
Abstract
We examine vigilance within a mixed-species troop of saddleback (Saguinus fuscicollis) and moustached (S. mystax) tamarins over a complete year. Saddleback tamarins were consistently more vigilant than moustached tamarins. This may be linked to their preference for lower strata. In accordance with previous studies of other primates, vigilant tamarins of both species were significantly further away from their nearest neighbours, and were also at lower heights in the forest than non-vigilant individuals. There was no observed sex difference in the amount of time spent vigilant. In terms of modes of scanning, the saddleback tamarins looked up significantly more frequently than the moustached tamarins, whereas there was no difference between the species in the frequency of side sweeps. There were no differences between the sexes in the frequencies of either type of vigilant behaviour. The proportion of time spent vigilant was higher than average immediately prior to entering a sleeping site for saddleback tamarins, but not for moustached tamarins. Both species were more vigilant immediately after exiting a sleeping site than at other times of the day. There was significant variation in the amount of time devoted to vigilance over the course of the year. These findings are discussed with respect to the social structure, ecology and main predator threats facing these species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Predation can act both directly through mortality and also shape ecology and behaviour (Isbell 1994; Lima 1998; Boinski et al. 2000; Treves 2002). Vigilance often serves to detect predators before attack, thus allowing an appropriate anti-predation response (see Elgar 1989; Lima and Dill 1990 for reviews). The link between predation and vigilance is clear. Species respond to an increased risk of predation with increased levels of vigilance (Powell 1974; Lima and Zollner 1996; Hunter and Skinner 1998; Childress and Lung 2003) and vigilant animals have been shown to be safer from predation (FitzGibbon 1989, 1990).
A vigilant animal may be less at risk from predators, but vigilance comes at a cost to other behaviours. Group living has been suggested as an antipredator strategy whereby an animal can potentially reduce both its individual predation risk and its time spent vigilant through two principal mechanisms, the dilution (Hamilton 1971) and detection effects (Pulliam 1973; Lima 1995). Individuals may also benefit from increased predator detection through the greater collective vigilance of the group (Powell 1974; Lazarus 1979; Boland 2003) (but see Fernández et al. 2003). Collective vigilance also allows an individual to reduce the amount of time it spends vigilant, and this has been shown for a range of taxa including birds (Bertram 1980; Pöysä 1994; Boland 2003; Fernández et al. 2003), ungulates (Berger 1978; Alados 1985; FitzGibbon 1990; Hunter and Skinner 1998), macropod marsupials (Heathcote 1987; Blumstein et al. 2001), seals (Krieber and Barrette 1984), sciuromorph rodents (Holmes 1984), lagomorphs (Monaghan and Metcalfe 1985) and primates (Leighton-Shapiro 1986; de Ruiter 1986) (see Elgar 1989 for review). Further, individuals can modify their vigilance based on the behaviour and experience of conspecifics (Griffin and Evans 2003). In addition to the benefits accrued by conspecific groups, species may also benefit from forming mixed-species groups which offer the potential for increased predator detection through complimentary vigilance. This has been suggested for birds (Metcalfe 1984), ungulates (FitzGibbon 1990) and primates (e.g. Pook and Pook 1982; Gautier-Hion et al. 1983).
Following Hamilton (1971), a number of studies have highlighted the need to reconsider the role of group size per se in vigilance and the need to consider other factors, such as the effect of neighbours and inter-individual distance (Pöysä 1987, 1994; Elgar 1989; Catterall et al. 1992; Bekoff 1995; Lima 1995; Roberts 1996; Steenbeek et al. 1999). Indeed, vigilance rates have been found to be lower for individuals with nearer neighbours for an increasingly diverse number of taxa, including birds (Pöysä 1994; Rolando et al. 2001), ungulates (FitzGibbon 1989), macropod marsupials (Blumstein and Daniel 2003), sciuromorph rodents (Holmes 1984), lagomorphs (Roberts 1988) and primates (e.g. Robinson 1981). Although, as Treves (2000) notes, there is no evidence yet that closer individual spacing actually results in reduced predation, despite theoretical grounds for expecting this to be the case (Hamilton 1971).
Like other group-living species, such as ungulates and birds, primates may benefit from collective vigilance and the potential to reduce individual vigilance. Indeed, there is evidence that individuals in larger groups devote less time to vigilance (Leighton-Shapiro 1986; de Ruiter 1986). Individuals with nearer neighbours have also been shown to have lower levels of vigilance (Robinson 1981; van Schaik and van Noordwijk 1989; Cords 1990; Rose and Fedigan 1995; Cowlishaw 1998, 1999; Treves 1998, 1999, 2000; Steenbeek et al. 1999; Rolando et al. 2001; Treves et al. 2001; Hirsch 2002), which may be related to the dilution effect and a lower risk of predation. Primates, like ungulates (Hunter and Skinner 1998; Childress and Lung 2003) and birds (Powell 1974; Lima and Zollner 1996), may alter their vigilance in response to the perceived risk of predation. This can be influenced by many factors, including the density of vegetation which could conceal a predator (Treves 2002).
