
Abstract
Field observations and model-presentation experiments have shown that yellow warblers (Dendroica petechia) produce “seet” calls preferentially in response to brood-parasitic brown-headed cowbirds (Molothrus ater). In this study, we investigated whether seet calls are functionally referential alarm calls denoting cowbirds by determining whether female warblers responded appropriately to seet calls in the absence of a cowbird, whether alarm calling by warblers varied with response urgency, and how warblers in a population allopatric with cowbirds responded to cowbird and avian predator models and seet playbacks. As a control, we presented “chip” calls, which are elicited by nest predators as well as by non-threatening intruders, but are not strongly associated with cowbirds. Yellow warblers responded differently to playbacks of seet than chip calls. To seet playbacks, almost 60% of females gave seet calls and rushed to sit in their nests, responses typically elicited by cowbirds, whereas these responses were given infrequently in response to chip calls. Yellow warblers seet called equally in situations that simulated low, medium and high risk of parasitism, which suggests that these calls did not vary with response urgency. In a population allopatric with cowbirds, seet calls were rarely produced in response to cowbird or avian nest predator models and never to seet playbacks. These results suggest that seet calls are functionally referential signals denoting cowbirds and that cowbird parasitism was a strong selective pressure in the evolution of functional referentiality in the seet call of yellow warblers.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In the presence of predators, animals produce complex and diverse alarm signals. In primates and some birds, the acoustic structure of alarm calls varies predictably with predator type and receivers respond with appropriate anti-predator behaviours to these calls in the absence of the eliciting stimuli (e.g. Seyfarth et al. 1980; Evans et al. 1993; Zuberbühler et al. 1997). These calls are termed functionally referential as they apparently convey information about the type of predator encountered (reviewed in Evans 1997). To be classified as functionally referential, alarm calls must display three features. First, the calls should be acoustically distinct from other vocalisations in the species’ repertoire (Marler et al. 1992; Evans 1997). Second, alarm calls should be preferentially produced during interactions with a particular type of predator and be infrequently elicited in other contexts; that is, they should display high degrees of production specificity (Marler et al. 1992; Macedonia and Evans 1993). Finally, anti-predator responses of receivers should be elicited by the alarm calls in the absence of visual cues produced by the stimulus or conspecifics; that is, signals should exhibit perception specificity (Marler et al. 1992; Macedonia and Evans 1993).
In contrast to the functionally referential calls of primates, the alarm calls of rodents convey the immediacy of response required to escape predation, by varying with the mode of hunting, speed of approach, and distance of the predator, but they are not highly predator-specific (e.g. Leger et al. 1979; Macedonia and Evans 1993; Blumstein 1995; Le Roux et al. 2001; Warkentin et al. 2001; see also, e.g., Ficken 1990; Naguib et al. 1999 for avian examples). Similar calls are produced in response to aerial and ground predators, with individuals varying call rate and call structure depending on predation risk (e.g. Leger et al. 1979; Blumstein and Armitage 1997). Few studies have examined whether the acoustic structure of alarm calls varies simultaneously with response urgency and predator type. In ringtailed lemurs (Lemur catta), response urgency does not influence which alarm call is produced (Pereira and Macedonia 1991), whereas structural variation in alarm calls conveys information about response urgency and predator type in suricates (Suricata suricatta: Manser 2001; Manser et al. 2001) and possibly Diana monkeys (Cercopithecus diana: Zuberbühler 2000).
The use of divergent strategies to escape predators with different hunting modes may favour the evolution of functionally referential alarm calls (Macedonia and Evans 1993). Vervet monkeys (C. aethiops), for instance, use multiple escape strategies, climbing trees to escape leopards and moving out of trees and into dense brush to escape eagles, and have evolved different alarm calls for these two predator classes (Seyfarth et al. 1980). In the case of ground squirrels and their relatives, the only mode of escape is to run into burrows (e.g. Leger et al. 1979; Blumstein 1995; Le Roux et al. 2001), and selection has favoured alarm calls that communicate the urgency of response rather than predator identity. In suricates, there is additional complexity as high degrees of sociality favour maintaining group cohesion and coordinating vigilance under predation pressure, tasks best accomplished by communicating intruder type as well as response urgency (Manser et al. 2001). Comparative studies of populations and species that vary in exposure to different predators are needed to understand fully the evolution of functionally referential systems (Evans 1997), but these are generally lacking (but see Macedonia 1990; Fichtel and Kappeler 2002).
