
Abstract
In groups with multiple males, direct mate competition may select for the evolution of dominance hierarchies that sort males into a queue for access to fertile females. The priority-of-access (PoA) model proposed by Altmann in 1962 makes explicit predictions about the resulting paternity distribution based on an interaction between male dominance rank and the overlap of female receptive phases. Here, we investigated whether the logic of the PoA model predicted the distribution of paternity across ranks in a seasonal breeder with high reproductive synchrony over six consecutive mating seasons. We studied 18 males that resided in a group of wild Assamese macaques (Macaca assamensis) in their natural habitat at Phu Khieo Wildlife Sanctuary, Thailand, between 2006 and 2011 with 5 to 13 conceptions per season. We assessed whether mate guarding increased paternity success, described “short-term” deviations from predicted paternity distribution, and examined how these are related to the number of competitors and fertile females. We determined genetic paternity of 43 (93 %) offspring born into the study group and found reproductive skew to be relatively low with 29 % alpha male paternity in accordance with the high degree of female reproductive synchrony observed. Short-term deviations from expected paternity distribution over ranks were not explained by the number of resident males or the number of conceiving females or their interaction. Within the limits of this study, these results suggest that even if males cannot discern female fertile phases, if reproduction is seasonal, and if reproductive synchrony is high, males may also compete directly over access to females.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sexual selection theory posits that the sex that is in oversupply concerning readiness to breed or the sex that has the higher potential reproductive rate will compete for access to the other sex (Clutton-Brock and Parker 1992; Trivers 1972). Internal fertilization, gestation, and lactation, all slowing down female reproduction, make males the competing sex in most polygynous mammals (Clutton-Brock and Vincent 1991; Clutton-Brock and Parker 1992). In gregarious species, males may try to expel rivals from bisexual groups and form harems (Emlen and Oring 1977). Where exclusion is not economically feasible, males are predicted to exclude competitors from access to fertile females present in the group at a given point in time. Direct competition among individually recognizable conspecifics promotes the formation of dominance hierarchies that reduce the frequency and intensity of fighting (Willisch and Neuhaus 2010; Franz et al. 2011). Where hierarchies evolved, like in most primates, male reproductive success is generally predicted by dominance rank, but the degree of inequality, i.e., male reproductive skew, varies widely across time, groups, and species (Cowlishaw and Dunbar 1991; Ellis 1995; Alberts 2012; Majolo et al. 2012).
The priority-of-access (PoA) model originally aimed at investigating the consequences of extended periods of sexual receptivity in anthropoid primates (Altmann 1962). It proposes that variation in reproductive skew is driven by variation in defendability of receptive females living in a group (Altmann 1962) and provides explicit predictions for the distribution of reproductive success across male residents depending on the number of females in the group and the temporal overlap of their receptive phases. Assuming that a male can only monopolize access to one female at a time, the alpha male is allotted all matings/fertilizations in times when only one female is receptive; the alpha and beta males should receive half the matings/fertilizations each when two females are receptive simultaneously; the alpha, beta, and gamma males should receive each a third when three females are available; and so on. Each male has an access to a fertile female only when the number of synchronously available females is equal to or exceeds his ordinal dominance rank.
Explicitly predicted mating and paternity distributions for a given monopolizability situation have been shown to overestimate or underestimate mating and paternity skew and particularly the alpha male’s share in primates and other mammals (e.g., Engh et al. 2002; Alberts et al. 2003, 2006; Boesch et al. 2006; Dubuc et al. 2011; Young et al. 2013). Several factors have been proposed to explain the residual variation: female group size, the number of competitors, male relatedness, leveling coalitions, female choice, and energetic constraints on males (Alberts et al. 2003; Port and Kappeler 2010; Bissonnette et al. 2011; Alberts 2012; Young et al. 2013). Negative deviation from PoA-predicted skew may increase as the number of females in the group increases. This factor is often investigated, but it is unclear whether it creates an independent effect or is a covariate of the number of males or costs of mate guarding (Alberts et al. 2003; Charpentier et al. 2005; Boesch et al. 2006). Male monopolization potential may decrease with increasing numbers of competitors (Alberts et al. 2003; Kutsukake and Nunn 2006). Reduced skew may be predicted by male relatedness if more closely related males are more tolerant around each other in a competitive situation (Charpentier et al. 2005; but see Widdig et al. 2004; Kutsukake and Nunn 2006). Alternatively, increased relatedness theoretically may favor high reproductive skew because subordinates gain inclusive fitness benefits from related dominant individuals (Clutton-Brock 1998; Johnstone 2000; Port and Kappeler 2010). Males may also form coalitions against males to disrupt temporal mating monopolies and to reallocate matings to less successful males which, in effect, level the mating skew (Noë and Sluijter 1990; Kuester and Paul 1992; Alberts et al. 2003; Pandit and van Schaik 2003; Bissonnette et al. 2011; Young et al. 2013). Male mate-guarding activity may be energetically constrained, because of increased energy expenditure and/or decreased energy intake during temporal mating monopolies, which has been proposed to be especially problematic in nonseasonal breeders that have little time to recuperate between mate-guarding episodes (Alberts et al. 1996; Engelhardt et al. 2006; but see Henzi et al. 2010). Apart from these factors that determine how males can and do monopolize access to females, females themselves may play a crucial role, and their precopulatory and postcopulatory mate choice may affect males’ ability to live up to the expectations of the PoA model (Bissonnette et al. 2011; Dubuc et al. 2011; Young et al. 2013). On a more methodological level, variation in males’ inability to assess female fertility may also explain published results on deviation from PoA if not implemented into how the PoA expectations are calculated (Henzi et al. 2010).
Here, we provide data on wild male Assamese macaques (Macaca assamensis) that are unusual among cercopithecines, because during the 4-month mating season, females do not show behavioral estrus (periodic phases of high proceptivity and receptivity, Nadler 1994) or typical fertility indicators (exaggerated sexual swellings, variation in skin coloration, or female behavior indicative of fertility, Fürtbauer et al. 2010, 2011). Male-mating frequency with females does not change from nonfertile to fertile preconception periods or within a conception cycle from 5 days prior to five fertile days to 5 days after, indicating that ovulation is concealed from males (Fürtbauer et al. 2011). As a consequence, female reproductive synchrony (as it can be assessed by males) is unusually high. Nevertheless, males engage in mate guarding with short episodes lasting 1.5 days and long episodes (>1 week) lasting 19 days on average (Schülke et al. 2014). Male mate guarding very closely follows the pattern predicted by the PoA model, i.e., the rank of the most subordinate guard equaled the number of simultaneously guarded females on the vast majority of days, and only the highest-ranking males (ranks 1–4) engage in long consortships (>1 week, Ostner et al. 2011). Thus, despite high female reproductive synchrony, male competition has a significant direct component. However, males do never guard one female throughout the mating season, and males may fail to guard females during the fertile phase of their ovarian cycle (Fürtbauer et al. 2011). Thus, male competition does not only take the direct form of close mate guarding but also other forms of contest competition and sperm competition, and female choice may play a role in determining paternity in Assamese macaques as well.
