due to resource acquisition; Biro et al. 2014). Importantly, such a trade-off between components of fitness may enable the evolutionary maintenance of behavioral diversity (Dall et al. 2004). Additionally, theoretical work indicates that individuals expressing higher levels of boldness should also have higher metabolic rates, linking physiological covariation to the growth-mortality trade-off paradigm (Careau et al. 2008). This expectation has been evaluated with mixed support in studies in fish (e.g., Binder et al. 2016), birds (e.g., Mathot et al. 2014), and mammals (e.g., Guenther and Trillmich 2015), but there has been a lack of research on the relationships between boldness, survival, and metabolism in terrestrial ectothermic vertebrates (i.e., non-avian reptiles and amphibians), for which there has also been little study of personality traits in general (but see Mell et al. 2016; Goulet et al. 2017b). Notably, metabolic rates (and performance in general) in terrestrial ectotherms are usually highly dependent on thermoregulatory behavior and access to thermal resources (e.g., basking locations), as they have little metabolic heat production and typically occupy a thermally heterogenous landscape (Basson et al. 2017). The metabolic benefits of basking, however, are countered by the predation risks involved in being exposed outside of refuge (Huey and Slatkin 1976). Altogether, this suggests that the syndrome linking metabolic rates, survival, and boldness in ectotherms could be mediated in part by thermoregulatory behavior—bolder individuals will bask more and therefore experience greater predation risk due to exposure, but will also have greater growth and fecundity due to metabolic benefits. In support of this, lizards (Lampropholis delicata) have been characterized by individual thermal types along a cold-hot continuum, with “hot” individuals preferring higher body temperatures and having greater locomotor performance than “cold” individuals (Goulet et al. 2017a). Moreover, individual lizards with bold, exploratory, and active personalities also preferred higher body temperatures (Goulet et al. 2017b), supporting the possible link between boldness and thermoregulatory behavior in ectotherms. In another study, Carter et al. (2010) found that lizards (Agama planiceps) that are bolder had higher rates of injury (tail loss) but spent more time basking, though the latter was largely attributed to its role in territorial displays. We know of no studies that have jointly examined thermoregulatory behavior, predation risk, and boldness in a wild ectotherm population, which is critical for assessing the ecological consequences of personality (Archard and Braithwaite 2010).
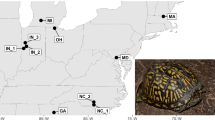
We examined boldness in eastern box turtles (Terrapene carolina; Fig. 1), a widespread but threatened species found in the eastern USA (Dodd 2001). Box turtles have been anecdotally observed to exhibit among-individual behavioral variation in response to humans (Dodd 2001, p. 140), and individual turtles vary reliably in emergence times from winter dormancy (DeGregorio et al. 2017), collectively suggesting that variation in personality types (and perhaps thermal types) may be present within these turtles. As terrestrial ectotherms (Gatten 1974) often found in forests with dense canopy cover, eastern box turtles occupy habitats with limited access to sunlight for basking, often relying on habitat edges and small gaps in the canopy to escape the shade. The lack of concealment at basking sites and the movement (at a slow pace) required to access these sites likely exposes these typically secretive turtles to predation risk, creating an appropriate system for studying ectotherm personality as it pertains to boldness, thermoregulation, and mortality. Furthermore, damages to the shell of turtles can serve as a record of previous encounters with predators (as well as aggressive interactions with conspecifics; Dodd et al. 1997), and equipment for tracking and logging environmental data can readily be affixed to turtle shells, making turtles an experimentally tractable study subject.

Example eastern box turtles (Terrapene c. carolina) from the study population. The turtle on the left is hiding in the shell with the plastron (ventral portion) angled upward and largely protecting the head, whereas the turtle on the right has emerged from the shell
In this study, we evaluated boldness repeatedly in individual, free-ranging box turtles and tested whether boldness was related to ecologically important characteristics that may mediate selection on boldness: thermoregulation, shell injuries (i.e., predation risk), and movement behavior. First, we hypothesized that individual box turtles would exhibit consistent latencies to extend their head from the shell and begin moving after standardized handling by humans (Ibáñez et al. 2014), reflective of a shy-bold axis of personality variation, even after accounting for differences in age, sex, and size. Second, we predicted that bolder turtles would experience higher body temperatures due to greater access to and/or use of basking areas. We tested this by correlating mean behavior scores with temperatures experienced in the field as recorded on data loggers mounted on the turtles. Next, we expected that bolder turtles would have more damage due to higher encounter rates with predators. Alternatively, shyer turtles may have more damage due to a greater probability of having survived encounters with predators (whereas bold turtles who sustain such injuries are also killed because of reduced hiding behavior). We tested this by correlating mean behavior scores with shell injury scores. Finally, we predicted that bolder turtles would exhibit greater rates of movement. This is supported by the following points: activity levels are often positively correlated with boldness in behavioral syndromes (Sih et al. 2004); boldness in lizards is associated with larger home ranges (Carter et al. 2010); and maintaining higher body temperatures in bold individuals may require more movement to access shifting thermal resources. Alternatively, bolder turtles may occupy better territories and therefore require less movement. We tested this by correlating mean behavior scores with estimates of home range size and rates of movement calculated using relocations of individual turtles.
Methods
Study site
Eastern box turtles were studied at Allee Memorial Woods (AMW), a 72-ha nature preserve in Parke County, Indiana. AMW consists largely of old growth (virgin) forest, as well as some secondary forest growth. Intermittent canopy openings provide areas of sunlight on the forest floor. The terrain is characterized by several steep gorges with small streams at the bottom. From 1958 to 1983, the box turtle population at AMW was the subject of research focused on demographic characteristics of the population (using mark-recapture methods; Williams and Parker 1987). Turtles marked in that earlier work were still alive and included in this study, but presumably had little interaction with humans in over 30 years.
Initial capture and data collection
Turtles were collected by searching on foot June 2–15, 2015, and May 16–25, 2016. We collected 19 box turtles in 2015 and 14 additional turtles in 2016. Because turtles had to be observed to be included in the study, it is possible that particularly secretive individuals would have been underrepresented in the sample (Biro and Dingemanse 2009). This would yield a conservative underestimate of the extent of behavioral variation in the population.
Initial handling of the turtles differed between years. In 2015, we placed newly captured turtles in a zippered black nylon bag until they could be processed (approximately 30–90 min). We then brought them to a common location on site and, as part of disease surveillance, swabbed the mouth and cloaca of each turtle and drew approximately 0.5 mL of blood from the subcarapacial sinus. In 2016, we immediately drew approximately 0.2 mL of blood, placed the turtles in a black nylon bag on the ground at the point of capture for 30 min, drew another 0.2 mL of blood, and then observed the behavior of the turtles for up to 10 min, as part of a study on stress-induced changes in hormone levels.