Most observations of functionally referential alarm calls have been made in primates (e.g. Seyfarth et al. 1980; Macedonia 1990; Zuberbühler et al. 1997), although recent evidence suggests the existence of such calls in some bird species (Seyfarth and Cheney 1990; Evans et al. 1993; Regosin 2002; Seddon et al. 2002). In particular, studies on nest defence by yellow warblers (Dendroica petechia) have provided evidence for two of the three criteria for functional reference, namely acoustical distinctiveness and production specificity. Yellow warblers are small (10 g), socially monogamous passerines (Yezerinac et al. 1997) that are common hosts of brood-parasitic brown-headed cowbirds (Molothrus ater). Cowbirds lay eggs in the nests of hosts, which raise the cowbird chick often to the detriment of their own young (Ortega 1998; Lorenzana and Sealy 1999). Yellow warblers produce acoustically distinct alarm calls, termed “seet” calls, when confronting live and model cowbirds around the nest (Hobson and Sealy 1989a; Gill 1995; Sealy et al. 1995; Gill and Sealy 1996) and to playbacks of male and female cowbird vocalisations (Gill et al. 1997). The responses of yellow warblers to cowbirds vary between nesting stages, as seet calls are given much more frequently during the egg-laying period, when nests are vulnerable to cowbird parasitism, than during the nestling period (Hobson and Sealy 1989a; Gill and Sealy 1996), when cowbirds typically pose no threat to warbler nests (McLaren and Sealy 2000). Moreover, yellow warblers in a population not exposed to cowbirds rarely produced seet calls in response to cowbird models (Briskie et al. 1992). By contrast, yellow warblers typically emit “chip” calls, and less frequently “metallic chip” and “warble” calls (Gill 1995), to other intruders, including conspecifics, non-threatening species such as sparrows, avian and mammalian nest predators, and predators of adult birds such as hawks (Ficken and Ficken 1965; Reid and Sealy 1986; Studd and Robertson 1985; Hobson et al. 1988; Hobson and Sealy 1989a, 1989b; Sealy 1994; Gill 1995; Gill and Sealy 1996). These studies suggest that yellow warbler seet calls may be functionally referential as they are acoustically distinct from other alarm calls and they are preferentially produced during interactions with cowbirds.
In this study, we used a combination of model-presentation and playback experiments in populations sympatric and allopatric with cowbirds to investigate whether yellow warbler seet calls are functionally referential alarm calls denoting cowbirds, and to examine the evolution of functional reference during nest defence. First, we performed playback experiments to examine the perception specificity of seet calls, that is, whether playbacks of seet calls elicit anti-parasite behaviours in receivers. We used nest-protection behaviour as a behavioural assay in this experiment. Nest-protection behaviour is a striking defensive response in which females, upon sighting a cowbird, fly quickly to and sit tightly in their nests, typically while uttering seet calls (Hobson and Sealy 1989a; Gill and Sealy 1996). If seet calls are perceived as denoting cowbirds, then yellow warblers should respond with nest-protection behaviour to seet call playbacks, supporting the perception-specificity requirement. Second, we examined whether response urgency influences the use of seet calls by yellow warblers by presenting cowbird models in positions that simulate high-, medium- and low-risk of brood parasitism. Previous experiments have placed cowbird models close to the nest mimicking only high-risk situations (e.g. Hobson and Sealy 1989a; Gill and Sealy 1996), so how yellow warblers respond to cowbird models presenting a less immediate threat is unknown. If yellow warblers produce seet calls only in high-risk situations, this suggests that response urgency also may be conveyed. Finally, we examined the responses to cowbird and avian nest predator models and seet call playbacks of yellow warblers in a population allopatric with cowbirds. In an earlier study, Briskie et al. (1992) showed that yellow warblers allopatric with cowbirds rarely gave seet calls when presented with cowbird models. However, how these warblers respond to nest predators is unknown and it is possible that they give seet calls when confronting nest predators rather than cowbirds in this population. If so, then seet calls may have a different referent in populations that are not parasitized by cowbirds.