The PoA model is, strictly speaking, a model of male mate-guarding activity and monopolization. Many previous studies, e.g., on chimpanzees (Pan troglodytes) where other tactics than monopolization are evident (Stumpf and Boesch 2006; Duffy et al. 2007), did not investigate the predictive power of the PoA model in this strict sense. These studies nevertheless tested the basic logic of the PoA, i.e., that rank determines mating or reproductive success and that rank-related reproductive skew is negatively affected by increasing female reproductive synchrony (Boesch et al. 2006; Kutsukake and Nunn 2006; Ostner et al. 2008a, Gogarten and Koenig 2012). Implicitly, these studies assumed that if males do not mate guard all females throughout their entire receptive phases, rank and reproductive synchrony may still determine reproductive success as a result of contest competition. In this situation, contest effects may result from the ability to interfere in others’ mating being rank related or from subordinate males avoiding aggression by refraining from mating in sight of dominants.
Here, we aim at investigating the effect of dominance rank and female receptive synchrony on the paternity distribution across ranks based on genetic paternity data for 93 % of the 46 infants born into one group of wild Assamese macaques across a 6-year study period. We acknowledge that our analyses ignore the role of precopulatory and postcopulatory female choices and provide a discussion of these mechanisms. We investigate whether male mate-guarding activity was associated with increased paternity success, and we complement our main test of observed paternity distribution against the expectation from the PoA model with an investigation of the “short-term” deviation from PoA-predicted paternity distribution in every mating season and relate deviation to (i) the number of coresident males, (ii) the number of females that conceive in a given mating season, and (iii) the interaction of (i) and (ii). We do not consider the costs of mate guarding, female choice, and male coalitions leveling the mating skew as factors related to deviation from PoA in this study because (i) our research shows that extensive male mate guarding is performed almost exclusively by the highest-ranking males, follows a PoA-predicted pattern instead of deviating from it (Ostner et al. 2011), and does not seem to carry energetic costs for guarding males (Schülke et al. 2014); (ii) because of low sampling density, data indicative of female choice were too scarce to allow sensible analyses of its differential effect on deviation from PoA-predicted paternity success; and (iii) because coalitions did not serve to level the mating skew in our study group but have other functions (Young et al. 2014).
Methods
Study site and study group
The study area Huai Mai Sot Yai is situated in Phu Khieo Wildlife Sanctuary (PKWS, 16° 05′–35′ N and 101° 20′–55′ E) that is part of the 6,500-km2 Western Isaan Forest Complex in Thailand and comprises well-protected natural hill and dry evergreen forest with patches of bamboo forest (Borries et al. 2002). The mammal fauna is rich with large herbivores and carnivores, the vegetation is dense, and the terrain is hilly (Kumsuk et al. 1999; Borries et al. 2002). Assamese macaques are strictly seasonal breeders, they mate from October to January/February and give birth between April and June/July (Fürtbauer et al. 2010). The study spans the mating seasons 2005/2006–2010/2011 in study group AS that ranged in size from 47 to 56 individuals with 9–15 males and 12–15 females. We collected fecal samples of all individuals in the study group for DNA analyses and only missed 3 out of 46 offspring born in 6 years. For analyses, the mating season 2008/2009 was divided into two subperiods (2008/2009 1: 1 October to 2 December 2008; 2008/2009 2: 3 December 2008 to the end of the mating season) because of drastic changes in male group composition and dominance ranks.
Behavioral data collection
Behavioral data were collected from October 2006 to February 2011 using 20- or 30-min focal animal sampling of males (Altmann 1974). Observations were approximately evenly distributed across 2-h blocks from 6 a.m. to 6 p.m. and across focal animals and supplemented with ad libitum sampling (Martin and Bateson 1993). During observations, all social and sexual interactions were recorded including submissive behaviors such as give ground, make room, flee, crouch and silent bared-teeth display, and aggressive behaviors including bite, head bob, chase, lunge, open-mouth display, ground slap, push-pull, point, slap, and stare (Ostner et al. 2008b).
The ordinal dominance hierarchy was constructed based on the exchange of clear submissive signals (make room and bared teeth) in decided dyadic agonistic interactions (i.e., when one individual showed only submission while the other did or did not show aggression but no submission) that were used to calculate the corrected normalized David’s score (nDS, de Vries et al. 2006), a linear measure of dominance. The hierarchy was based on 290.4 ± 86.3 interactions per season; was significantly linear in all cases, left 19.3 ± 15.4 % of relationships unknown; and had a directional consistency index of 91.9 ± 3.9. Only 13 % of dyads on average had two-way dominance relations. Inspection of the two-way relationships did not suggest definite rank changes during a mating season except in mating season 2008/2009 that was consequently split into two and was treated as two separate season in all analyses. More details on male dominance relations and how they are related to mating success, mate-guarding activity, as well as androgen and glucocorticoid excretions can be found in Ostner et al. (2008b; 2011). No dominance data were available for mating season 2005/2006.
Predicting male paternity success from the PoA model
According to the PoA model, male dominance hierarchies function as queues in which males wait for mating opportunities (Alberts et al. 2003). The logic is that a lower-ranked male will be able to mate guard a female only when higher-ranking males are occupied guarding other mates. Other forms of direct competition should be affected by the number of synchronously available females as well. Female Assamese macaques that will conceive in a given mating season do not show behavioral estrus, and their mating frequency is consistently low and not related to their hormonal status (Fürtbauer et al. 2011). Males seem to neither distinguish between peri-ovulatory periods and nonfertile days nor between conceptive and nonconceptive cycles (Fürtbauer et al. 2011). The original PoA model used the average proportion of time a female was in estrus to predict the overlap of female receptive phases for a given number of females per group in rhesus macaques (Altmann 1962). Because receptivity is even more extended and ovulation concealed in female Assamese macaques, all females that conceived in a given mating season (always a subset of resident females) were considered receptive and attractive throughout the entire mating season when calculating the predictions of the PoA model. We have previously shown that the probability to conceive depends on the timing of the last birth (Fürtbauer et al. 2010), food availability, and female physical condition prior to the onset of the mating season (Heesen et al. 2013). At the onset of the mating season, it is decided who will “cycle” and conceive in a given mating season and who will not, and the increase in female androgen levels prior to the onset of the mating season determines the timing of conception within the mating season (Fürtbauer et al. 2013). Together, this creates considerable overlap of receptive periods which (according to PoA) provides many males with mating opportunities even if they strictly queue by rank. The expected proportion of offspring sired by a male on a given rank position is one/number of conceiving females in that season for alpha males down to the rank that equaled the number of conceiving females in that season. Thus, the expected paternity success is equal for all males that are expected to get any share and varies between seasons as a strict function of the number of conceiving females.
Paternity assignment
All fecal samples were collected immediately after defecation and stored using the two-step ethanol-silica storage method (Nsubuga et al. 2004). Genomic DNA was extracted from 0.1 g of fecal samples by using the QIAamp stool kit (QIAGEN) with slight modifications (Nsubuga et al. 2004), and blanks were included during extraction to monitor for contamination.