Within primates, tamarins (Saguinus spp.) are a good model for studies of vigilance. They face a wide range of potential predators (Moynihan 1970; Terborgh 1983; Sussman and Kinzey 1984; Peres 1993) and rely on advanced detection to avoid predation (Peres 1993) since they are not well equipped to retaliate against most of their predators (but see Shahuano et al. 2002). These small-bodied Neotropical primates live in groups, with saddleback tamarins (Saguinus fuscicollis) forming consistent mixed-species troops with either moustached (S. mystax) (Castro and Soini 1978; Heymann 1990), red-bellied (S. labiatus) (Pook and Pook 1982; Buchanan-Smith 1990) or emperor tamarins (S. imperator) (Terborgh 1983) or Snethlage’s marmosets (Callithrix emiliae) (Martins et al. 1987; Lopes and Ferrari 1994) in areas where they are sympatric. Further, a number of authors have commented on the potential for increased predator detection within mixed-species groups of tamarins (see Heymann and Buchanan-Smith 2000 for review). Indeed, it has been shown that saddleback tamarins are more adept at locating terrestrial predators, and moustached tamarins at locating aerial or arboreal predators in the wild (Peres 1993) or objects in captivity (Hardie and Buchanan-Smith 2000).
From what is known about their biology, a number of predictions can be made regarding factors affecting vigilance in saddleback and moustached tamarins. Since they face a potentially high predation risk, and their co-operative social structure reduces the need for social monitoring of conspecifics (Caine and Marra 1988), vigilance in these species is primarily antipredatory in function. As such, it is predicted to decrease with decreasing inter-individual distance if group size is constant (Hirsch 2002). Individuals may be expected to be more vigilant when at higher levels in the forest since diurnal raptors are reported to be their biggest threat, and as such predation risk may be greatest in the upper forest strata (Peres 1993). Moreover, given the well-documented interspecific vertical stratification within mixed-species groups of tamarins (e.g. Yoneda 1981; Terborgh 1983; Buchanan-Smith 1990, 1999; Peres 1993; Smith 1997), it may be expected that saddleback tamarins, found lower in the forest, would be less vigilant than the moustached tamarins and other species with which they associate. Indeed, Peres (1993) and Hardie and Buchanan-Smith (1997) found moustached and red-bellied tamarins respectively were more vigilant than saddleback tamarins. Similarly, Buchanan-Smith and Hardie (1997) reported red-bellied tamarins looked up more often than saddleback tamarins. Consequently, it may also be expected that moustached tamarins would look up with a greater frequency than saddleback tamarins.
Male primates have been reported to devote more time to vigilance than females in a wide range of primate species (Cheney and Seyfarth 1981; Boinski 1988; van Schaik and van Noordwijk 1989; Fragaszy 1990; Koenig 1998; Rose 1998), including tamarins (Buchanan-Smith 1999). Although it has been argued that this may represent investment in social monitoring by males (Cords 1990; Baldellou and Henzi 1992; Rose and Fedigan 1995), this possibility may be reduced in species such as tamarins, where the main function of vigilance is antipredatory (Caine and Marra 1988). Moreover, differences in the visual systems of male and female tamarins and the majority of other Neotropical primates (Mollon et al. 1984) may mean that dichromatic males are not as adept as trichromatic females at detecting some predators, such as felids with an orange/yellow coat against a green leaf background (Coss and Ramakrishnan 2000; Caine 2002). Since an individual can alter its vigilance in response to the ease with which predators can be perceived due to obscuring foliage (Treves 2002), individuals that differ physiologically in their perceptual capabilities may similarly be expected to differ in their vigilance levels. Consequently, male tamarins may be expected to be more vigilant than females.
Tamarins may be at particular risk to predators whilst in or emerging from a sleeping site since, like other callitrichids, they spend over half their lives in them (Yoneda 1981; Buchanan-Smith 1991; Heymann 1995; Smith 1997) and often sleep in enclosed cavities (Heymann 1995; Smith 1997). Further, their level of alertness during the night may be decreased (Moynihan 1970; Hampton 1973; Erkert 1989; Thompson et al. 1994) due to a reduction in their metabolism (Hetherington 1978; Thompson 1991; Schnell and Wood 1993; Thompson et al. 1994). The risk from predation whilst in or entering a sleeping site may be reduced through increased vigilance (Caine 1984), and there is anecdotal evidence of increased vigilance prior to entering a sleeping site by wild tamarins (Dawson 1976; Neyman 1978; Heymann 1995; Smith 1997). This is further supported by captive studies of red-bellied tamarins (Caine 1984, 1987). Similar extra vigilant behaviour may also be expected at dawn when leaving a sleeping site in the morning, since light levels will be low and crepuscular predators may still be active. However, this has not yet been examined.
This paper examines factors affecting vigilance in wild mixed-species troops of saddleback and moustached tamarins. Following the rationale above, five hypotheses are made. Vigilance will increase with distance to nearest neighbour (H1). Vigilance will increase with distance from the ground (H2). Moustached tamarins will be more vigilant than saddleback tamarins (H3). Males will be more vigilant than females (H4). Vigilance immediately prior to entry to, and following exiting from, sleeping sites will be higher than at other times of the day (H5).
Methods
A mixed-species troop of saddleback and moustached tamarins were observed (by A.C.S.) for 112 full days from January 2000 until December 2000 at the Estación Biológica Quebrada Blanco (EBQB) (4°21′S, 73°09′W). The site is located approximately 1 km northwest from the right bank of the Quebrada Blanco in northeastern Peru (for details see Heymann and Hartmann 1991). The annual rainfall at EBQB is 2,740±454 mm (n=5 years). The climate at EBQB can be divided into wet and dry seasons. The wet season, characterized by higher rainfall, runs from February until May and the dry season from June to January. Mean daily maximum and minimum temperatures were 28.5±2.2°C (n=634 days) and 22.1±1.4°C (n=384 days).