Methods
Study sites
We conducted this research from May to July 1993 and 1994 at Delta Marsh (50°11′N, 98°19′W) and June 1994 at Churchill (58°40′N, 94°25′W), Manitoba, Canada. About 21% (Sealy 1995) of yellow warbler nests were parasitised by brown-headed cowbirds in the forested dune ridge at Delta Marsh (hereafter the sympatric population), which separated the southern end of Lake Manitoba from the surrounding marsh. The Churchill site (hereafter the allopatric population) was located at the northern edge of the boreal tree line and consisted of areas of willow-birch thickets along lake and pond shores. Cowbirds are vagrants in the Churchill area and have not been confirmed to breed there (Briskie et al. 1992). At both sites, nests were located at all nesting stages and monitored until the young fledged or the nest failed. We randomly selected nests for use in playback and model-presentation experiments and conducted all experiments between 0500 and 1800 hours Central Standard Time.
Responses to playbacks and models
Yellow warblers respond with more than 10 different behaviours to models of cowbirds and nest predators (e.g. Gill and Sealy 1996). For the purposes of this study, we quantified the intensity with which females performed nest-protection behaviour, seet calls, and chip calls, which have been shown in previous studies to be produced with differing frequency to cowbirds versus other nest threats (Hobson and Sealy 1989a; Gill and Sealy 1996). Spectrograms of typical seet and chip calls are given in Hobson and Sealy (1989a) and Gill and Sealy (2003). Nest-protection behaviour was quantified as the time (in seconds) that females sat in their nests to a maximum of 60 s (see below), and seet and chip calls as the actual number produced during the trials. Although males give seet calls to cowbirds, they do not perform nest-protection behaviour (Hobson and Sealy 1989a; Gill and Sealy 1996); thus, females were the test subjects. S.A.G. recorded all observations on audiocassettes and transcribed them later.
Playback experiments
Playback stimuli
Playbacks were used to test the perception specificity of seet calls, that is, whether receivers extract enough information from the playbacks alone to respond as though a cowbird was present. We did not know whether birds would discriminate between the calls of unfamiliar versus known individuals. Therefore, we presented seet and chip calls recorded from three unfamiliar birds (hereafter stranger calls) and from the test subjects themselves (hereafter self calls) in the sympatric population. Test subjects would have had little or no experience with the unfamiliar birds, as they nested approximately 1.5 km away from the females used in the experiments. Not all test subjects vocalised in response to models (see below), and thus the sample size of self call playbacks unavoidably varied between call types and nesting stages (laying: seet calls, n=22, chip calls, n=18; nestling stage: seet calls, n=9, chip calls, n=19). Twenty-six and 23 subjects at laying and nestling, respectively, were played tapes of the stranger calls (n=3) in the sympatric population. In the allopatric population, we played only one tape of each call to 15 subjects; thus, playbacks of additional exemplars are warranted.
Using a Uher 4000 Report-L recorder and Sennheiser ME 88 directional microphone, we recorded seet and chip calls given by yellow warblers when presented with models of a female cowbird and a female common grackle (Quiscalus quiscula), respectively (see Gill and Sealy 1996). From these recordings, we selected sections of alarm calling of good quality with low background noise from which we produced 1-min playbacks. We limited our playbacks to 1 min because receivers may habituate to longer playbacks (Falls 1982) and parasitism and predation events occur within 1–2 min (Sealy 1994; Sealy et al. 1995). A potential weakness in our playback experiments was that we did not standardise call rate among playbacks. We examined the effect of playback call rate on response intensity by performing Spearman rank correlations on the self-call data sets and Kruskal-Wallis tests on the stranger call data set, with playback number (n=3) as the grouping variable. Call rate apparently had little effect on female responsiveness, as only the rate during self-seet playbacks was positively correlated with the duration of nest-protection behaviour at the laying stage (r s=0.476, n=22, P=0.03) but not the number of seet calls elicited (r s=0.304, n=22, P=0.16). None of the other 11 comparisons was significant. These results suggest that call rate may affect female responsiveness and indicate that further experiments are warranted to clarify the role of call rate.