PCR reactions for microsatellite analysis for all individuals were performed with a two-step multiplex polymerase chain reaction protocol (Arandjelovic et al. 2009). We used 15 microsatellite markers with 5′ fluorescently labeled forward primers. Primer names and 2-step PCR conditions have been reported elsewhere (Schülke et al. 2010). Multiple negative controls were added in each reaction for monitoring the contamination. The concentration of PCR product was estimated by gel electrophoresis, and all successful PCR reactions were subjected to fragment analysis.
The genotyping was done on two systems because lab work was first conducted in Leipzig, Germany, and later in Bangkok, Thailand. Genotyping of samples from mating seasons 2005/2006–2008/2009 was performed on an ABI system (ABI 3100 genetic analyzer; Applied Biosystems), and GeneMapper (version 3.7) software was employed to assess fragment size compared with a ROX HD400 internal size standard (Schülke et al. 2010). Samples of offspring conceived in mating seasons 2009/2010–2010/2011 were genotyped with a CEQ system (CEQ™ 8000 Genetic Analysis; Beckman Coulter, Inc.), and CEQ™ 8000 Genetic Analysis System (Beckman Coulter, Inc.) was used for fragment size assessment compared to a GenomeLab DNA Size Standard Kit—400 internal standard sizes. In order to avoid genotyping errors due to the change of systems, nine samples that had been genotyped with the ABI system were also analyzed with the CEQ system which allowed comparing and standardizing the allele size scores. A locus was accepted as heterozygous when it was amplified at least two times in independent PCR reactions. For a locus to be scored as homozygous, it needed to show one allele only in at least four reactions (Morin et al. 2001; Schülke et al. 2010). An allele which was observed only once in six replicate reactions was rejected as a false allele. A maximum of nine replicates were run. Individual identity was confirmed by comparing the genotypes of known mother-offspring pairs, i.e., genotyping two or more independently extracted DNA from the same identified individual in order to confirm the identity of that individual (average of 2.23 samples/individual). Expected heterozygosity, the exclusionary power of the locus, and the deviation from Hardy-Weinberg equilibrium (HWE) for each locus were calculated by ARLEQUIN v.3.1. F is was calculated by FSTAT 2.9.3.2 (Goudet 1995).
We identified the paternity of 43 offspring born between 2006 and 2011 combining exclusion and likelihood approaches. All 18 adult and large subadult males that resided in the group over that period were included in the paternity analysis until their death was verified (three males). Since subadult males have been shown to sire offspring sometimes (de Ruiter et al. 1992), we also included all four small subadult males. Eighteen females were included in our analyses as mothers. In addition, five juveniles were included in the analysis of allele frequencies (88 individuals in total). A male was excluded from paternity if he showed one allelic mismatch with the specific mother-offspring dyad. The remaining male that was not excluded after comparing all loci was considered the most likely father of the respective offspring. Subsequently, we used CERVUS 3.0.3 (Field Genetic Ltd., Marshall et al. 1998) to confirm paternity in two sets of likelihood analyses with (1) mothers as known from behavioral observations and (2) with the most likely mother among all adult females given hers and the offspring’s genotype. Likelihood analyses with 95 % confidence levels confirmed all our maternity assignments from behavioral observations. Thus, the paternity analysis (1) with one parent known was supported. In order to confirm assigned paternities derived via exclusion, all paternities were additionally supported at the 95 % confidence level in favor of the male with the highest LOD score calculated by CERVUS 3.0.3 (genotyping error rate set to 0.041 as calculated by CERVUS from mother-offspring mismatches, minimum number of loci typed 9, 22 candidate fathers).
The mean number of alleles per locus was 6.93. The range of observed heterozygosity (H O) was 0.47–0.89 and 0.72 on average. The mean expected heterozygosity (H E) was 0.715 and the range 0.46–0.90 (Supplementary Material Table 1). The mean polymorphic information content (P IC) over 15 microsatellite markers was 0.66. The mean ± SD nonexclusion probability of a randomly chosen male as the second parent was 0.000038 ± 0.000076 across offspring (range 0.000000013–0.00040), the individual probability of identity (P ID) was 5.38 × 10−15, and the probability of sib identity (P IDsib) was 2.53 × 10−6. Thus, it is extremely unlikely that another male than the assigned father has sired the offspring, i.e., paternity probability is very high.
Data analysis
To calculate whether the observed skew was different from a random distribution, we chose Nonacs’ binomial skew index or B index (Nonacs 2000, 2003) which assesses male skew independently of male dominance hierarchy. The B index was calculated using the program Skew Calculator 2003 (https://www.eeb.ucla.edu/Faculty/Nonacs/PI.html) and tested whether the observed B index occurred by chance. Large values of the B index above one indicate high reproductive skew. We calculated the B index for the observed number of offspring sired by each male across 2006–2011.
To estimate the deviation from the PoA model, we built linear mixed models (LMMs) with Gaussian error structure and log link function in R 2.15.0 using the function lmer of the package lme4 (Bates et al. 2012; R Development Core Team 2012). We used the proportion of offspring sired on every rank position in a single period as the response (i.e., one data point per rank position per season) and the proportion of offspring expected to be sired from PoA predictions on that rank position that year, the number of coresident males, the number of conceiving females, and the interaction between the number of males and number of conceiving females as fixed effects and male ID and season as random effects. The response variable was first log transformed to better fit assumptions about normality. All variables were then z transformed. To control for possible within-subject or within-season differences, we included random slopes as well as random intercepts in our models (Schielzeth and Forstmeier 2009). We used likelihood ratio tests to examine the significance of the full model versus the null model (including only random factors) using the R function ANOVA. We also compared the full model with a model excluding random slopes using the function ANOVA and found no difference between the models indicating that there were no significant within-subject differences in the data. Assumptions about normality and homogeneous residuals were respected for the analyses, and colinearity was not a problem in our analyses as indicated by variation inflation factors generated from general regression models all being below 3 (Bowerman and O’Connell 1990; Mayers 1990), calculated using the function Vif from the R package car (Fox and Weisberg 2010). To increase the reliability of our P values, we fitted the model with a maximum likelihood rather than a restricted likelihood (Bolker et al. 2009). Markov chain Monte Carlo sampling was used to generate P values for the individual effects (Baayen 2008) derived from the function pvals.fnc from the R package languageR (Baayen 2010).
For comparison with published studies, we also ran a simple regression of observed overexpected paternity success across the entire study period in Statistica 10 (StatSoft, Tulsa, OK, USA). Inspection of residuals (see above) did not indicate problems with violation of regression assumptions. It may be argued that the model fit is increased artificially if too many low-ranking males are included in the analysis that have both zero observed and zero expected paternity success (see Bercovitch 1986; McMillan 1989 for similar arguments). Exclusion of males ranked 11 and lower did not change the significance of predictors in our models. Explained variance changed only slightly to 80 % when we excluded ranks 11 and up. Alpha was set to 0.05 and all tests were two tailed.