At the beginning of the study, the troop consisted of four saddleback tamarins (one adult female, one subadult female, and two adult males) and five moustached tamarins (two adult females, two adult males, and one subadult male). Neither of the female saddleback tamarins was observed to be pregnant during the study, whereas both of the female moustached tamarins gave birth in February. Female 1 was seen with a single infant that died on the same day it was born. Female 2 gave birth to twin males the following day which were raised by both their mother and female 1 (see Smith et al. 2002). Female 2 emigrated on 15 October 2000. The composition of the saddleback tamarin group did not alter over the course of the study. Differences in body size, genital size and colour, ear nicks and the pattern of pelage facilitated the identification of individuals. The tamarins were habituated to the presence of human observers for at least 2 years prior to the reported observations being made.
The tamarins were observed from February until December 2000. They were observed for approximately 14 days each month from February until June, and for 8 days each month from July until December. Each species was the focus of observations for half the time. The tamarins were followed from when they left their sleep tree in the morning until they entered their next sleep tree in the afternoon. The two species spent 93.0% of their active period (n=2,268 scans) over the year in association with each other, i.e. were within 50 m of one another.
Vigilance was examined using two methods, scan samples of all observed animals, and focal animal all-occurrences sampling. For the first method, the behaviour of all visible tamarins of the focal species, whether vigilant or not, was recorded every 30 min, at zero and 30 min past the hour (by A.C.S.). A vigilant animal was stationary and actively attending to the surrounding environment, through observable head and eye movements. The height and distance from each tamarin’s nearest conspecific neighbour were also noted. These data were used to examine the effects of month, species, sex, height, and distance of nearest neighbour on vigilance. For the second method, a single vigilant tamarin was observed for 1 min and the number of times it looked up and swept its head from side to side was noted (by S.K.). Look-ups were defined as the tamarin raising its head in the vertical plane to focus its attention on the space above it, whereas sweeps were defined by head movements in the horizontal plane enabling the tamarin to look left and right. These two component behaviours of vigilance may serve different functions, look-ups to detect aerial predators and sweeps to detect terrestrial or arboreal predators. The tamarin’s height and distance from its nearest conspecific neighbour were noted. Between 18 and 30 focal samples were recorded for each tamarin. The tamarins’ activity period was divided into quarters, and observations were distributed evenly across quarters. On a given day, each animal was sampled no more than once in any quarter. All focal samples were collected in August. These data were used to examine the component behaviours of vigilance, sweeps and look-ups. For both methods, only the distance to the nearest conspecific, as opposed to heterospecific, neighbour was recorded for the sake of simplicity.
Data sets were tested for normality and homogeneity of variance, and analysed using parametric statistics (ANOVA). Data from infants or those that were infants at the beginning of the study were excluded in analyses, as were those from unidentified individuals. Individual identification was possible for 64.5% of the 4,679 scans for the moustached and 83.4% of the 3,602 scans for the saddleback tamarins. An ANOVA with species and month as factors was used to examine the effect of month on the proportion of the daily scans spent vigilant. Since the result of the ANOVA showed a significant effect of month, monthly variation in vigilance and variation in the number of monthly observations were controlled for in subsequent analyses by a repeated measures ANOVA, using monthly mean values for individual tamarins. Differences in the height and nearest-neighbour distance of vigilant and non-vigilant individuals were examined using ANOVAs with species, sex and behaviour (vigilant or not) as factors. Species and sex differences in the two component behaviours of vigilance were examined with a MANOVA, using the number of sweeps and look-ups in focal samples for individual tamarins, with height and nearest-neighbour distance as co-variables.
Results
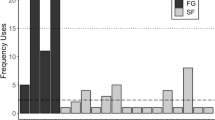
Based on the half-hourly behavioural scans, the proportion of time spent vigilant each month varied significantly over the course of the year (F 10,90 =2.99, P=0.003) (Fig. 1). Saddleback tamarins were significantly more vigilant than moustached tamarins (10.7±7.4% vs 5.8±4.9% scans; F 10,90 =15.17, P<0.001). This contradicts H3. The monthly pattern did not differ between species (F 8,88 =1.71, P=0.090); however, both species were most vigilant in August.

Monthly variation in the proportion of time spent vigilant by saddleback and moustached tamarins
The data were then collapsed and analysed using a repeated measures ANOVA. Saddleback tamarins were more vigilant than moustached tamarins (F 1,4 =34.96, P=0.004) (Fig. 2). There was no effect of sex (F 1,4 =0.04, P=0.852) or the interaction of sex and species (F 1,4 =2.61, P=0.181) on the proportion of time vigilant. That neither sex was more vigilant does not support H4.

The effect of species and sex on the proportion of time spent vigilant (bars indicate SD)
Saddleback tamarins were significantly closer to their nearest conspecific neighbour than moustached tamarins (F 1,2 =170.97, P=0.006) (Fig. 3). Vigilant tamarins were significantly further from their nearest neighbour than their non-vigilant counterparts (F 1,2 =183.61, P=0.005). This supports H1. There was also a significant interaction effect of species and behaviour on nearest-neighbour distance (F 1,2 =41.87, P=0.023). There was no effect of sex on nearest-neighbour distance (F 1,2 =0.001, P=0.987), and nor were the interactions of behaviour and sex (F 1,2 =0.007, P=941), and sex and species (F 1,2 =1.04, P=0.415) significant.