In both years, we then measured the length and width of the carapace using calipers and weighed the turtles with a spring scale. We sexed each turtle by a thorough evaluation of the presence of an indentation on the plastron, a flared carapace margin, the position of the cloaca relative to the tail, and coloration (Dodd 2001). We also estimated the age of the turtles by counting the number of annuli (growth rings) in the scutes of the carapace, which provide an approximate but inexact estimate of age until about 15–20 years (Wilson et al. 2003). We developed an age classification system to extend the estimates of age beyond the easily counted 15 annuli (and indeed we knew based on prior marks that several turtles were ≥ 68 years old). We classified the turtles into one of four age classes: 1—turtles with a carapace length < 115 mm and considered juveniles (Williams and Parker 1987); 2—growing adults with fewer than 15 annuli but > 115 mm in length; 3—non-growing adults with ≥ 15 well-defined annuli; and 4—older adults with vertebral annuli that had become worn and uncountable with age.
We assessed the injuries to the carapace and plastron following the system of Saumure et al. (2007). Briefly, the carapace and plastron were each divided into four quadrants, and each quadrant was scored on a 0–3 scale, indicating the most severe injury to that quadrant. The scale ranged from no injuries (0), small damages only to the scute layer (1), small damages to both the scute and underlying bone, large areas of damage confined to the scute, or missing portions at the margins of the shell (2), and large damages to both the scute and bone layer (e.g., fractures; 3). Very shallow scratches and scrapes, such as would occur while crawling through brush or burrowing, were not considered. We also did not include any holes drilled in the margin of the carapace for marking purposes during prior research activities. We added together the scores for each quadrant of the carapace and plastron to produce a single measure of the severity of injuries sustained, with potential values ranging from 0 to 24. Most of these injuries were likely derived from predators, though vehicles, lawnmowers, and farm equipment could be a contributing factor on occasions when these turtles leave the forest. Known predators (Dodd 2001, p. 140) that have been documented at the AMW site include raccoons (Procyon lotor), coyotes (Canis latrans), and Virginia opossum (Didelphis virginiana), and both raccoons and coyotes have been observed or photographed with game cameras during the activity period of the turtles (B.E. Carlson, personal observations). While these predators can successfully kill adult box turtles (Dodd 2001, p. 140), it is possible that many of these injuries were inflicted as juveniles when the shells were weaker and more easily fit into the mouth of medium-sized carnivores.
If turtles lacked a tag from previous research, we marked them by filing notches into the marginal scutes of the carapace (Ernst et al. 1974). We then used a 5-min epoxy gel (Devcon, Danvers, MA) to attach a 15.3-g radio transmitter (R1860, Advanced Telemetry Systems, Isanti, MN) to the pleural scutes on one side of the carapace. A Thermochron iButton temperature logger (DS1921G, Maxim Integrated, San Jose, CA) was similarly attached on the opposite side of the carapace to monitor body temperatures. Carapace and internal body temperatures correlate well in box turtles (do Amaral et al. 2002), including when carapacial temperatures are measured with iButtons as in this study (Bernstein and Black 2005). Prior to attachment, the iButtons were programmed to record temperature every 30 min and weather-sealed with a black plastic coating (Plasti Dip, Blaine, MN), which minimally influences the reliability of temperature readings (Roznik and Alford 2012). The combined weight of the radio transmitter, the iButton, and the epoxy was approximately 19.4–22.8 g, which ranged from 3.5–6% of turtle body weight. After the epoxy had set, we released the turtles at the location of capture. The time from initial capture to release ranged from approximately 1–3 h.
Tracking and behavioral assays
After a 6 to 8-day recovery period, we began tracking the turtles using a receiver and a directional antenna. Each turtle was generally tracked and behaviorally assayed twice per week, though practical limitations and difficulties relocating turtles resulted in 2 to 12-day intervals (mean = 3.4 days) between consecutive encounters with the same turtle. All behavioral assays were performed during the daytime between 0900 and 1700.
When located, we placed the turtle in a black, heavy-duty nylon bag for 3 min. In 2016, we used an infrared thermometer (Fluke 62 Max, Fluke Corporation, Everett, WA) to record the plastron temperature prior to placing the turtle in the bag. While the turtle was in the bag, we recorded the coordinates of its capture location using a MobileMapper 20 GPS unit (Spectra-Precision, Westminster, CO) to a stated accuracy of approximately ± 2 m, the air temperature and relative humidity (as percent), both with a Mini Environmental Quality Meter (Sper Scientific, Scottsdale, AZ), the percent canopy cover (using a spherical densiometer; Forest Densiometers, Rapid City, SD), and whether the turtle was found exposed (partly or entirely) or concealed (under leaves, buried in mud, etc.). During this time, the turtle would have been able to hear the researchers speaking to each other and may have been able to smell them. After 3 min, all but one of the researchers moved approximately 10 m away, and then the remaining researcher removed the turtle from the bag, placed it back on the ground, and walked 10 m away, while one of the other researchers observed the turtle through binoculars. Frequently, we had to relocate the turtle 1–2 m (and rarely up to 10 m) from its original location to ensure it was visible to the observer through vegetation, and we recorded the approximate distance turtles were displaced for inclusion in analyses. We recorded the time elapsed from placing the turtle on the ground to (1) head emergence (“emergence latency”), operationally defined as extending the head from the shell such that the eyes surpassed the anterior margin of the carapace (modified after Ibáñez et al. 2014; Fig. 1) and (2) movement, when the turtle began walking. Practical limitations prevented the use of blinding to minimize bias with respect to individual turtle identity and previous behavioral scores, though head emergence and movement were easily recognized and variable enough among individuals so as to minimize the possible influence of bias (see “Results” section). We terminated trials as soon as the turtle moved or after 10 min and then left the area.
After tracking and assaying each turtle eight times, we removed the radio transmitter and iButton. Five of the turtles either lost their iButtons or radio transmitters, or could not be relocated, and thus did not yield temperature data. Turtles that left the Allee Woods property could no longer be studied and had their iButtons and radio transmitters removed. These turtles, along with those who lost their radio transmitters or could not be relocated, resulted in seven turtles being assayed fewer than eight times (range of 2–7).
Data analysis
We first determined the repeatability of emergence and movement latencies, with the latter calculated as the elapsed time from head emergence to the first movement of the turtle. For 11/237 trials, emergence was never observed in the 10-min observation period, and we used the maximum value (600 s) as the emergence latency. This practice, though it censors the data, is representative of how slow these turtles were to emerge. For 36/237 trials, the movement latency could not be estimated as no movement occurred within the 10-min observation period. We excluded these trials to avoid calculating movement latencies that may be non-representative of observed behavior. For example, a turtle that neither moved nor emerged in the 10-min period would have a calculated movement latency of 0 s if we assigned a 600 s value for the movement time and subtracted the emergence latency (also 600 s). One to three observations of movement latency were therefore incalculable for each of 18/33 turtles.