Experimental procedures
Each pair was tested on the same day with all playback stimuli in randomly determined order. We performed both seet and chip playbacks at a given nest on the same day because high predation rates on yellow warbler nests would have resulted in the loss of almost 40% of nests by the second day (Gill and Sealy, unpublished data). At least 15 min prior to initiating the playback series, we placed an Audio-Technica amplified speaker 1 m below the focal nest and set up a blind from which observations were made. We connected the speaker by a 10-m cord to a Sony TCM-5000EV recorder from which we played the calls. We adjusted the volume of the calls by ear to the amplitude observed for calls given during model-presentation experiments and during naturally occurring encounters. Playbacks were initiated when both the male and female were out of the immediate nesting area (>10 m from the nest), and we waited at least 10 min before presenting the next playback, as established in model-presentation protocols (Sealy et al. 1998).
Model-presentation experiments
Test of response urgency in the sympatric population
To examine whether seet calls convey response urgency, we presented in random order a single taxidermic model of a female cowbird at three distances (0.5, 2.5 and 4.5 m) from 27 nests during the egg-laying stage. We ensured that the model was equally visible from the nest in the forested habitat by placing the model in areas of low vegetation density where possible and by tying back vegetation that may have obscured the model. We assumed these distances represented different risks to nesting females, but the distances chosen necessarily reflect a compromise between visibility to the test subjects and variation in risk. The model was clipped to vegetation or positioned on a pole, level with and facing the nest, when the female was >5 m from her nest. When models were presented on poles, we positioned the poles 15 min prior to the first trial and repositioned them at the end of each trial. The trial began when the female arrived within 5 m of the nest and subsequent trials were separated by at least 15 min. Stimuli were presented for 5 min, but we present data from the first minute only to permit comparison with playback results. In general, most females gave seet calls and rapidly returned to their nests in the first minute, and sat silently in their nests for the remainder of the experiments.
Responses to cowbirds and avian nest predator models in the allopatric population
We tested whether yellow warblers in a population allopatric with cowbirds gave seet calls when confronting a nest predator, and thus whether seet calls have a different referent in a population not exposed to cowbird parasitism. We presented models of a cowbird, an avian nest predator (gray jay Perisoreus canadensis), and a non-threatening control (fox sparrow Passerella iliaca) to yellow warblers during their egg-laying period. A model of a gray jay was used in these experiments because they are the smallest avian nest predator in the Churchill area, and grackles are uncommon (Godfrey 1986). Models were presented in random order 0.5 m from focal nests; other elements of this protocol were as above.
Statistical analyses
We used non-parametric procedures because the data were not normally distributed. To analyse the influence of familiarity (self vs stranger calls) and playback call type (seet vs chip) on the intensity of female responses at each nesting stage, we used Wilcoxon signed-ranks tests. We used Wilcoxon two-sample tests to examine the influence of nesting stage (laying vs nestling) and exposure to cowbirds (sympatric vs allopatric) on female responses. Friedman tests were used to analyse the responses to the three models in the response-urgency and Churchill experiments. Analyses were performed using Statview v.4.5. Values are given as mean ± standard error.
Results
Responses to seet calls in the sympatric population
Comparison of self versus stranger calls
The responses of female yellow warblers to seet and chip playbacks did not differ significantly depending upon whether playbacks consisted of the subject’s own calls or those from unfamiliar individuals (Table 1). Females gave a similar number of seet calls and performed nest-protection behaviour for a similar duration to self- and stranger-seet playbacks. Although the mean number of chip calls given during the self-seet playback was more than double than for stranger-seet playback, the difference was not significant. Both seet calls and nest-protection behaviour were infrequently given in response to either self-chip or stranger-chip playback, and the number of chip calls given by females in response to self-chip playbacks was not significantly different from the number given to stranger-chip playbacks. These results suggest that female yellow warblers did not discriminate between their own calls and those of unfamiliar birds. Because sample sizes for playbacks of stranger’s calls were greater than for self calls, results in the remaining sections are drawn from stranger-call playbacks. Results from self calls were qualitatively similar to the results presented here.