Results
Description of paternity success across males and ranks
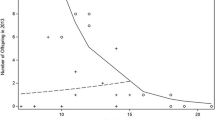
A majority of 61 % (11/18) of males sired at least one offspring. The average number of infants sired per male across the study period was 2.4 ± 2.6 (mean ± SE) and ranged from zero to nine. All offspring were fathered by resident males, and we detected no evidence of extra-group paternity. The alpha males sired 29 % (11/38) of all infants born into the study group, and paternity success across the study period was highly correlated with male dominance rank (average number of offspring sired across years correlated to dominance rank position with alpha-assigned rank 1: Spearman’s rho = −0.90, P < 0.001). This correlation was maintained if only the top half of the hierarchy was considered (ranks 1–7: rho = 0.90; P < 0.01). The observed paternity distribution across ranks for 2006/2007–2010/2011 was shallow but significantly skewed (B = 0.087, P < 0.0001). Sires ranked between the alpha and as low as the 12th position in the male hierarchy, but males that ranked 8th and 11th did not father an offspring (Fig. 1).

PoA-predicted and PoA-observed reproductive skews (paternity success) by dominance rank from 2006/2007 to 2010/2011; upper pane regression of observed over expected percent offspring sired by males on specific dominance rank positions (line linear regression), lower pane observed percent offspring sired over rank position of the male and expected percent of offspring sired over rank
Paternity success and male mate guarding
Male mate guarding increased paternity success. In mating seasons 2007/2008 and 2008/2009, 11 out of 14 conceiving females were mate guarded throughout or for parts of the fertile phase of their conception cycle by one or several males in succession. In 64 % of the cases, the only guarding male or the one with the highest mate-guarding frequency sired the offspring which is a much larger proportion than the 29 % of all paternities that the most successful male can secure in the long run. For this analysis, we added information on paternity and on short consortships (<1-week duration) to the published data set on timing of ovulation and male mate-guarding activity in Fürtbauer et al. (2011).
Observed paternity success and predictions of the PoA model
For comparison with other studies, we ran a simple regression of observed overexpected paternity success and found that paternity success on a given rank position across the study period was predicted by the logic of the PoA model (adj. R 2 = 0.62, F 1,13 = 23.7, P < 0.001, Fig. 1). The largest deviations were found at ranks 1, 4, and 7.
In order to describe variation over time in deviation from PoA predictions (short-term variation), we split the data into mating seasons. In some mating seasons, the fit between observed paternity distributions and each PoA expected distribution was less pronounced than that across the entire study period (Fig. 2). The alpha males sired at least one offspring every year, never sired less than the predicted proportion of offspring, and had the highest paternity success in half of the cases. In only three cases, from three different mating seasons, a single offspring was sired by a low-ranking male (ranks 9, 10, and 12, respectively) that was not predicted to sire offspring in that season. In the remaining 33 cases, the right tails of the expected and the observed distributions matched at zero paternity success. In accordance with the logic of the PoA model, the number of different males that sired offspring was positively correlated to the number of conceiving females (Spearman’s rho = 0.72, P < 0.05). We investigated potential predictors of deviation from PoA in LMMs of the observed proportion of offspring sired on every rank position in every period (Table 1). Both the models with and the one without the interaction between the number of males and number of conceiving females as a predictor were significant (full model vs null model including random effects Chi2 = 9.6, P < 0.05; reduced model without interaction term vs null model including random effects Chi2 = 9.5, P < 0.05). Expected paternity success was the only significant predictor in all models (Table 1). Residual variation after accounting for the effect of expected paternity success can be construed as the deviation from PoA. The interaction between the number of males and the number of conceiving females did not explain this deviation. In a reduced model without the interaction, neither the number of conceiving females nor the number or coresident males explained the significant parts of the residual variation.

a–e “Short-term” deviation from PoA-predicted reproductive skew across six mating seasons. Expected and observed percent offspring sired plotted over male rank; with each plot, the ID of the alpha male and the number of offspring genotyped are given
Discussion
In accordance with general patterns across mammals living in multimale groups (Say et al. 2001; Engh et al. 2002; Alberts et al. 2006; Rodriguez-Llanes et al. 2009; Wroblewski et al. 2009), paternity in wild Assamese macaques was significantly skewed toward higher-ranking males, paternity was predicted by dominance rank, and across the entire study period, alpha males sired the largest proportion of offspring. The observed reproductive skew was quite low, however, with B = 0.87, 29 % alpha male share of paternity and only a third of males not siring any offspring. The three males that held the alpha position all had similar paternity success of 28, 25, and 33 %, respectively. Both the distribution across ranks and the shallow skew across the study period were not unexpected given the logic of the PoA model that rank-related reproductive skew is negatively affected by the degree of female reproductive synchrony. Thus, within its limits, our 6-year study suggests that constraints on male monopolization may affect male reproductive skew in predictable ways even when female reproductive synchrony is very high and the species reproduces seasonally.
Observed paternity distribution also deviated to some extent from PoA predictions. Expected rank-related reproductive skew (16 % alpha male paternity) was even shallower than observed patterns, observed paternity success did not decrease strictly with rank, and “short-term” deviation of observed paternity from expected success was pronounced in some mating seasons and could not be explained by variation in the number of males or conceiving females or their interaction. In the following, we first go through caveats of our study and then discuss the state of the art concerning factors predicting deviation from PoA-predicted paternity skew.
Our analyses were set up as test of the logic of the PoA model. We investigated the combined effects of male monopolization and other mechanisms of direct precopulatory competition like the dominants’ ability to aggressively interfere in others’ mating and subordinate avoidance of such aggression. We have shown here that monopolization in the form of mate guarding during the peri-ovulatory phase (possessive strategy sensu Wroblewski et al. 2009) is imperfect, because the guarding male often did not sire the offspring. Nevertheless, monopolization seems to increase the siring probability with individual guarded females (64 %) above the level that can be achieved across females by the most successful males (28 %). Strong rank effects on male mate-guarding activity (Ostner et al. 2011) suggest that this form of access is contested among males. It remains unclear, however, whether the assumption of the PoA model that male mating success predicts male reproductive success is met in Assamese macaques. This test would require dense observational data on mating during many peri-ovulatory phases which are difficult to obtain if females do not show morphological or behavioral signs of ovulation (Fürtbauer et al. 2010; 2011).
The observed paternity distribution could also result from other mechanisms of sexual selection than direct male competition via monopolization, interference in others’ mating, and avoidance of aggression (Engelhardt et al. 2006). First, frequent mating with individual females (sensu Møller and Birkhead 1991) could be used as an alternative strategy if mate guarding is ineffective and sperm competition ensues. Sperm competition does occur, because female Assamese macaques mate with all resident males over the course of a mating season and usually with more than one male during the peri-ovulatory phase of their conceptive cycles (Fürtbauer et al. 2011). For the reasons laid out above, good estimates of relative male mating success during these phases are lacking though. Thus, it has to remain unclear whether frequent mating (or cryptic female choice sensu Eberhardt 1996) affected male reproductive skew. Males may also succeed in sperm competition, because their ejaculates have more, faster, or otherwise higher-quality sperm (Stockley 2004; Tourmente et al. 2006). If this mechanism was the main driver of paternity distribution in our study, success in sperm competition would have to be related to dominance rank. However, we have shown previously that male androgen levels which play a pivotal role in sperm production and quality (Sharpe 1994; McLachlan et al. 2002) are not related to male dominance rank in our study group (Ostner et al. 2011). Therefore, it seems unlikely that postcopulatory male competition was the main factor driving paternity distribution in this study.