Repeatability of behaviors was estimated both with and without correcting for the potential influence of variation in assay conditions and non-behavioral phenotypic characteristics. We used linear mixed models (LMMs) fitted with restricted maximum likelihood (REML) estimation, using the lmer function in the package “lme4” (Bates et al. 2015) in program R (version 3.1.0; R Core Team 2014). Individual turtle ID was incorporated as a random intercept, and log (x + 1) transformed emergence or movement latencies were the response variables. In the model in which we corrected for assay conditions, we also included as fixed covariates relative humidity, air temperature, whether turtles were exposed or concealed upon capture, the distance relocated by the experimenters from their initial location of capture, time of day, year (2015 or 2016), and test number (1–8, to test for habituation or learning). We also ran this same analysis with and without the plastron temperature of the turtles instead of air temperature, using only the 14 turtles in 2016 for which this was measured. Body temperature did not significantly affect either head emergence or movement latency, nor did it substantively alter the estimate of repeatability, and therefore, we focused on air temperature. In the model in which we corrected for individual phenotypic differences, we included as covariates sex, age class, carapace length, body condition, and nesting status. Body condition was calculated as the residuals from a standardized major axis regression of log-transformed mass against log-transformed carapace length. Females were considered nesting if they left Allee Woods; all but one of these turtles were located in unforested areas more suitable for nesting, and the remaining female could not be located until her return a few weeks later. This latter analysis for the effects of individual phenotypic differences is considerably less powerful than the analysis of the effects of testing conditions, as phenotypic values remained the same across repeated samples of individuals, and therefore, the functional sample size is dictated by the number of turtles. This sample size was low for evaluating the effects of five covariates, and therefore, caution is warranted in interpreting these data. We estimated the repeatability as (among individual variance/[among individual variance + residual variance]), using the estimated variance components from the LMM output. For repeatability and for all covariates, we evaluated significance using parametric bootstrapping to calculate 95% confidence intervals, as implemented in the package “rptR” for repeatability estimates (Nakagawa and Schielzeth 2010) and the confint function in the package “lme4” for fixed effects. We also report the coefficient of variation for between-individual variance (CVB) in behavior as a standardized measure of the degree of variation among individuals, following recent recommendations (Holtmann et al. 2017).
We next evaluated relationships between mean emergence and movement latencies and the ecological variables of interest: body temperature, injury scores, and movement rates. For body temperature, we anticipated that the mean body temperatures of turtles would be negatively correlated with emergence and movement latencies. We calculated the daily mean temperatures for each individual turtle from their attached iButtons, excluding the first and last days for which we did not have a full 24 h of data. We subtracted from each daily mean the mean reference temperatures for the corresponding days that were acquired from an iButton placed in an arbitrary, shaded location on the forest floor. Subtracting the temperature at a reference site allowed us to account for variation in body temperature that is due to day-to-day variation in the ambient temperature (Feaga and Haas 2015). Generally, turtle body temperatures were higher than reference temperatures, suggesting that the turtles non-randomly favored sunnier, warmer areas than our reference site. We calculated each turtle’s mean temperature difference from the reference across all days it was monitored (ranging from 11 to 35 days) and used this as a dependent variable in a linear regression with log-transformed mean emergence and movement latencies as independent variables. The correlation between mean emergence and movement latencies was modest (r = 0.42; see “Results” section) resulting in minimal variance inflation due to collinearity. For this analysis, we also included the year of the study (2015 or 2016), as the studies in the 2 years occurred at slightly different times of year and 2016 was characterized by lower cloud cover and precipitation than 2015. For injury scores, we predicted a negative relationship between the extent of injuries and emergence and movement latencies. We used linear regression to assess the association between injury score and log-transformed mean emergence and movement latencies, and we included age class as an additional covariate in this analysis, as older turtles would be expected to have accumulated more damage to their shells.
For movement rates, we expected mean rates of movement to be negatively associated with emergence and movement latencies. We measured the distance moved between coordinates for consecutive recaptures and divided by the days elapsed between recaptures to yield meters moved per day. The values obtained were within the range previously reported for this species (40 ± 50 m/day [mean ± 1 standard deviation]; Strang 1983). We also preliminarily evaluated minimum convex polygon (MCP) estimates of home range size, but MCP and other home range estimators are very sensitive to the number of locations recorded (Seaman et al. 1999; Harless et al. 2010), which was very low in this study. The results of analyses of home range and movement rates were nonetheless qualitatively similar. Distances between recaptures were measured using the spDistsN1 function in the “sp” package in R (Pebesma and Bivand 2005). We used the mean movement rate for each individual turtle, but excluded four individual turtles for which we had data for fewer than five relocations (and hence had four or fewer estimates of meters moved per day). We then used linear regression to evaluate the association between mean movement rates (dependent variable) and log-transformed mean emergence and movement latencies (independent variables). We also included nesting status as a covariate, as we expected nesting females to move at higher rates than males and non-nesting females.
Data availability
The datasets generated and analyzed in this study are available through Figshare (https://doi.org/10.6084/m9.figshare.5631979.v1).
Results
Characteristics of the sampled turtles are described in Table 1. We found evidence that emergence latency and movement latency were consistent personality traits. Emergence latency was highly repeatable within individuals, with an estimated r = 0.73 (95% CI [0.61, 0.82]; CVB = 1.52; Fig. 2a). Emergence latency repeatability remained high after accounting for variables associated with assay conditions (r = 0.75, 95% CI [0.61, 0.83]). The only assay condition that significantly affected emergence latency was time of day, with emergence time being shorter later in the day (Table 2). After accounting for individual phenotypic differences, emergence latency was still similarly repeatable (r = 0.75, 95% CI [0.62, 0.84]), and emergence times were not explained by any of the included phenotypic characteristics (Table 2); the effects of particular phenotypic characteristics should be interpreted with caution, however, given the relatively high number of model parameters for the sample size at the level of individual turtles.

Individual box turtle a head emergence latency and b movement latency. Values are presented as means for each turtle (large gray circle), with individual observation values (small open circles) located above and below means to demonstrate intraindividual variation. A small amount of random noise has been added to individual observation values to reduce overlap. Individual turtles have been sorted from left to right by mean head emergence latency. Axes for latencies are logarithmic
Similarly, the latency to begin moving after emerging from the shell was repeatable within individuals, but less so than emergence latency (r = 0.43, 95% CI [0.25, 0.58]; CVB = 0.75; Fig. 2b). As before repeatability was affected little by accounting for assay conditions (r = 0.41, 95% CI [0.23, 0.56]), none of which were significantly associated with movement latency, though there was a tendency for shorter movement latencies later in the day and in turtles that were exposed when they were located (Table 3). Repeatability was also similar after adjusting for individual phenotypic differences (r = 0.40, 95% CI [0.21, 0.57]). None of these covariates were significantly associated with movement latency.