Laying stage
For seet calls to satisfy the requirement of perception specificity, playbacks of seet calls should be sufficient to elicit nest-protection behaviour from receivers. Female yellow warblers responded differently to playbacks of seet and chip calls at the laying stage. Most females promptly returned to their nest areas when playbacks were performed, and this was striking for seet playbacks, as females typically stopped foraging within the first few seconds of the playback, and flew immediately to their nests. When chip calls were played, females also stopped foraging, but typically remained 2–5 m from the nest while changing perches, chip calling, and remaining alert. Almost 60% of females performed nest-protection behaviour and gave seet calls during seet playbacks, whereas only one female sat in the nest and two females gave seet calls in response to chip playbacks. Females gave significantly more seet calls (Fig. 1a; Wilcoxon signed-ranks test, Z=3.233, n=26, P=0.001) and sat in the nest significantly longer (Fig. 2; Z=3.010, n=26, P=0.003) in response to seet-call playbacks than chip-call playbacks. There was no difference between playbacks in the number of chip calls given by females (Fig. 1a; Z=1.131, n=26, P>0.2).

Number (mean±SE) of seet and chip calls given by female yellow warblers (Dendroica petechia) in response to seet and chip playbacks in the sympatric population during a egg-laying and b nestling stages

Mean±SE time (in seconds) female yellow warblers sat in their nests in response to seet and chip playbacks in the sympatric population during egg-laying and nestling stages
Nestling stage
Cowbirds present little threat to nestlings at our study site (see McLaren and Sealy 2000), so we did not expect females to respond to seet playbacks when caring for young. Almost all females returned to their nest areas during both playbacks, giving chip calls and moving within 2 m of the nest and speaker. There was no significant difference between playbacks in the number of seet calls elicited, as few females seet called during the seet playback, and none did so during the chip playback (Fig. 1b; Z=1.604, n=23, P>0.1). However, the duration of nest-protection behaviour was significantly greater during seet playbacks than chip playbacks (Fig. 2; Z=2.201, n=23, P=0.02), during which no females returned to their nests. The mean rate of chip calling during chip playbacks was more than 3 times greater than during seet playbacks, but the difference was not significant (Fig. 1b; Z=1.556, n=23, P>0.12).
Comparison of nesting stages
Females gave seet calls and nest-protection behaviour at a higher rate (Fig. 1; Wilcoxon two-sample test, Z=3.201, P=0.001) and for longer duration (Fig. 2; Z=2.903, P=0.004), respectively, when they were laying than when caring for nestlings. There were no differences between nesting stages in the intensity of females’ responses to chip playbacks (Fig. 1; Z>1.371, P>0.2).
Test of response urgency in the sympatric population
If seet calls reflect the immediacy of responses required to prevent brood parasitism, then female yellow warblers should give seet calls primarily when the cowbird model is close to the nest, simulating the highest and most immediate risk. The degree of risk appeared to have little effect on alarm calling (Fig. 3), as females gave seet calls (Friedman test, χ 2=3.947, n=30, P>0.1) and chip calls (χ 2=1.636, n=30, P>0.4) at similar rates to the model at each distance. However, the distances at which the models were presented appeared to be perceived as a change in threat as the duration of nest-protection behaviour was significantly greater when the model was beside the nest than when farther away (Fig. 4; χ 2=11.439, n=30, P=0.003).

Number (mean±SE) of seet and chip calls given by female yellow warblers in response to models of cowbirds simulating different risks of parasitism

Mean ± SE time (in seconds) female yellow warblers sat in their nests in response to models of cowbirds simulating different risks of parasitism
Responses of yellow warblers allopatric with cowbirds
Model presentation
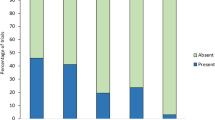
We explored whether seet calls have a different referent in a population of yellow warblers allopatric with cowbirds by presenting models of a cowbird, an avian nest predator, and a non-threatening control. Overall, females responded similarly to all models, typically chipping and changing perches within 2 m of the models. There were no differences among models in the number of chip calls (Fig. 5; χ 2=1.64, n=17, P>0.4) or seet calls (χ 2=2.00, n=17, P>0.3) elicited. Only one of 17 females produced seet calls in response to each model (two males also gave seet calls during presentations of the cowbird model; see Gill 1995). In conjunction with seet calling, this female also performed nest-protection behaviour during presentation of the cowbird model. Several other females sat in their nests during playbacks, but they gave neither seet calls nor rushed back to the nest, behaviours that characterise the nest-protection behaviour of females in the sympatric population. The time females spent in their nests did not differ significantly among models (Fig. 6; Friedman’s test, χ 2=3.739, n=17, P>0.15).