Effect of species, sex and behaviour on mean distance to nearest neighbour during vigilant and non-vigilant behaviour (bars indicate SD)
Moustached tamarins were higher in the forest than saddleback tamarins (F 1,2 =84.83, P=0.012) (Fig. 4). Vigilant tamarins were significantly closer to the ground than their non-vigilant counterparts. (F 1,2 =53.38, P=0.018) (Fig. 4). This contradicts H2. Sex (F 1,2 =0.19, P=0.708), and the interactions of behaviour and sex (F 1,2 =0.13, P=0.785), behaviour and species (F 1,2 =0.27, P=0.655), and sex and species (F 1,2 =0.116, P=0.766), had no significant effect on height in the forest.

Effect of species, sex and behaviour on mean height during vigilant and non-vigilant behaviour (bars indicate SD)
Next, a MANOVA using the data from the focal-animal samples was conducted to examine differences in the two principal vigilance behaviours, look-ups and sweeps. Given the effect of height and distance from nearest-neighbour, mean values for each individual for these were included as covariables. When look-ups were considered, H4 was not supported since males did not look up significantly more than females (F 1,218 =3.37, P=0.068). Saddleback tamarins looked up significantly more often than moustached tamarins (F 1,218 =31.05, P<0.001). This contradicts H3. There was no significant interaction of sex and species (F 1,218 =1.59, P=0.209) (Fig. 5). Height and neighbour distance had no significant effect (F 1,218 =0.37, P=0.543; F 1,218 =0.05, P=0.818). This does not support H1 and H2.

Effect of species and sex on the number of look-ups (bars indicate SD)
Neither species (F 1,218 =0.39, P=0.534) nor sex, (F 1,218 =0.66, P=0.417) nor their interaction (F 1,218 =0.46, P=0.498) had a significant effect on the frequency of sweep scans (Fig. 6). This does not support H3 and H4. Tamarins with further nearest neighbours had a significantly greater frequency of sweep scans (F 1,218 =5.58, P=0.019). Height did not affect the frequency of sweep scans (F 1,218 =0.86, P=0.355). This supports H1 but not H2.

Effect of species and sex on the number of sweeps (bars indicate SD)
The proportion of time spent vigilant in the 10 min immediately before entering and after exiting a sleeping site was compared using t-tests to that at all other times, using the data from the scan samples. Saddleback tamarins were significantly more vigilant both in the first (t 4,4 =29.85, P=0.001) and last (t=−4.51, 3df, P=0.02) 10 min of their activity period than during the rest of the day (Fig. 7). Moustached tamarins were also significantly more vigilant in the first 10 min (t 5,5 =5.79, 4df, P=0.004), but were less vigilant in the last (t=0.85, 4df, P=0.446) but not significantly so. This generally supports H5. This hypothesis was not tested using the focal observations due to sample-size constraints.

Proportion of time spent vigilant during the first and last 10 min of the active period versus that at other times (bars indicate SD)
Discussion
That vigilant tamarins had a further nearest conspecific neighbour supports H1 (vigilance will increase with distance to nearest neighbour) and suggests the function of vigilance is anti-predatory (Hirsch 2002), and is consistent with findings for other primates (e.g. Rose and Fedigan 1995; Treves 1998, 1999; Treves et al. 2001) and other taxa (Holmes 1984; FitzGibbon 1989; Pöysä 1994; Blumstein and Daniel 2003). This increasing evidence has prompted the suggestion that, for primates, nearest-neighbour distance may be a more important factor in determining vigilance than group size (Treves 1998; Treves et al. 2001). However, some studies have not found the same effect. Cords (1990) found that the presence or absence of con- or heterospecifics in a tree did not affect vigilance of blue monkeys (Cercopithecus mitis). Hirsch (2002) found that vigilance increased with increasing numbers of near neighbours in brown capuchins (Cebus apella) although, as he argues, the reason for this latter finding was that in the population under study the main function of vigilance was social monitoring of conspecifics rather than anti-predation. The comparative effects of conspecific and heterospecific neighbours on vigilance would be of interest given that complimentary vigilance has been proposed as a function of these mixed-species groups, and that captive studies have found that the vigilance is altered in the presence or absence of heterospecific congeners (Hardie and Buchanan-Smith 1997), but the data did not permit this analysis.
That vigilant tamarins were lower in the forest, and that saddleback tamarins were significantly more vigilant than moustached tamarins, both in terms of time spent in general vigilance and in the number of look-ups, contradicts H2 (vigilance will increase with distance from the ground) and H3 (moustached tamarins will be more vigilant than saddleback tamarins). Previously reported findings of greater vigilance in moustached (Peres 1993) and red-bellied tamarins (Hardie and Buchanan-Smith 1997) have been explained by these species’ use of higher strata (e.g. Yoneda 1981; Buchanan-Smith 1990; Peres 1991; Smith 1997), it being argued that this would put them at greater risk from raptors, which have been suggested to represent their greatest threat (Peres 1993). Consequently, vigilance would be expected to be greater at higher rather than lower strata. Indeed, Buchanan-Smith (1999) found that red-bellied tamarins detect objects located at higher levels more frequently than saddleback tamarins.