We evaluated the association between head emergence latency and movement latency to determine the extent to which they were independent traits. Head emergence latencies and movement latencies were significantly but modestly correlated, both at the level of individual observations (r = 0.42, t 199 = 6.61, p < 0.0001) and at the level of mean values for individual turtles (r = 0.43, t 30 = 2.59, p = 0.01).
We next tested for the effects of emergence and movement latencies on temperature, injury scores, and movement rates. After accounting for higher temperatures experienced by box turtles in 2016 relative to reference iButtons than in 2015 (β = 1.31, t 23 = 5.82, p < 0.0001), turtles with longer mean head emergence latencies maintained relatively cooler temperatures (β = − 0.16, t 23 = 2.82, p = 0.01; Fig. 3). There was no association between mean movement latency and temperature (β = 0.10, t 23 = 1.17, p = 0.26). Mean head emergence latency was not associated with injury scores (β = 0.42, t 28 = 1.34, p = 0.19), but higher mean movement latencies tended to be associated with reduced injury scores (β = − 0.91, t 28 = 1.86, p = 0.07; Fig. 4). Older age classes of turtles tended to have higher injury scores (β = 1.99, t 28 = 2.09, p = 0.046). Average movement rates were not associated with mean head emergence latency (β = − 0.57, t 25 = 0.42, p = 0.68) or movement latency (β = 1.72, t 25 = 0.81, p = 0.43). However, nesting female turtles moved significantly more per day (β = 21.80, t 25 = 3.01, p = 0.006).

The relationship between relative temperature (mean temperature in °C of shell-affixed data logger minus mean temperature of reference data logger) and mean head emergence latencies for individual turtles. Circles indicate data from 2015, and triangles represent data from 2016. The line indicates the relationship between relative temperature and head emergence latency, adjusted for the effects of movement latency and year in a multiple regression model

The relationship between shell injury scores and mean movement latencies for individual turtles. Movement latency is presented on a logarithmic axis. The relationship tended to be negative (p = 0.07; see “Results” section)
Discussion
Our research supports and extends previous work (e.g., Carter et al. 2010; Goulet et al. 2017b) on ecological correlates of personality in terrestrial ectotherms. We found that eastern box turtles display consistent behavioral differences across time and within individuals, and these behavioral differences are associated in expected ways with thermoregulation and injuries; bolder turtles maintained higher body temperatures but tended (albeit non-significantly) to have more injuries. This suggests a potentially important trade-off between thermoregulation and predation risk that may influence selection on personality in turtles. We explore each of these findings in greater depth below.
Individual turtles consistently differed from each other in their latency to emerge from the shell and to begin moving following a standardized disturbance, suggestive of the existence of a boldness-shyness axis of variation in personality (Briffa et al. 2008). The breadth of personality differences was dramatic (Table 1; Fig. 2), with some individuals reliably emerging immediately from their shells after a presumably stressful interaction with humans while others remained in the shell for several minutes. Notably, the mean emergence times for individuals are strongly bimodal, with similar numbers of individuals having average emergence times of less than 7 s and more than 58 s, with no intermediate behavioral phenotypes. This bimodal distribution of personality types has been observed in other species (Pruitt et al. 2008; Bergeron et al. 2013) and is consistent with models of disruptive selection on personality (Wolf et al. 2007). Moreover, testing conditions (such as environmental factors or test number) and other features of individual phenotypes (such as sex and age) minimally influenced these behaviors, and the repeatability of these behaviors was not attributable to these other potential generators of inter-individual behavioral differences (López et al. 2005; Näslund and Johnsson 2016). When compared to a meta-analysis of repeatability estimates of animal behavior (Bell et al. 2009), our data show relatively high repeatability for head emergence latency and moderate repeatability for movement latency, particularly in comparison to other herpetofauna. Such behavioral consistency that is not dependent on the environment or individual state indicates strong evidence that boldness (as measured by our assay) is a personality trait in eastern box turtles. However, future work could validate the presence of an underlying boldness-shyness axis of behavioral variation in box turtles by performing alternative behavioral tests of responses to risk and demonstrating inter-assay consistency among these measures (Beckmann and Biro 2013).
Boldness appeared to be positively associated with thermoregulation. As ectotherms, turtles must access thermal resources (i.e., basking sites) to regulate body temperatures and, consequently, physiological processes such as immune function (Zimmerman et al. 2017) and digestion (Harlow et al. 1976). As inhabitants of forests, which may offer limited access to sunlight, eastern box turtles may be especially constrained in their ability to maintain high body temperatures. Exposure to and movement to basking sites are likely to present a greater risk of predation, and therefore, we anticipated that bolder individuals would maintain higher body temperatures than more risk-averse conspecifics. We indeed found that bolder turtles are warmer turtles—average body temperatures relative to a stationary reference point were higher in turtles with shorter latencies for head emergence (but not movement). This finding joins a growing body of evidence of individual variation in thermoregulatory behavior. Within reptiles, Goulet et al. (2017a, b) described “thermal types” in the delicate skink (Lampropholis delicata), finding that individuals varied consistently in thermoregulation, and Carter et al. (2010) found that bolder male Namibian agama lizards (Agama planiceps) spent more time basking but did not document or discuss the thermoregulatory consequences of these behavioral differences. We interpret the relationship between boldness and thermoregulation in turtles as integration of personality and physiology (Goulet et al. 2017b), with bolder behaviors favoring the maintenance of higher body temperatures (and therefore higher metabolic rate) for greater performance of escape behavior (Dodd 2001, p. 140; Hertz et al. 1982) and/or for enhanced rates of growth and reproduction to compensate for greater mortality (Smith and Blumstein 2008). However, it is possible that differences in body temperature causally precede behavioral differences. Under this assumption, turtles that tend to be warmer as a consequence of idiosyncratic differences in movement behavior or home range features may exhibit bolder behaviors. This explanation is less likely as neither shell temperature nor air temperature at the time of testing was correlated with latency to emerge or move, and therefore, boldness is generally stable across the range of temperatures experienced in this study. It is possible, however, that long-term exposure to alternative thermal regimes could support the expression of differing personality types (Stahlschmidt et al. 2016). This could be evaluated by comparing behavior among populations of turtles that experience different average temperatures as a result of climate or by rearing turtles in controlled environments and evaluating behavioral variation.