As we have laid out earlier (Fürtbauer et al. 2011; Ostner et al. 2011), a high degree of female reproductive synchrony as found in Assamese macaques facilitates precopulatory female choice, because monopolization of access to all receptive females by a single or few males is hampered. Compared to situations with reproductive asynchrony, more females, i.e., those females that are not being monopolized by the highest-ranking males, may more easily realize their mate preferences (Fürtbauer et al. 2011; Ostner et al. 2011). Female behavior may also affect which female is guarded by one of the few top dominants that use the possessive strategy, because receptive females are always in oversupply and males could move on to another female if their initial partner was uncooperative. Thus, female choice may influence both whether a male can guard a female for extended periods in the possessive form and whether a male and a female simply associate spatially more often and longer (consortship strategy sensu Wroblewski et al. 2009). Male-female affiliation or association has recently been shown to affect male mating and paternity success in rhesus macaques, chimpanzees, Assamese macaques, and in all species with low expected and observed reproductive skews (Kulik et al. 2011; Massen et al. 2012; Langergraber et al. 2013; Ostner et al. 2013). With the exception of the Massen et al. (2012) study in which both males and females invested heavily in the affiliative relationship, male-female sociality was measured on the dyadic level with a socio-spatial index or a composite sociality index. Therefore, it remains to be established whether high rates of male-female affiliation and association result from female mate choice. In all chimpanzee communities studied to date as well as in Assamese macaques, paternity success is positively correlated to male dominance rank though (Boesch et al. 2006; Wroblewski et al. 2009; Newton-Fisher et al. 2010) a pattern seems unlikely to result from female choice alone.
All these other mechanisms are set aside, parts of the deviation of observed from expected paternity success in our study may result from two methodological issues. First, expected paternity distribution was calculated based on the number of females that conceived during a given mating season irrespective of whether the offspring was included in the paternity analyses or not. In three cases (one in 2008, two in 2009–2), offspring could not be genotyped, because they died before a sample was taken. Thus, the expected values mirror the competitive situation but could have never been matched perfectly by observed values. Second, information about the degree to which males can predict the probability of female ovulation is crucial for calculating PoA-predicted paternity distributions. The positive deviation of alpha male paternity from the expected value may have resulted from an overly conservative interpretation of male behavior in relation to female fertility in Assamese macaques (Fürtbauer et al. 2011), indicating that males consider all conceiving females as equally attractive from the onset to the end of the mating season. This problem may partly be responsible for the variation in deviation from PoA across different studies.
It is currently difficult to assess the generality of the PoA model as a mechanism generating reproductive skew across mammals living in multimale groups. Few studies provide the necessary data on paternity, dominance rank, and the overlap of female reproductive phases. In mandrills (Mandrillus sphinx; Setchell et al. 2005) and chimpanzees at Taï and Budongo (Boesch et al. 2006; Newton-Fisher et al. 2010), alpha males sired 5–30 % more offspring than expected, while in rhesus macaques (Dubuc et al. 2011), chimpanzees at Gombe (Wroblewski et al. 2009), and yellow baboons (Papio cynocephalus; Alberts et al. 2006), the alpha male share was lower than expected from the PoA model. The variation in deviation from PoA model predictions both between species/populations and within groups over time may stem from sampling issues, methodological differences, theoretical issues, or actual differences in male competitive regimens.
Sampling issues may arise when tests of the model’s fit are run on small numbers of genotyped offspring. The PoA model predicts the proportion of offspring sired on a continuous scale while the observed values are generated from count data that produce step functions with large increments when the number of observations is small. The residuals become smaller as the number of females (and offspring) increases which makes it difficult to test the prediction that deviation from PoA increases with female group size. However, the study with the largest sample size (208 offspring) also produced the largest deviation from PoA-predicted patterns (Alberts et al. 2006).
A methodological issue concerns the accuracy of male dominance rank assessment. Discrepancies between expected paternity success on a given rank position and observed success on that position may simply result from rank assessments being inadequate or slow at picking up changes in the hierarchy. Some studies assume rank to be relatively stable and include years for which no observational data on male agonistic relationships are available. Other studies do not report how many observations their dominance assessments are based upon or which method was used to order males into a hierarchy so that the quality of the rank assessment remains unclear. Despite the efforts we made to avoid this problem (see “Methods”), it still seems possible that males and females sense upcoming rank changes before they are reflected in decided dyadic conflicts.
A theoretical explanation for the observed variation in deviation from the PoA model predictions is that the alpha male’s goal may not always be to monopolize all reproduction but that alpha males cede reproduction to lower-ranking males in certain situations. These concessions may be adaptive (i) if subordinates support the alpha in conflicts with competitors from outside the group as in geladas (Theropithecus gelada, Snyder-Mackler et al. 2012) or from within the group as in chimpanzees (Duffy et al. 2007) or (ii) if subordinates aid in the protection of offspring (likely sired by the alpha male) for which subordinates have a nonzero chance of paternity (Boyko and Marshall 2009; chacma baboons, Papio ursinus, Henzi et al. 2010) or (iii) if the subordinate is very closely related to the alpha male (assessed by Widdig et al. 2004; Snyder-Mackler et al. 2012). Theoretical reproductive skew models are not readily applicable here, because they make explicit assumptions that are usually not fulfilled in nonhuman primates (Port and Kappeler 2010). Generalizations about male relatedness, e.g., based on male dispersal regime, do not easily explain variation in the PoA model fit across species because positive and negative deviations are observed in both male philopatric (Boesch et al. 2006 vs. Wroblewski et al. 2009) and male dispersal species (Setchell et al. 2005 vs. Dubuc et al. 2011). We refrained from analyzing relatedness effects on reproductive skew, because dyadic relatedness estimates based on relatively small numbers of autosomal microsatellite markers are associated with large errors (Csilléry et al. 2006; Van Horn et al. 2008).