Although the present findings contradict H2 (vigilance will increase with distance from the ground), they are in line with studies of other primate species that show vigilance decreases with increasing height from the ground (de Ruiter 1986; van Schaik and van Noordwijk 1989; Steenbeek et al. 1999; Hirsch 2002). There are two important points to consider. First, there is the possibility that the greater level of vigilance at lower strata was a reaction to the terrestrial human observer, albeit with a non-overt predator response. Indeed, all but exceedingly habituated groups of moustached tamarins are reticent to come to the ground to retrieve flushed prey or to feed on soil in the presence of an observer (A.C.S., personal observation). This factor was minimized in the present study, with both species descending to the ground, on occasions within 5 m of the observer. Moreover, the saddleback tamarins, the more vigilant of the two species, were frequently observed on the ground within 2 m of the observer. Even if an observer has minimal effect on the behaviour of the primate, it would certainly affect that of any terrestrial predator. Consequently, the threat of cats and other terrestrial carnivores to primates may be underestimated (Peres 1993). Second, and perhaps more importantly, the raptors that could take tamarins (e.g. harpy eagle, Harpia harpyja, ornate hawk eagle, Spizaetus ornatus, and crested eagle, Morphnus guianensis) are typically ambush predators, taking their prey from a stationary perch rather than flying over the canopy (Hilty and Brown 1986). A perched raptor will gain most attack velocity if it includes a component of vertical drop. As a consequence, the upper strata may not be as dangerous as first supposed. In fact, the lower strata may be more dangerous since the amount of vertical space above a given monkey in which a swooping raptor can hide and gain attack velocity increases as the monkey’s height in the forest decreases. This is supported by the finding that saddleback tamarins, which occupied lower strata than the moustached tamarins, looked up more often, suggesting a greater investment in detection of predators above them. Individuals at lower heights may also be at greater risk due to their closer proximity to the four, predominantly terrestrial, cat species that may prey on tamarins, namely ocelot (Felis pardalis), margay (F. wiedii), oncilla (F. tigrina), and jaguarundi (F. yagouaroundi) (Emmons and Feer 1990). The hypothesis that the lower levels are more dangerous is supported by the fact that, for other primates, vigilance, and hence perceived predation risk, decrease with increasing height from the forest floor (de Ruiter 1986; van Schaik and van Noordwijk 1989; Steenbeek et al. 1999; Hirsch 2002).
Male tamarins were not significantly more vigilant than females in either species. This does not support H4 (males will be more vigilant than females), although greater levels of male vigilance have been reported in captive studies (e.g. Buchanan-Smith 1999). Factors other than sex per se may play a role. Within tamarins and other Neotropical primates, this may include their sex-linked polymorphic visual system, where heterozygous females are trichromatic and homozygous females, like all males, are dichromatic (Mollon et al. 1984), since it will affect an individual’s ability to perceive predators. Although trichromacy may be advantageous for detecting orange/yellow predators against a leaf background (Coss and Ramakrishnan 2000), dichromacy has been shown to be advantageous in the detection of camouflaged objects where texture is masked by hue (Morgan et al. 1992). This has led to the suggestion that dichromats may be better at detecting cryptic predators (Caine 2002). Given the potential advantages to both di- and trichromats in predator detection, and that not all female tamarins are trichromats, the effects of sex and colour vision status must be examined carefully. The distribution of the visual phenotypes among the study animals did not permit this analysis.
That the variation over the course of the year in the proportion of time spent vigilant each month differed between the two species is surprising given that both were exposed to the same environmental variables. The birth of infants may be expected to raise vigilance levels, but this was not observed for the moustached tamarins, although the saddleback tamarins’ vigilance did increase following the arrival of moustached infants. That vigilance for both species was highest in August may be related to a reduction in fruit resources, a known ecological change at the field site (Tirado Herrera and Heymann, unpublished data), or perhaps an unknown change in the behaviour of the predators. That two observers made observations in this month is unlikely to be a factor, since observers working on different projects were also present in other months.
The increase in vigilance 10 min prior to entry to a sleeping site for saddleback tamarins is in line with captive studies of red-bellied tamarins (Caine 1987) and supports H5 (vigilance immediately prior to entry to, and following exiting from, sleeping sites will be higher than at other times of the day). In contrast, Peres (1993) found a decrease in vigilance in the hour before entry to sleeping sites for saddleback tamarins, although he notes that the tamarins were more sensitive to disturbance and potential threats at this time. The difference in the pre-retirement period analysed (1 h vs 10 min) may be key to these differences. Peres noted that social behaviours increased before retirement. A similar increase was noted in the present study, but the increased social grooming is terminated before the tamarins make a more vigilant progression to their sleeping site during the last 10 min. The interspecific difference, with the moustached tamarins showing a decrease in vigilance, may be linked to their respective modes of progression to sleeping sites. In comparison to saddleback tamarins which typically follow a steady cling-and-leap progression at relatively low heights with frequent and occasionally long pauses, moustached tamarins progress quadrupedally, at a faster rate using the upper strata.
The high level of vigilance in both species after leaving the sleeping sites may reflect the need to visually check the surroundings for predators before progressing to the day’s first fruit tree. Moreover, since the tamarins exit their sleeping trees within a few minutes of sunrise (Smith 1997), the light level in the forest is low, reducing visual acuity, and predators may be more difficult to detect. Tamarins have been found to alarm call to either real or perceived threats more frequently in the first hours of the day (Peres 1991; Smith 1997). This supports the finding of the present study of a greater perceived risk of predation early in the day, which may be associated with low light levels and/or the activity cycles of the predators.