We also found a tendency for greater injuries to the shell in bolder turtles. As boldness reflects the propensity towards tolerating versus avoiding risk, animals with bolder personalities are generally hypothesized to experience more encounters with and/or successful attacks by predators (Stamps 2007), and this is often supported (Dugatkin 1992; Hulthén et al. 2017; but see, Langkilde and Carlson 2014; Delnat et al. 2017). In the box turtles in this study, thermoregulatory behavior, in particular, presents a plausible mechanistic link between boldness and predatory risk, as described above. Fortunately, turtles often carry a record of their history of attempted predation events in the form of damage to the shell. There was a tendency for turtles with shorter movement latencies to bear more extensive injuries on their shell, similar to the correlation observed between tail loss (presumably by predators) and boldness in lizards (Carter et al. 2010). Given the consistency of this result with our expectations and prior work, we suggest it is likely that this represents a real pattern but should nonetheless be considered cautiously. We propose that turtles that move more quickly after an attack are more likely to be attacked again, whereas shyer turtles remain motionless longer and may not begin moving until the predator has left the area, preventing a second attack. Alternatively, shorter movement latency in our assays may indicate a greater propensity for movement in general, which could make bolder turtles more likely to attract the attention of a predator. It is also possible that the history of predation events and injuries determines behavior, with attacked individuals becoming desensitized to predator encounters (Brown et al. 2007; Thaker et al. 2010), generating a correlation between boldness and injuries. Determining whether turtles were emboldened by their prior experiences of predators, or bolder turtles are more likely to encounter predators, could be addressed by continually monitoring behavior and injuries in free-ranging turtles to determine whether boldness changes following additional predation events or is instead predictive of the occurrence of new injuries.
The tendency for boldness to positively correlate with injuries makes the continued persistence of bold turtles in the population to be puzzling, especially considering the presumed effectiveness of the defensive behavior they are minimally utilizing (hiding in the shell). The plastron (a ventral portion of the shell) of box turtles is hinged, enabling them to completely enclose the retracted head and limbs, forming a defensive barrier that is nearly impenetrable to most common predators (Dodd 2001). Nonetheless, we found that a large proportion of the turtles we observed extended their head and/or limbs very quickly after handling by a human, and many remained unenclosed in the shell even while being handled and having blood drawn (B.E. Carlson, personal observations). The failure of some individuals to employ what is apparently a highly effective defense during encounters with predators has been previously observed and credited for the occasional successful predation event by medium-sized carnivores (e.g., raccoons, opossums, and coyotes) upon adult box turtles (Dodd 2001, p. 140), though many of the injuries we observed could have been inflicted when the turtles were younger and more easily damaged. While intraspecific variation in boldness is widespread, it remains striking that such apparently maladaptive behavior is maintained. Behavioral syndromes are present when behavioral traits are correlated across contexts, demonstrating limited plasticity (Sih et al. 2004). This can generate trade-offs in which a behavioral trait is maladaptive in certain circumstances, but either the same or a linked behavior provides a compensatory benefit in other situations (Duckworth 2006). In box turtles, we found evidence of thermoregulatory benefits for bold turtles, and it is possible that the fitness effects of this or other, unmeasured characteristics (e.g., fecundity) of bold turtles compensate for the increased risk of predation. Alternatively, the lack of concealment in the shell in some individuals could be because the threat perceived by human handling and subsequent confinement was not strong enough to elicit an antipredator response, though a higher risk situation would have resulted in the expression of appropriate behaviors (Quinn and Cresswell 2005). It would be valuable to conduct similar behavioral assays after exposure to more realistic simulated predation events. Finally, it is also possible that bold turtles simply exhibit alternative, but effective, antipredator behaviors. Fleeing, especially to cover, may be a deterrent to some predators, as could be biting and defecating, behaviors the turtles in this study occasionally performed. Moreover, the shell may be utilized on a moment-to-moment basis rather than for prolonged periods of hiding, with bold turtles retreating into and emerging from the shell repeatedly at appropriate times during the interaction with the predator. Addressing this possibility would entail a more comprehensive assessment of defensive behaviors.
We hypothesized that turtles who exhibited higher levels of boldness would also exhibit higher rates of movement because frequent relocation would be needed to track shifting patches of sunlight and because movement could constitute a risky behavior and, thus, ought to be correlated with boldness. Moreover, boldness and general activity levels are often correlated in other animals, forming a behavioral syndrome (Wilson and Godin 2009), and larger home ranges have been previously documented in bolder individuals of other species (e.g., Spiegel et al. 2017). This hypothesis was not supported by our data. This may indicate that movement behavior is unrelated to boldness, or bold turtles could actually be more active and mobile but restrict these movements to similar home range areas as shy turtles. However, our ability to draw conclusions from these data is limited by how we quantified the movements of the turtles. By measuring the linear distance between consecutive coordinates collected several days apart, we failed to account for any non-linear movements, which can lead to dramatically underestimated path lengths (Rowcliffe et al. 2012). Future efforts to evaluate the relationship between personality and movement behavior should employ either more frequent relocations of turtles by telemetry or GPS loggers to record positional data at regular intervals (Hebblewhite and Haydon 2010).
Personality in terrestrial ectotherms, namely non-avian reptiles and adult amphibians, has not been examined as extensively as it has in other vertebrates (but see: Brodie and Russell 1999; López et al. 2005; Stapley 2006; Cooper 2009; Carter et al. 2010, 2016; Mafli et al. 2011; Wilson and Krause 2012; Citadini and Navas 2013; González-Bernal et al. 2014; Kuo et al. 2015; Stahlschmidt et al. 2016; Riley et al. 2017; Šimková et al. 2017; Siviter et al. 2017). Our study adds to the nascent understanding of behavioral differences in this group of animals. Specifically, we suggest that the emergence behavior of box turtles in response to the stress-inducing experience of human handling and confinement is highly variable among individuals and stable within individuals, and such variation in boldness could be maintained by a trade-off between thermoregulation and predation risk. The concept of a trade-off between growth and mortality is well-established as a potential contributor to the persistence of interindividual variation in boldness, but this present study demonstrates how this trade-off may be uniquely manifested in terrestrial ectotherms faced with a thermally challenging environment. Continued work in reptiles and amphibians may reveal whether this is a general pattern within these taxa and thus expand our understanding of the evolution of animal personality.