Although not significant in our study, variation in the number of competitors has received most support as a driver of alpha male paternity (de Ruiter et al. 1992; Muniz et al. 2010; Gogarten and Koenig 2012) or mating success (Cowlishaw and Dunbar 1991; Kutsukake and Nunn 2006) and of deviation from PoA-predicted paternity distribution (Alberts et al. 2006; Boesch et al. 2006). The more males coreside in a group, the lower the alpha share and the more pronounced the deviation from PoA (but see Charpentier et al. 2005; Henzi et al. 2010) are, which has been explained by at least three mechanisms. First, increasing numbers of competitors may require high-ranking males to increase investment into fending-off rivals from proximity to a guarded female. Where mate guarding carries time or energy constraints (Alberts et al. 1996; Higham et al. 2011), additional chases and fights may render mate guarding economically inefficient and consequently may cause negative deviation from PoA-predicted skew. Male Assamese macaques do not suffer these mate-guarding costs though (Schülke et al. 2014). Second, more males may decrease high-ranking males’ efficiency at prohibiting sneak copulations (Soltis et al. 2001; Overduin-De Vries et al. 2012). It is currently not possible to assess the generality of this argument for deviation from PoA across primates, but densely structured habitats and higher female cycle synchrony should both promote this alternative mating strategy. In our study group, not all females were always mate guarded, and females managed to mate with virtually all males in the group over the course of the mating season (Fürtbauer et al. 2011; Ostner et al. 2011). It may be argued that all males should be equally efficient in sneaking copulations, and paternity should therefore be reallocated from high-ranking males to any resident. The largest positive deviations from PoA expectations, however, were observed in the top third of the dominance hierarchy, and only 7 % of offspring were sired by low-ranking males from the bottom third of the hierarchy. Third, an increase in the number of males may promote the formation of coalitions that level the mating skew. The number of coresident males was a strong predictor of the frequency of male coalition formation across primates in a comparative analysis (Bissonnette et al. 2014). Leveling coalitions may peripheralize males to restrict their access to fertile females (Bissonnette et al. 2011) but are used more commonly to break up a consortship/mate-guarding episode between a male (usually high ranking) and a female (Noë and Sluijter 1990; Kuester and Paul 1992; Bissonnette et al. 2011). Leveling coalitions may be a major driver of deviation from PoA-predicted paternity distribution in yellow baboons (Alberts et al. 2006). Although, mathematical models suggest that leveling coalitions evolve when male contest competition potential is low (Pandit and van Schaik 2003); male coalitions do not affect mating skew in rhesus or Assamese macaques where both expected and observed paternity skews are low (Higham and Maestripieri 2010; Dubuc et al. 2011; Ostner and Schülke 2014; Young et al. 2014).
Although the role of female cooperation in both the possessive and the consortship strategies remains to be assessed in more detail, we conclude that the rather close match of expected and observed paternity distributions in relation to dominance rank suggests that even when (i) males cannot discern the female fertile phase (Engelhardt et al. 2006), (ii) reproduction is seasonal (Dubuc et al. 2011), and (iii) female reproductive synchrony is high, males in multimale groups may still compete also directly over access to receptive females and therefore invest in attaining and maintaining high social status.
References
Alberts SC (2012) Magnitude and sources of variation in male reproductive performance. In: Mitani JC, Call J, Kappeler PM, Palombit RA, Silk J (eds) Evolution of primate societies. University of Chicago Press, Chicago, pp 412–431
Alberts SC, Altmann J, Wilson ML (1996) Mate guarding constrains foraging activity of male baboons. Anim Behav 51:1269–1277
Alberts SC, Heather EW, Altmann J (2003) Queuing and queue-jumping: long-term patterns of reproductive skew among male savannah baboons. Anim Behav 65:821–840
Alberts SC, Buchan JC, Altmann J (2006) Sexual selection in wild baboons: from mating opportunities to paternity success. Anim Behav 72:1177–1196
Altmann SA (1962) A field study of the sociobiology of rhesus monkeys, Macaca mulatta. Ann N Y Acad Sci 102:338–435
Altmann J (1974) Observational study of behavior: sampling methods. Behaviour 49:227–267
Arandjelovic M, Guschanski K, Schubert G, Harris TR, Thalmann O, Siedel H, Vigilant L (2009) Two-step multiplex PCR improves the speed and accuracy of genotyping using DNA from noninvasive and museum samples. Mol Ecol Resour 9:28–36
Baayen HR (2008) Analyzing linguistic data: a practical introduction to statistics using R. Cambridge University Press, Cambridge
Baayen RH (2010) language4: Data sets and functions with “Analyzing Linguistic Data: a practical introduction to statistics” http://CRAN.R-project.org/package=language4
Bates D, Maechler M, Bolker B (2012) lme4: linear mixed effects models using S4 classes. In: R package version 0.999999-0. http://CRAN.R-project.org/package=lme4
Bercovitch FB (1986) Male rank and reproductive activity in savannah baboons. Int J Primatol 7:533–550
Bissonnette A, Bischofberger N, van Schaik C (2011) Mating skew in Barbary macaque males: the role of female mating synchrony, female behavior, and male–male coalitions. Behav Ecol Sociobiol 65:167–182
Bissonnette A, Franz M, Schülke O, Ostner J (2014) Socioecology, but nor cognition, predicts male coalitions across primates. Behav Ecol (in press)
Boesch C, Kohou G, Nene H, Vigilant L (2006) Male competition and paternity in wild chimpanzees of the Taï forest. Am J Phys Anthropol 130:103–115
Bolker BM, Brooks ME, Clark CJ, Geange SW, Poulsen JR, Stevens MHH, White J-SS (2009) Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol Evol 24:127–135
Borries C, Larney E, Kreetiyutanont K, Koenig A (2002) The diurnal primate community in a dry evergreen forest in Phu Khieo Wildlife Sanctuary, Northeast Thailand. Nat Hist Bull Siam Soc 50:75–88
Bowerman B, O’Connell R (1990) Linear statistical models: an applied approach, 2nd edn. Duxbury, Belmont
Boyko RH, Marshall AJ (2009) The willing cuckold: optimal paternity allocation, infanticide and male reproductive strategies in mammals. Anim Behav 77:1397–1407
Charpentier M, Peignot P, Hossaert-McKey M, Gimenez O, Setchell JM, Wickings EJ (2005) Constraints on control: factors influencing reproductive success in male mandrills (Mandrillus sphinx). Behav Ecol 16:614–623
Clutton-Brock TH (1998) Reproductive skew, concessions and limited control. Trends Ecol Evol 13:288–292
Clutton-Brock TH, Parker GA (1992) Potential reproductive rates and the operation of sexual selection. Q Rev Biol 67:437–456