References
Alados CL (1985) An analysis of vigilance in the Spanish ibex (Capra pyrenaica). Z Tierpsychol 68:58–64
Baldellou M, Henzi SP (1992) Vigilance, predator detection and the presence of supernumerary males in vervet monkey troops. Anim Behav 43:451–461
Bekoff M (1995) Vigilance, flock size, and flock geometry: information gathering by western evening grossbeaks (Aves, Fringilidae). Ethology 99:150–161
Berger J (1978) Group size, foraging and antipredation ploys: an analysis of bighorn sheep decisions. Behav Ecol Sociobiol 4:91–99
Bertram BCR (1980) Vigilance and group size in ostriches. Anim Behav 28:278
Blumstein DT, Daniel JC (2003) Red kangaroos (Macropus rufus) receive an antipredator benefit from aggregation. Acta Ethol 5:95–99
Blumstein DT, Daniel JC, Evans CS (2001) Yellow-footed rock-wallaby (Petrogale xanthopus) group size effects reflect a trade-off. Ethology 107:655–664
Boinski S (1988) Sex differences in the foraging behavior of squirrel monkeys in a seasonal habitat. Behav Ecol Sociobiol 23:177–186
Boinski S, Treves A, Chapman CA (2000) A critical evaluation of the influence of predators on primates: effects on group travel. In: Boinski S, Garber PA (eds) On the move: how and why animals travel in groups. University of Chicago Press, Chicago, pp 43–72
Boland CRJ (2003) An experimental test of predator detection rates using groups of free-living emus. Ethology 109:209–222
Buchanan-Smith HM (1990) Polyspecific association of two tamarin species, Saguinus labiatus and Saguinus fuscicollis, in Bolivia. Am J Primatol 22:205–214
Buchanan-Smith HM (1991) A field study on the red-bellied tamarin, Saguinus l. labiatus, in Bolivia. Int J Primatol 12:259–276
Buchanan-Smith HM (1999) Exploration of unfamiliar areas and detection of potentially threatening objects in single- and mixed-species groups of tamarins. Int J Comp Psychol 12:2–20
Buchanan-Smith HM, Hardie SM (1997) Tamarin mixed-species groups: an evaluation of a combined captive and field approach. Folia Primatol 68:272–286
Caine NG (1984) Visual scanning by tamarins: a description of the behaviour and tests of two derived hypotheses. Folia Primatol 43:59–67
Caine NG (1987) Vigilance, vocalizations, and cryptic behaviour at retirement in captive groups of red-bellied tamarins, Saguinus labiatus. Am J Primatol 12:241–250
Caine NG (2002) Seeing red: consequences of individual differences in color vision in callitrichid primates. In: Miller LE (ed) Eat or be eaten: predator sensitive foraging among primates. Cambridge University Press, New York, pp 58–73
Caine NG, Marra SL (1988) Vigilance and social organization in two species of primates. Anim Behav 36:897–904
Castro R, Soini P (1978) Field studies on Saguinus mystax and other callitrichids in Amazonian Peru. In: Kleiman DG (ed) The biology and conservation of the Callitrichidae. Smithsonian Institution Press, Washington DC, pp 73–78
Catterall CP, Elgar MA, Kikkawa J (1992) Vigilance does not covary with group size in an island population of silvereyes (Zosterops lateralis). Behav Ecol 3:207–210
Cheney DL, Seyfarth RM (1981) Selective forces affecting the predator alarm calls of vervet monkeys. Behaviour 76:25–61
Childress MJ, Lung MA (2003) Predation risk, gender and the group size effect: does elk vigilance depend upon the behaviour of conspecifics? Anim Behav 66:389–398
Cords M (1990) Vigilance and mixed-species association of some east African forest monkeys. Behav Ecol Sociobiol 26:297–300
Coss RG, Ramakrishnan U (2000) Perceptual aspects of leopard recognition by wild bonnet macaques (Macaca radiata). Behaviour 137:315–335
Cowlishaw G (1998) The role of vigilance in the survival and reproductive strategies of desert baboons. Behaviour 135:431–452
Cowlishaw G (1999) Ecological and social determinants of spacing behaviour in desert baboon groups. Behav Ecol Sociobiol 45:67–77
Dawson GA (1976) Behavioural ecology of the Panamanian tamarin, Saguinus oedipus geoffroyi (Callitrichidae, Primates). PhD Thesis, Michigan State University, East Lansing
Elgar MA (1989) Predator vigilance and group size in mammals and birds: a critical review of the empirical evidence. Biol Rev 64:13–33
Emmons LH, Feer F (1990) Neotropical rainforest mammals: a field guide. University of Chicago Press, Chicago
Erkert HG (1989) Characteristics of the circadian activity rhythm in common marmosets (Callithrix j. jacchus). Am J Primatol 17:271–286
Fernández GJ, Capurro AF, Reboreda JC (2003) Effect of group size on individual and collective vigilance in greater rheas. Ethology 109:413–425
FitzGibbon CD (1989) A cost to individuals with reduced vigilance in groups of Thomson’s gazelles hunted by cheetahs. Anim Behav 37:508–510
FitzGibbon CD (1990) Mixed-species grouping in Thomson’s gazelles: the antipredator benefits. Anim Behav 39:1116–1126
Fragaszy DM (1990) Age and sex differences in the organization of behaviour in wedge-capped capuchins (Cebus olivaceus). Behav Ecol 1:81–94