References
Adriaenssens B, Pauliny A, Blomqvist D, Johnsson JI (2016) Telomere length covaries with personality in wild brown trout. Physiol Behav 165:217–222. https://doi.org/10.1016/j.physbeh.2016.07.005
Archard GA, Braithwaite VA (2010) The importance of wild populations in studies of animal temperament. J Zool 281:149–160
Ballew NG, Mittelbach GG, Scribner KT (2017) Fitness consequences of boldness in juvenile and adult largemouth bass. Am Nat 189(4):396–406. https://doi.org/10.1086/690909
Basson CH, Levy O, Angilletta MJ, Clusella-Trullas S (2017) Lizards paid a greater opportunity cost to thermoregulate in a less heterogeneous environment. Funct Ecol 31(4):856–865. https://doi.org/10.1111/1365-2435.12795
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48
Beckmann C, Biro PA (2013) On the validity of a single (boldness) assay in personality research. Ethology 119:937–947
Bell AM, Hankison SJ, Laskowski KL (2009) The repeatability of behaviour: a meta-analysis. Anim Behav 77(4):771–783. https://doi.org/10.1016/j.anbehav.2008.12.022
Bergeron P, Montiglio P-O, Réale D, Humphries MM, Gimenez O, Garant D (2013) Disruptive viability selection on adult exploratory behaviour in eastern chipmunks. J Evol Biol 26(4):766–774. https://doi.org/10.1111/jeb.12081
Bergmüller R, Taborsky M (2010) Animal personality due to social niche specialisation. Trends Ecol Evol 25(9):504–511. https://doi.org/10.1016/j.tree.2010.06.012
Bernstein NP, Black RW (2005) Thermal environment of overwintering ornate box turtles, Terrapene ornata ornata, in Iowa. Am Midl Nat 153(2):370–377. https://doi.org/10.1674/0003-0031(2005)153[0370:TEOOOB]2.0.CO;2
Binder TR, Wilson ADM, Wilson SM, Suski CD, Godin J-GJ, Cooke SJ (2016) Is there a pace-of-life syndrome linking boldness and metabolic capacity for locomotion in bluegill sunfish? Anim Behav 121:175–183. https://doi.org/10.1016/j.anbehav.2016.09.006
Biro PA, Dingemanse NJ (2009) Sampling bias resulting from animal personality. Evolution 24:66–67
Biro PA, Stamps JA (2008) Are animal personality traits linked to life-history productivity? Trends Ecol Evol 23(7):361–368. https://doi.org/10.1016/j.tree.2008.04.003
Biro PA, Adriaenssens B, Sampson P (2014) Individual and sex-specific differences in intrinsic growth rate covary with consistent individual differences in behaviour. J Anim Ecol 83(5):1186–1195. https://doi.org/10.1111/1365-2656.12210
Boyer N, Réale D, Marmet J, Pisanu B, Chapuis JL (2010) Personality, space use and tick load in an introduced population of Siberian chipmunks Tamias sibiricus. J Anim Ecol 79(3):538–547. https://doi.org/10.1111/j.1365-2656.2010.01659.x
Bremner-Harrison S, Prodohl PA, Elwood RW (2004) Behavioural trait assessment as a release criterion: boldness predicts early death in a reintroduction programme of captive-bred swift fox (Vulpes velox). Anim Conserv 7(3):313–320. https://doi.org/10.1017/S1367943004001490
Briffa M, Rundle SD, Fryer A (2008) Comparing the strength of behavioural plasticity and consistency across situations: animal personalities in the hermit crab Pagurus bernhardus. Proc R Soc Lond B 275(1640):1305–1311. https://doi.org/10.1098/rspb.2008.0025
Brodie ED III, Russell NH (1999) The consistency of individual differences in behaviour: temperature effects on antipredator behaviour in garter snakes. Anim Behav 57:445–451
Brown C, Burgess F, Braithwaite VA (2007) Heritable and experiential effects on boldness in a tropical poeciliid. Behav Ecol Sociobiol 62(2):237–243. https://doi.org/10.1007/s00265-007-0458-3
Careau V, Thomas D, Humphries MM, Réale D (2008) Energy metabolism and animal personality. Oikos 117(5):641–653. https://doi.org/10.1111/j.0030-1299.2008.16513.x
Carter AJ, Goldizen AW, Tromp SA (2010) Agamas exhibit behavioral syndromes: bolder males bask and feed more but may suffer higher predation. Behav Ecol 21(3):655–661. https://doi.org/10.1093/beheco/arq036
Carter AW, Paitz RT, McGhee KE, Bowden RM (2016) Turtle hatchlings show behavioral types that are robust to developmental manipulations. Physiol Behav 155:46–55. https://doi.org/10.1016/j.physbeh.2015.11.034
Citadini JM, Navas CA (2013) Inter-individual variation and temperature-dependent antipredator behavior in the snake Tomodon dorsatus (Dipsadidae). Behav Process 97:11–17. https://doi.org/10.1016/j.beproc.2013.03.008
Cooper WE (2009) Variation in escape behavior among individuals of the striped plateau lizard Sceloporus virgatus may reflect differences in boldness. J Herpetol 43(3):495–502. https://doi.org/10.1670/08-197R1.1
Dall SRX, Houston AI, McNamara JM (2004) The behavioural ecology of personality: consistent individual differences from an adaptive perspective. Ecol Lett 7(8):734–739. https://doi.org/10.1111/j.1461-0248.2004.00618.x
Debeffe L, Morellet N, Bonnot N, et al (2014) The link between behavioural type and natal dispersal propensity reveals a dispersal syndrome in a large herbivore. Proc R Soc B 281(1790):20140873. https://doi.org/10.1098/rspb.2014.0873
DeGregorio BA, Tuberville TD, Kennamer RA, Harris BB, Brisbin IL Jr (2017) Spring emergence of eastern box turtles (Terrapene carolina): influences of individual variation and scale of temperature correlates. Can J Zool 95(1):23–30. https://doi.org/10.1139/cjz-2016-0149
Delnat V, Debecker S, Stoks R (2017) Integrating trait multidimensionality, predation and autotomy to explain the maintenance of boldness. Anim Behav 130:97–105. https://doi.org/10.1016/j.anbehav.2017.06.014
do Amaral JPS, Marvin G, Hutchison VH (2002) Thermoregulation in the box turtles Terrapene carolina and Terrapene ornata. Can J Zool 80(5):934–943. https://doi.org/10.1139/z02-070
Dodd CK Jr (2001) North American box turtles: a natural history. University of Oklahoma Press, Norman, OK
Dodd CK Jr, Franz R, Johnson SA (1997) Shell injuries and anomalies in an insular population of Florida box turtles (Terrapene carolina bauri). Herpetol. Nat Hist 51:66–72
Duckworth RA (2006) Behavioral correlations across breeding contexts provide a mechanism for a cost of aggression. Behav Ecol 17(6):1011–1019. https://doi.org/10.1093/beheco/arl035