Clutton-Brock TH, Vincent ACJ (1991) Sexual selection and the potential reproductive rates of males and females. Nature 351:58–60
Cowlishaw G, Dunbar RIM (1991) Dominance rank and mating success in male primates. Anim Behav 41:1045–1056
Csilléry K, Johnson T, Beraldi D, Clutton-Brock T, Coltman D, Hansson B, Spong G, Pemberton JM (2006) Performance of marker-based relatedness estimators in natural populations of outbred vertebrates. Genetics 173:2091–2101
de Ruiter JR, Scheffrahn W, Trommelen GJJM, Uitterlinden AG, Martin RD, van Hoof JARAM (1992) Male social rank and reproductive success in wild long-tailed macaques. In: Martin RD, Dixson AF, Wickings EJ (eds) Paternity in primates: genetic tests and theories. Karger, Basel, pp 175–191
de Vries H, Stevens J, Vervaecke H (2006) Measuring and testing the steepness of dominance hierarchies. Anim Behav 71:585–592
Dubuc C, Muniz L, Heistermann M, Engelhardt A, Widdig A (2011) Testing the priority-of-access model in a seasonally breeding primate species. Behav Ecol Sociobiol 65:1615–1627
Duffy KG, Wrangham RW, Silk JB (2007) Male chimpanzees exchange political support for mating opportunities. Curr Biol 17:R586–R587
Eberhardt WG (1996) Female control: sexual selection by cryptic female choice. Princeton University Press, Princeton
Ellis L (1995) Dominance and reproductive success among non-human animals: a cross-species comparison. Ethol Sociobiol 16:257–333
Emlen ST, Oring LW (1977) Ecology, sexual selection, and the evolution of mating systems. Science 197:215–223
Engelhardt A, Heistermann M, Hodges JK, Nurnberg P, Niemitz C (2006) Determinants of male reproductive success in wild long-tailed macaques (Macaca fascicularis)—male monopolisation, male mate choice or post-copulatory mechanisms? Behav Ecol Sociobiol 59:740–752
Engh AL, Funk SM, van Horn RC, Scribner KT, Bruford MW, Libants S, Szykman M, Smale L, Holekamp KE (2002) Reproductive skew among males in a female-dominated mammalian society. Behav Ecol 13:193–200
Fox J, Weisberg HS (2010) An R companion to applied regression. Sage, Thousand Oaks
Franz M, van der Post D, Schülke O, Ostner J (2011) The evolution of cooperative turn-taking in animal conflict. BMC Evol Biol 11:323
Fürtbauer I, Schülke O, Heistermann M, Ostner J (2010) Reproductive and life history parameters of wild female Macaca assamensis. Int J Primatol 31:501–517
Fürtbauer I, Heistermann M, Schülke O, Ostner J (2011) Concealed fertility and extended female sexuality in a non-human primate (Macaca assamensis). PLoS ONE 6:e23105
Fürtbauer I, Heistermann M, Schülke O, Ostner J (2013) Female fecal androgens prior to the mating season reflect readiness to conceive in reproductively quiescent wild macaques. Am J Phys Anthropol 151:311–315
Gogarten JF, Koenig A (2012) Reproductive seasonality is a poor predictor of receptive synchrony and male reproductive skew among nonhuman primates. Behav Ecol Sociobiol 67:123–134
Goudet J (1995) FSTAT (Version 1.2): a computer program to calculate F-statistics. J Hered 86:485–486
Henzi SP, Clarke PMR, van Schaik CP, Pradhan GR, Barrett L (2010) Infanticide and reproductive restraint in a polygynous social mammal. Proc Natl Acad Sci U S A 107:2130–2135
Heesen M, Rogahn S, Ostner J, Schülke O (2013) Food abundance affects energy intake and reproduction in frugivorous female Assamese macaques. Behav Ecol Sociobiol 67:1053–1066
Higham JP, Maestripieri D (2010) Revolutionary coalitions in male rhesus macaques. Behaviour 147:1889–1908
Higham JP, Heistermann M, Maestripieri D (2011) The energetics of male–male endurance rivalry in free-ranging rhesus macaques, Macaca mulatta. Anim Behav 81:1001–1007
Johnstone RA (2000) Models of reproductive skew: a review and synthesis. Ethology 106:5–26
Kuester J, Paul A (1992) Influence of male competition and female mate choice on male mating success in Barbary macaques (Macaca sylvanus). Behaviour 120:192–217
Kulik L, Muniz L, Mundry R, Widdig A (2011) Patterns of intervention and the effect of coalitions and sociality on male fitness. Mol Ecol 21:699–714
Kumsuk M, Kreetiyutanont K, Suvannakorn V, Sanguanyat N (1999) Diversity of wildlife vertebrates in Phu Khieo Wildlife Sanctuary, Chaiyaphum Province. Phu Khieo Wildlife Sanctuary, Wildlife Conservation Division, Natural Resources Conservation Office, Royal Forest Department, Bangkok, p 124
Kutsukake N, Nunn CL (2006) Comparative tests of reproductive skew in male primates: the roles of demographic factors and incomplete control. Behav Ecol Sociobiol 60:695–706
Langergraber KE, Mitani JC, Watts DP, Vigilant L (2013) Male–female socio-spatial relationships and reproduction in wild chimpanzees. Behav Ecol Sociobiol 67:861–873
Majolo B, Lehmann J, de Bortoli VA, Schino G (2012) Fitness-related benefits of dominance in primates. Am J Phys Anthropol 147:652–660
Marshall TC, Slate J, Kruuk LEB, Pemberton JM (1998) Statistical confidence for likelihood-based paternity inference in natural populations. Mol Ecol 7:639–655
Martin P, Bateson P (1993) Measuring Behaviour. Cambridge University Press, Cambridge
Massen JJM, Overduin-de Vries AM, de Vos-Rouweler AJM, Sptuijt BM, Doxiadis GGM, Sterck EHM (2012) Male mating tactics in captive rhesus macaques (Macaca mulatta): the influence of dominance, markets and relationship quality. Int J Primatol 33:73–92
Mayers R (1990) Classical and modern regression with applications, 2nd edn. Duxbury, Boston
McLachlan RI, O’Donnell L, Meachem SJ, Stanton PG, de Kretser DM, Pratis K, Robertson DM (2002) Identification of specific sites of hormonal regulation of spermatogenesis in rats, mice, and man. Recent Prog Horm Res 57:149–179
McMillan CA (1989) Male age, dominance, and mating success among rhesus macaques. Am J Phys Anthropol 80:83–89
Møller AP, Birkhead TR (1991) Frequent copulations and mate guarding as alternative paternity guards. Behaviour 118:170–186
Morin PA, Chambers KE, Boesch C, Vigilant L (2001) Quantitative polymerase chain reaction analysis of DNA from noninvasive samples for accurate microsatellite genotyping of wild chimpanzees (Pan troglodytes verus). Mol Ecol 10:1835–1844
Muniz L, Perry S, Manson JH, Gilkenson H, Gros-Louis J, Vigilant L (2010) Male dominance and reproductive success in wild white-faced capuchins (Cebus capucinus) at Lomas Barbudal, Costa Rica. Am J Primatol 72:1118–1130
Nadler R (1994) Walter Heape and the issue of estrus in primates. Am J Primatol 33:83–87
Newton-Fisher NE, Thompson ME, Reynolds V, Boesch C, Vigilant L (2010) Paternity and social rank in wild chimpanzees (Pan troglodytes) from the Budongo Forest, Uganda. Am J Phys Anthropol 142:417–428
Noë R, Sluijter AA (1990) Reproductive tactics of male savanna baboons. Behaviour 113:117–170
Nonacs P (2000) Measuring and using skew in the study of social behavior and evolution. Am Nat 156:577–589
Nonacs P (2003) Measuring the reliability of skew indices: is there one best index? Anim Behav 65:615–627
Nsubuga AM, Robbins MM, Roeder A, Morin PA, Boesch C, Vigilant L (2004) Factors affecting the amount of genomic DNA extracted from ape feces and the identification of an improved sample storage method. Mol Ecol 13:2089–2094