Gautier-Hion A, Quris R, Gautier JP (1983) Monospecific vs. polyspecific life: a comparative study of foraging and antipredatory tactics in a community of Cercopithecus monkeys. Behav Ecol Sociobiol 12:325–335
Griffin AS, Evans CS (2003) Social learning of antipredator behaviour in a marsupial. Anim Behav 66:485–492
Hamilton WD (1971) Geometry for the selfish herd. J Theor Biol 31:295–311
Hampton JK Jr (1973) Diurnal heart rate and body temperature in marmosets. Am J Phys Anthropol 38:339–342
Hardie SM, Buchanan-Smith HM (1997) Vigilance in single- and mixed-species tamarin groups (Saguinus labiatus and Saguinus fuscicollis). Int J Primatol 18:217–234
Hardie SM, Buchanan-Smith HM (2000) Responses of single- and mixed-species groups of Saguinus to novel non-threatening objects. Int J Primatol 21:629–648
Heathcote CF (1987) Grouping of eastern grey kangaroos in open habitat. Aust Wildl Res 14:343–348
Hetherington CM (1978) Circadian oscillation of body temperature in the marmoset, Callithrix jacchus. Lab Anim 12:107–108
Heymann EW (1990) Interspecific relations in a mixed- species troop of moustached tamarins, Saguinus mystax, and saddle-back tamarins, Saguinus fuscicollis (Platyrrhini: Callitrichidae) at the Rio Blanco, Peruvian Amazonia. Am J Primatol 21:115–127
Heymann EW (1995) Sleeping habits of tamarins, Saguinus mystax and Saguinus fuscicollis (Mammalia; Primates; Callitrichidae), in north-eastern Peru. J Zool Lond 237:211–226
Heymann EW, Buchanan-Smith HM (2000) The behavioural ecology of mixed-species troops of callitrichine primates. Biol Rev Cam Philos Soc 75:169–190
Heymann EW, Hartmann G (1991) Geophagy in mustached tamarins, Saguinus mystax (Platyrrhini: Callitrichidae), at the Rio Blanco, Peruvian Amazonia. Primates 32:533–537
Hilty SL, Brown WL (1986) A guide to the birds of Colombia. Princeton University Press, Princeton
Hirsch BT (2002) Social monitoring and vigilance behavior in brown capuchin monkeys (Cebus apella). Behav Ecol Sociobiol 52:458–464
Holmes WG (1984) Predation risk and foraging in the hoary marmot in Alaska. Behav Ecol Sociobiol 15:293–301
Hunter LTB, Skinner JD (1998) Vigilance behaviour in African ungulates: the role of predation pressure. Behaviour 135:195–211
Isbell LA (1994) Predation on primates: ecological patterns and evolutionary consequences. Evol Anthropol 3:61–71
Koenig A (1998) Visual scanning by common marmosets (Callithrix jacchus): functional aspects and the special role of adult males. Primates 39:85–90
Krieber M, Barrette C (1984) Aggregation behaviour of harbour seals at Forillon National Park, Canada. J Anim Ecol 53:913–928
Lazarus J (1979) The early warning function of flocking in birds: an experimental study with captive quelea. Anim Behav 27:855–865
Leighton-Shapiro ME (1986) Vigilance and the costs of motherhood to rhesus monkeys. Am J Primatol 10:414–415
Lima SL (1995) Back to the basics of anti-predatory vigilance: the group size effect. Anim Behav 49:11–20
Lima SL (1998) Non-lethal effects in the ecology of predator-prey interactions. BioScience 48:25–34
Lima SL, Dill LM (1990) Behavioural decisions made under the risk of predation: a review and prospectus. Can J Zool 68:619–640
Lima SL, Zollner PA (1996) Anti-predatory vigilance and the limit to collective detection: visual and spatial separation between foragers. Behav Ecol Sociobiol 38:355–363
Lopes MA, Ferrari SF (1994) Foraging behaviour of a tamarin group (Saguinus fuscicollis weddelli) and interactions with marmosets (Callithrix emiliae). Int J Primatol 15:373–387
Martins E, Schneider H, Leao VF (1987) Syntopy and troops association between Callithrix and Saguinus from Rondonia, Brazil. Int J Primatol 8:57
Metcalfe NB (1984) The effects of mixed-species flocking on the vigilance of shorebirds: who do they trust? Anim Behav 32:986–993
Mollon JD, Bowmaker JK, Jacobs GH (1984) Variations of color vision in a New World primate can be explained by polymorphism of retinal photopigments. Proc R Soc Lond B 222:373–399
Monaghan P, Metcalfe NB (1985) Group foraging in wild brown hares: effects of resource distribution and social status. Anim Behav 33:993–999
Morgan MJ, Adam A, Mollon JD (1992) Dicromats detect color-camouflaged objects that are not detected by trichromats. Proc R Soc Lond B 248:291–295
Moynihan M (1970) Some behavioural patterns of platyrrhine monkeys. II. Saguinus geoffroyi and some other tamarins. Smithsonian Contrib Zool 28:1–77
Neyman PF (1978) Aspects of the ecology and social organisation of free ranging cotton-top tamarins (Saguinus oedipus) and the conservation status of the species. In: Kleiman DG (ed) The biology and conservation of the Callitrichidae. Smithsonian Institution, Washington DC, pp 39–72
Peres CA (1991) Ecology of mixed-species groups of tamarins in Amazonian terra firme forests. PhD Thesis, University of Cambridge
Peres CA (1993) Anti-predation benefits in a mixed-species group of amazonian tamarins. Folia Primatol 61:61–76