Dugatkin LA (1992) Tendency to inspect predators predicts mortality risk in the guppy (Poecilia reticulata). Behav Ecol 3(2):124–127. https://doi.org/10.1093/beheco/3.2.124
Ernst C, Hershey M, Barbour R (1974) A new coding system for hard-shelled turtles. Trans Kentucky Acad Sci 35:27–28
Feaga JB, Haas CA (2015) Seasonal thermal ecology of bog turtles (Glyptemys muhlenbergii) in southwestern Virginia. J Herpetol 49(2):264–275. https://doi.org/10.1670/13-147
Gatten RE (1974) Effects of temperature and activity on aerobic and anaerobic metabolism and heart rate in the turtles Pseudemys scripta and Terrapene ornata. Comp Biochem Physiol A 48(4):619–648. https://doi.org/10.1016/0300-9629(74)90606-9
González-Bernal E, Brown GP, Shine R (2014) Invasive cane toads: social facilitation depends upon an individual’s personality. PLoS One 9(7):e102880. https://doi.org/10.1371/journal.pone.0102880
Goulet CT, Thompson MB, Chapple DG (2017a) Repeatability and correlation of physiological traits: do ectotherms have a “thermal type”? Ecol Evol 7(2):710–719. https://doi.org/10.1002/ece3.2632
Goulet CT, Thompson MB, Michelangeli M, Wong BBM, Chapple DG (2017b) Thermal physiology: a new dimension of the pace-of-life syndrome. J Anim Ecol 86(5):1269–1280. https://doi.org/10.1111/1365-2656.12718
Guenther A, Trillmich F (2015) Within-litter differences in personality and physiology relate to size differences among siblings in cavies. Physiol Behav 145:22–28. https://doi.org/10.1016/j.physbeh.2015.03.026
Harless ML, Walde AD, Delaney DK, Pater LL, Hayes WK (2010) Sampling considerations for improving home range estimates of desert tortoises: effects of estimator, sampling regime, and sex. Herpetol Conserv Biol 5:374–387
Harlow HJ, Hillman SS, Hoffman M (1976) The effect of temperature on digestive efficiency in the herbivorous lizard, Dipsosaurus dorsalis. J Comp Physiol B 111:1–6
Hebblewhite M, Haydon DT (2010) Distinguishing technology from biology: a critical review of the use of GPS telemetry data in ecology. Philos Trans R Soc B 365(1550):2303–2312. https://doi.org/10.1098/rstb.2010.0087
Hertz PE, Huey RB, Nevo E (1982) Fight versus flight: body temperature influences defensive responses of lizards. Anim Behav 30(3):676–679. https://doi.org/10.1016/S0003-3472(82)80137-1
Hirsch PE, Thorlacius M, Brodin T, Burkhardt-Holm P (2017) An approach to incorporate individual personality in modeling fish dispersal across in-stream barriers. Ecol Evol 7(2):720–732. https://doi.org/10.1002/ece3.2629
Holtmann B, Lagisz M, Nakagawa S (2017) Metabolic rates, and not hormone levels, are a likely mediator of between-individual differences in behaviour: a meta-analysis. Funct Ecol 31(3):685–696. https://doi.org/10.1111/1365-2435.12779
Huey RB, Slatkin M (1976) Cost and benefits of lizard thermoregulation. Q Rev Biol 51(3):363–384. https://doi.org/10.1086/409470
Hulthén K, Chapman BB, Nilsson PA, Hansson L-A, Skov C, Brodersen J, Vinterstare J, Brönmark C (2017) A predation cost to bold fish in the wild. Sci Rep 7(1):1239. https://doi.org/10.1038/s41598-017-01270-w
Ibáñez A, López P, Martín J (2014) Inter-individual variation in antipredator hiding behavior of Spanish terrapins depends on sex, size, and coloration. Ethology 120(8):742–752. https://doi.org/10.1111/eth.12245
Kanda LL, Louon L, Straley K (2012) Stability in activity and boldness across time and context in captive Siberian dwarf hamsters. Ethology 118(6):518–533. https://doi.org/10.1111/j.1439-0310.2012.02038.x
Kuo C-Y, Irschick DJ, Lailvaux SP (2015) Trait compensation between boldness and the propensity for tail autotomy under different food availabilities in similarly aged brown anole lizards. Funct Ecol 29(3):385–392. https://doi.org/10.1111/1365-2435.12324
Langkilde T, Carlson BE (2014) No evidence of selection by predators on tadpole boldness. Behaviour 151:23–45
López P, Hawlena D, Polo V, Amo L, Martín J (2005) Sources of individual shy–bold variations in antipredator behaviour of male Iberian rock lizards. Anim Behav 69(1):1–9. https://doi.org/10.1016/j.anbehav.2004.05.010
MacKay J, Haskell M (2015) Consistent individual behavioral variation: the difference between temperament, personality and behavioral syndromes. Animals 5(3):455–478. https://doi.org/10.3390/ani5030366
Mafli A, Wakamatsu K, Roulin A (2011) Melanin-based coloration predicts aggressiveness and boldness in captive eastern Hermann’s tortoises. Anim Behav 81(4):859–863. https://doi.org/10.1016/j.anbehav.2011.01.025
Mathot KJ, Nicolaus M, Araya-Ajoy YG, Dingemanse NJ, Kempenaers B (2014) Does metabolic rate predict risk-taking behaviour? A field experiment in a wild passerine bird. Funct Ecol 29:239–249
Mell H, Josserand R, Decencière B, Artacho P, Meylan S, Le Galliard J-F (2016) Do personalities co-vary with metabolic expenditure and glucocorticoid stress response in adult lizards? Behav Ecol Sociobiol 70(6):951–961. https://doi.org/10.1007/s00265-016-2117-z
Myers RE, Hyman J (2016) Differences in measures of boldness even when underlying behavioral syndromes are present in two populations of the song sparrow (Melospiza melodia). J Ethol 34(3):197–206. https://doi.org/10.1007/s10164-016-0465-9
Nakagawa S, Schielzeth H (2010) Repeatability for Gaussian and non-Gaussian data: a practical guide for biologists. Biol Rev 85(4):935–956. https://doi.org/10.1111/j.1469-185X.2010.00141.x
Näslund J, Johnsson JI (2016) State-dependent behavior and alternative behavioral strategies in brown trout (Salmo trutta L.) fry. Behav Ecol Sociobiol 70(12):2111–2125. https://doi.org/10.1007/s00265-016-2215-y
Pebesma EJ, Bivand RS (2005) Classes and methods for spatial data in R. R News 5:9–13
Pruitt J, Riechert S, Jones T (2008) Behavioural syndromes and their fitness consequences in a socially polymorphic spider, Anelosimus studiosus. Anim Behav 76(3):871–879. https://doi.org/10.1016/j.anbehav.2008.05.009
Quinn JL, Cresswell W (2005) Personality, anti-predation behaviour and behavioural plasticity in the chaffinch Fringilla coelebs. Behaviour 142(9):1377–1402. https://doi.org/10.1163/156853905774539391
R Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna https://www.R-project.org/