Ostner J, Schülke O (2014) The evolution of social bonds in primate males. Behaviour (published online, doi:10.1163/1568539X-00003191)
Ostner J, Nunn CL, Schülke O (2008a) Female reproductive synchrony predicts skewed paternity across primates. Behav Ecol 19:1150–1158
Ostner J, Heistermann M, Schülke O (2008b) Dominance, aggression and physiological stress in wild male Assamese macaques (Macaca assamensis). Horm Behav 54:613–619
Ostner J, Heistermann M, Schülke O (2011) Male competition and its hormonal correlates in Assamese macaques (Macaca assamensis). Horm Behav 59:105–113
Ostner J, Vigilant L, Bhagavatula J, Franz M, Schülke O (2013) Stable heterosexual associations in a promiscuous primate. Anim Behav 86:623–631
Overduin-de Vries AM, Olesen CU, Vries H, Spruijt BM, Sterck EHM (2012) Sneak copulations in long-tailed macaques (Macaca fascicularis): no evidence for tactical deception. Behav Ecol Sociobiol 67:101–111
Pandit S, van Schaik CP (2003) A model for leveling coalitions among primate males: toward a theory of egalitarianism. Behav Ecol Sociobiol 55:161–168
Port M, Kappeler PM (2010) The utility of reproductive skew models in the study of male primates, a critical evaluation. Evol Anthropol 19:46–56
R Development Core Team. 2012 R: A language and environment for statistical computing. Vienna, Austria. http://www.R-project.org/, R Foundation for Statistical Computing
Rodriguez-Llanes JM, Verbeke G, Finlayson C (2009) Reproductive benefits of high social status in male macaques (Macaca). Anim Behav 78:643–649
Say L, Pontier D, Natoli E (2001) Influence of oestrus synchronization on male reproductive success in the domestic cat (Felis catus L.). Proc R Soc Lond B 268:1049–1053
Schielzeth H, Forstmeier W (2009) Conclusions beyond support: overconfident estimates in mixed models. Behav Ecol 20:416–420
Schülke O, Bhagavatula J, Vigilant L, Ostner J (2010) Social bonds enhance reproductive success in male macaques. Curr Biol 20:2207–2210
Schülke O, Heistermann M, Ostner J (2014) Lack of evidence for energetic costs of mate-guarding in wild male Assamese macaques (Macaca assamensis). Int J Primatol. doi:10.1007/s10764-013-9748-y
Setchell J, Charpentier M, Wickings EJ (2005) Mate guarding and paternity in mandrills: factors influencing alpha male monopoly. Anim Behav 70:1105–1120
Sharpe RM (1994) Regulation of spermatogenesis. In: Knobil E, Neil JD (eds) The physiology of reproduction. Raven Press, New York, pp 1363–1434
Snyder-Mackler N, Alberts S, Bergman BJ (2012) Concessions of an alpha male? Cooperative defense and shared paternity in multi-male primate groups. Proc R Soc Lond B 279:3788–3795
Soltis J, Thomsen R, Takenaka O (2001) The interaction of male and female reproductive strategies and paternity in wild Japanese macaques, Macaca fuscata. Anim Behav 62:485–494
Stockley P (2004) Sperm competition in mammals. Hum Fertil 7:91–97
Stumpf RM, Boesch C (2006) The efficacy of female choice in chimpanzees of the Taï Forest, Côte d’Ivoire. Behav Ecol Sociobiol 60:749–765
Tourmente M, Gomendio M, Roldan ERS (2006) Sperm competition and the evolution of sperm design in mammals. BMC Evol Biol 11:12
Trivers RL (1972) Parental investment and sexual selection. In: Campbell B (ed) Sexual selection and the descent of man, 1871-1971. Aldine-Atherton, Chicago, pp 136–179
van Horn RC, Altmann J, Alberts S (2008) Can’t get there from here: inferring kinship from pairwise genetic relatedness. Anim Behav 75:1173–1180
Widdig A, Bercovitch FB, Streich WJ, Sauermann U, Nurnberg P, Krawczak M (2004) A longitudinal analysis of reproductive skew in male rhesus macaques. Proc R Soc Lond B 271:819–826
Willisch CS, Neuhaus P (2010) Social dominance and conflict reduction in rutting male Alpine ibex, Capra ibex. Behav Ecol 21:372–380
Wroblewski EE, Murray CM, Keele BF, Schumacher-Stankey JC, Hahn BH, Pusey A (2009) Male dominance rank and reproductive success in chimpanzees, Pan troglodytes schweinfurthii. Anim Behav 77:873–885
Young C, Hähndel S, Majolo B, Schülke O, Ostner J (2013) Male coalitions, dominance rank and female preferences independently affect male mating success in wild Barbary macaques. Behav Ecol Sociobiol 67:1665–1677
Young C, Schülke O, Ostner J (2014) How males form coalitions against group rivals and the Pandit/van Schaik coalition model. Behaviour. (published online, doi:10.1163/1568539X-00003166)
Acknowledgments
We thank the National Research Council of Thailand (NRCT) and the Department of National Parks, Wildlife and Plant Conservation (DNP) for the permission to conduct this study and for all the support granted. We are grateful to Kitti Kreetiyutanont, Mongkul Kumsuk, Kanjana Nitaya, and Jarupol Prabnasuk (Phu Khieo Wildlife Sanctuary) for their cooperation and permission to carry out this study. We thank A. Koenig and C. Borries (Stony Brook University), who developed the field site at Huai Mai Sot Yai. We thank D. Bootros, N. Bualeng, A. Chunchaen, I. Fürtbauer, M. Heesen, R. Intalo, N. Juntuch, S. Jumrudwong, M. Karlstetter, Thipusa Kilawit, Sally MacDonald, W. Nueorngshiyos, D. Pesek, N. Ponganan, S. Rogahn, P. Saaisawasthikul, M. Swagemakers, B. Whitman, and T. Wisate for their excellent help in the field. We also thank N. Bhumpakphan, W. Eiadthong (Kasetsart University), and W. Brockelman (Mahidol University) for the support and cooperation. We would also like to thank the Faculty of Veterinary Medicine, Kasetsart University, Thailand. We are grateful to Linda Vigilant and Jojo Bhagavatula who established the paternity analyses for this study and thank Chris Young for the help with the statistical analyses. We thank Anja Widdig, Antje Engelhardt, and five anonymous reviewers for helpful comments on different versions of the manuscripts. This research was supported by the Max-Planck Society, the National Geographic Society, and the German Initiative of Excellence through funds to the University of Göttingen as well as by the Center of Excellence on Agricultural Biotechnology, Science and Technology Postgraduate Education and Research Development Office, Office of Higher Education Commission, Ministry of Education (AG-BIO/PERDO-CHE).
Ethical standards
This study was conducted completely noninvasively and under the permission of the authorities of Thailand, i.e., the National Research Council of Thailand (NRCT) and the Department of National Parks, Wildlife and Plant Conservation (DNP) (permits 0004.3/3618 and 0002.3/2647), and we adhered to the Guidelines of the Use of Animals in Research and the guidelines of the involved institutions.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by A. Widdig
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Material Table 1
(DOC 49 kb)
Rights and permissions
About this article
Cite this article
Sukmak, M., Wajjwalku, W., Ostner, J. et al. Dominance rank, female reproductive synchrony, and male reproductive skew in wild Assamese macaques. Behav Ecol Sociobiol 68, 1097–1108 (2014). https://doi.org/10.1007/s00265-014-1721-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-014-1721-z