Pook A, Pook AG (1982) Polyspecific association between Saguinus fuscicollis and Saguinus labiatus, Callimico goeldii and other primates in north western Bolivia. Folia Primatol 38:196–221
Powell GVN (1974) Experimental analysis of the social value of flocking by starlings (Sturnus vulgaris) in relation to predation and foraging. Anim Behav 22:501–505
Pöysä H (1987) Feeding-vigilance trade-off in the teal (Anas crecca): effects of feeding method and predation risk. Behaviour 103:108–121
Pöysä H (1994) Group foraging, distance to cover and vigilance in the teal, Anas crecca. Anim Behav 48:921–928
Pulliam HR (1973) On the advantages of flocking. J Theor Biol 38:419–422
Roberts G (1996) Why individual vigilance declines as group size increases. Anim Behav 51:1077–1086
Roberts SC (1988) Social influences on vigilance in rabbits. Anim Behav 36:905–913
Robinson JG (1981) Spatial structure in foraging groups of wedge-capped capuchin monkeys Cebus nigrivittatus. Anim Behav 29:1036–1056
Rolando A, Caldoni R, De Sanctis A, Laiolo P (2001) Vigilance and neighbour distance in foraging flocks of choughs, Pyrrhocorax pyrrhocorax. J Zool Lond 253:225–232
Rose LM (1998) Behavioural ecology of white-faced capuchins (Cebus capucinus) in Costa Rica. PhD Thesis, Washington University, St. Louis, Miss
Rose LM, Fedigan LM (1995) Vigilance in white-faced capuchins, Cebus capucinus, in Costa Rica. Anim Behav 49:63–70
Ruiter JR de (1986) The influence of group size on predator scanning and foraging behaviour of wedgecapped capuchin monkeys (Cebus olivaceus). Behaviour 98:240–258
Schaik CP van, Noordwijk MA van (1989) The special role of male Cebus monkeys in predation avoidance and its effect on group composition. Behav Ecol Sociobiol 24:265–276
Schnell CR, Wood JM (1993) Measurement of blood pressure and heart rate by telemetry in conscious, unrestrained marmosets. Am J Physiol 264:1509–1516
Shahuano TN, Huck M, Heymann EW (2002) Boa constrictor attack and successful group defence in moustached tamarins, Saguinus mystax. Folia Primatol 73:146–148
Smith AC (1997) Comparative ecology of saddleback (Saguinus fuscicollis) and moustached (Saguinus mystax) tamarins. PhD Thesis, University of Reading
Smith AC, Tirado ER, Buchanan-Smith HM, Heymann EW (2002) Multiple breeding females and allo-nursing in a wild group of moustached tamarins (Saguinus mystax). Neotrop Primates 9:67–69
Steenbeek R, Piek RC, van Buul M, van Hooff JARAM (1999) Vigilance in wild Thomas’s langurs (Presbytis thomasi): the importance of infanticide risk. Behav Ecol Sociobiol 45:137–150
Sussman RW, Kinzey WG (1984) The ecological role of the Callitrichidae: a review. Am J Phys Anthropol 64:419–449
Terborgh J (1983) Five new world primates. A study in comparative ecology. Princeton University Press, Princeton
Thompson S (1991) Biotelemetric studies of mammalian thermoregulation. In: Asa C (ed) Biotelemetry applications for captive animal care and research. American Association of Zoological Parks and Aquariums, Wheeling, pp 19–28
Thompson SD, Power ML, Rutledge CE, Kleiman DG (1994) Energy metabolism and thermoregulation in the golden lion tamarin (Leontopithecus rosalia). Folia Primatol 63:131–143
Treves A (1998) The influence of group size and neighbors on vigilance in two species of arboreal monkeys. Behaviour 135:453–481
Treves A (1999) Within-group vigilance in red colobus and redtail monkeys. Am J Primatol 48:113–126
Treves A (2000) Theory and method in studies of vigilance and aggregation. Anim Behav 60:711–722
Treves A (2002) Predicting predation risk for foraging, arboreal monkeys. In: Miller LE (ed) Eat or be eaten: predator sensitive foraging among primates. Cambridge University Press, New York, pp 222–241
Treves A, Drescher A, Ingrisano N (2001) Vigilance and aggregation in black howler monkeys (Alouatta pigra). Behav Ecol Sociobiol 50:90–95
Yoneda M (1981) Ecological studies of Saguinus fuscicollis and S. labiatus with reference to habitat segregation and height preference. Kyoto Univ Overseas Rep 2:43–50
Acknowledgements
We are grateful to Dr. E. Montoya of the Proyecto Peruano de Primatologia and Biologo R. Pezo of the Universidad Nacional de la Amazonia Peruana (UNAP), Iquitos, Peru for help and support with logistical matters. Particular thanks are due to Ney Shahuano who provided unflagging assistance in the field. We thank A. Rennie for her comments on an early draft of the manuscript. This study was funded by the BBSRC (98/S11498 to H.M.B.-S.).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by D. Watts
Rights and permissions
About this article
Cite this article
Smith, A.C., Kelez, S. & Buchanan-Smith, H.M. Factors affecting vigilance within wild mixed-species troops of saddleback (Saguinus fuscicollis) and moustached tamarins (S. mystax). Behav Ecol Sociobiol 56, 18–25 (2004). https://doi.org/10.1007/s00265-003-0753-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-003-0753-6