Riley JL, Noble DWA, Byrne RW, Whiting MJ (2017) Early social environment influences the behaviour of a family-living lizard. R Soc Open Sci 4(5):161082. https://doi.org/10.1098/rsos.161082
Rowcliffe JM, Carbone C, Kays R, Kranstauber B, Jansen PA (2012) Bias in estimating animal travel distance: the effect of sampling frequency. Methods Ecol Evol 3(4):653–662. https://doi.org/10.1111/j.2041-210X.2012.00197.x
Roznik EA, Alford RA (2012) Does waterproofing Thermochron iButton dataloggers influence temperature readings? J Therm Biol 37(4):260–264. https://doi.org/10.1016/j.jtherbio.2012.02.004
Saumure RA, Herman TB, Titman RD (2007) Effects of haying and agricultural practices on a declining species: the North American wood turtle, Glyptemys insculpta. Biol Conserv 135(4):565–575. https://doi.org/10.1016/j.biocon.2006.11.003
Seaman DE, Millspaugh JJ, Kernohan BJ, Brundige GC, Raedeke KJ, Gitzen RA (1999) Effects of sample size on kernel home range estimates. J Wildl Manage 63:739–747
Sih A, Bell AM (2008) Insights for behavioral ecology from behavioral syndromes. Adv Stud Behav 38:227–281. https://doi.org/10.1016/S0065-3454(08)00005-3
Sih A, Bell A, Johnson JC (2004) Behavioral syndromes: an ecological and evolutionary overview. Trends Ecol Evol 19(7):372–378. https://doi.org/10.1016/j.tree.2004.04.009
Sih A, Cote J, Evans M, Fogarty S, Pruitt J (2012) Ecological implications of behavioural syndromes. Ecol Lett 15(3):278–289. https://doi.org/10.1111/j.1461-0248.2011.01731.x
Šimková O, Frýdlová P, Žampachová B, Frynta D, Landová E (2017) Development of behavioural profile in the northern common boa (Boa imperator): repeatable independent traits or personality? PLoS One 12(5):e0177911. https://doi.org/10.1371/journal.pone.0177911
Siviter H, Deeming DC, Rosenberger J, Burman OHP, Moszuti SA, Wilkinson A (2017) The impact of egg incubation temperature on the personality of oviparous reptiles. Anim Cogn 20(1):109–116. https://doi.org/10.1007/s10071-016-1030-1
Smith BR, Blumstein DT (2008) Fitness consequences of personality: a meta-analysis. Behav Ecol 19:448–455
Spiegel O, Leu ST, Bull CM, Sih A (2017) What’s your move? Movement as a link between personality and spatial dynamics in animal populations. Ecol Lett 20(1):3–18. https://doi.org/10.1111/ele.12708
Stahlschmidt ZR, Holcomb LM, Luoma RL (2016) Context-dependent effects of complex environments on behavioral plasticity. Behav Ecol 27(1):237–244. https://doi.org/10.1093/beheco/arv147
Stamps JA (2007) Growth-mortality tradeoffs and “personality traits” in animals. Ecol Lett 10(5):355–363. https://doi.org/10.1111/j.1461-0248.2007.01034.x
Stapley J (2006) Individual variation in preferred body temperature covaries with social behaviours and colour in male lizards. J Therm Biol 31(4):362–369. https://doi.org/10.1016/j.jtherbio.2006.01.008
Strang CA (1983) Spatial and temporal activity patterns in two terrestrial turtles. J Herpetol 17(1):43–47. https://doi.org/10.2307/1563779
Thaker M, Vanak AT, Lima SL, Hews DK (2010) Stress and aversive learning in a wild vertebrate: the role of corticosterone in mediating escape from a novel stressor. Am Nat 175(1):50–60. https://doi.org/10.1086/648558
Thomas P, Herrel A, Hardy I, Aujard F, Pouydebat E (2016) Exploration behavior and morphology are correlated in captive gray mouse lemurs (Microcebus murinus). Int J Primatol 37(3):405–415. https://doi.org/10.1007/s10764-016-9908-y
Williams EC Jr, Parker WS (1987) A long-term study of a box turtle (Terrapene carolina) population at Allee Memorial Woods, Indiana, with emphasis on survivorship. Herpetologica 43:328–335
Wilson ADM, Godin J-GJ (2009) Boldness and behavioral syndromes in the bluegill sunfish, Lepomis macrochirus. Behav Ecol 20(2):231–237. https://doi.org/10.1093/beheco/arp018
Wilson ADM, Krause J (2012) Personality and metamorphosis: is behavioral variation consistent across ontogenetic niche shifts? Behav Ecol 23(6):1316–1323. https://doi.org/10.1093/beheco/ars123
Wilson DS, Clark AB, Coleman K, Dearstyne T (1994) Shyness and boldness in humans and other animals. Trends Ecol Evol 9(11):442–446. https://doi.org/10.1016/0169-5347(94)90134-1
Wilson DS, Tracy CR, Tracy CR (2003) Estimating age of turtles from growth rings: a critical evaluation of the technique. Herpetologica 59(2):178–194. https://doi.org/10.1655/0018-0831(2003)059[0178:EAOTFG]2.0.CO;2
Wolf M, McNamara JM (2012) On the evolution of personalities via frequency-dependent selection. Am Nat 179(6):679–692. https://doi.org/10.1086/665656
Wolf M, Weissing FJ (2012) Animal personalities: consequences for ecology and evolution. Trends Ecol Evol 27(8):452–461. https://doi.org/10.1016/j.tree.2012.05.001
Wolf M, van Doorn GS, Leimar O, Weissing FJ (2007) Life-history trade-offs favour the evolution of animal personalities. Nature 447(7144):581–584. https://doi.org/10.1038/nature05835
Zimmerman LM, Carter AW, Bowden RM, Vogel LA (2017) Immunocompetence in a long-lived ectothermic vertebrate is temperature dependent but shows no decline in older adults. Funct Ecol 31(7):1383–1389. https://doi.org/10.1111/1365-2435.12867
Acknowledgements
B. Boles, T. Flock, S. Khoo, and W. Robinson provided essential assistance with fieldwork. The feedback of two anonymous reviewers greatly improved this manuscript. Financial support was provided by Wabash College, the Norman E. Treves Fund, and the Robert O. Petty Fund.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
All applicable international, national, and institutional guidelines for the care and use of animals were followed. All procedures performed were in accordance with the ethical standards of the institution at which the studies were conducted. Approval was provided by the Wabash College Institutional Animal Care and Use Committee and by the Indiana Department of Natural Resources (Scientific Purposes License 15-136 and 16-154).
Additional information
Communicated by T. Madsen
Rights and permissions
About this article

Cite this article
Kashon, E.A.F., Carlson, B.E. Consistently bolder turtles maintain higher body temperatures in the field but may experience greater predation risk. Behav Ecol Sociobiol 72, 9 (2018). https://doi.org/10.1007/s00265-017-2428-8
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-017-2428-8