Number (mean±SE) of seet and chip calls given by female yellow warblers to models of cowbird, gray jay and fox sparrow presented in the allopatric population

Mean±SE time (in seconds) female yellow warblers sat in their nests to models of cowbird, gray jay and fox sparrow presented in the allopatric population
Playback of cowbird-specific calls
Responses of female yellow warblers in the allopatric population were similar between seet and chip playbacks. Most females returned to within 5 m of their nests and changed perches around them and the speaker during playbacks. No females gave seet calls in response to either playback, although one female sat in her nest during the seet playback. Females gave more chip calls in response to chip playbacks (mean±SE=11.1±4.04 chips per min) than seet playbacks (1.5±0.58 chips per min; Wilcoxon signed-ranks test, t s=1.96, n=15, P=0.05).
Comparison of yellow warblers sympatric and allopatric with cowbirds
Clear differences existed in the responses to seet playbacks of females in populations allopatric and sympatric with cowbirds. During seet playbacks, females sympatric with cowbirds gave significantly more seet calls (Wilcoxon two-sample test, Z=3.545, n=41, P=0.0004) and sat in their nests significantly longer (Z=3.154, n=41, P=0.002) than females in the allopatric population. There was no significant difference between populations in the number of chip calls given (Z=0.455, n=41, P=0.65). During chip playbacks, females in the allopatric population gave, though not significantly, chip calls at higher rates than females in the sympatric population (Z=1.782, P=0.07), and there were also no significant differences between populations for seet calls or nest-protection behaviour (Z=1.109, n=41, P=0.27 for both comparisons).
Discussion
Most female yellow warblers sympatric with brown-headed cowbirds responded to seet playbacks with nest-protection behaviour, a response typically elicited by the presence of cowbirds in the nesting area. This result indicated that individuals receive and extract specific information about the presence of a cowbird from seet calls in the absence of both cowbirds and signallers, satisfying the requirement of perception specificity. Together with acoustical distinctiveness of seet calls and their preferential production in response to cowbirds and their vocalisations observed in previous studies (Hobson and Sealy 1989a; Gill 1995; Gill and Sealy 1996; Gill et al. 1997), we conclude that yellow warblers possess a functionally referential alarm call used during nest defence. To our knowledge, our study is the first to describe functional reference in alarm calls given during nest defence in general and specifically against a brood parasite, and is one of only a handful of studies to date that has investigated the meaning of alarm calls in birds (Ficken and Witkin 1977; Gyger et al. 1987; Ficken 1990; Regosin 2002; Seddon et al. 2002; see also Naguib et al. 1999).
The degree of production specificity shown by functionally referential alarm calls varies among species, with some species showing narrow classification of predators, to others where predators are broadly grouped (Evans 1997). For example, eagle alarm calls of vervet monkeys are given primarily to martial eagles (Polemaetus bellicosus: Seyfarth and Cheney 1980), whereas aerial alarm calls of domestic chickens are given to an array of flying objects (Gyger et al. 1987). Our findings suggest that the specificity of yellow warbler seet calls is quite high as they are given principally to the narrow class of brood-parasites and not the more general class of nest threats, including avian nest predators. Specificity in seet calls may occur because cowbirds threaten the nest in a way that is distinct from nest predators, including birds (see below). Yellow warblers are parasitised by shiny cowbirds (M. bonariensis) in other parts of their range (Wiley 1985; Post et al. 1990) and it would be interesting to determine whether these populations produce seet calls or other cowbird-specific vocalisations and, if so, whether functional referentiality exists.
Context plays an important role in modulating the responses of yellow warblers to cowbirds and seet playbacks as females adjusted their responses depending on nesting stage. Yellow warblers in the sympatric population responded most strongly to cowbird models (Hobson and Sealy 1989a; Gill and Sealy 1996) and seet playbacks (this study) during laying when nests are vulnerable to cowbird parasitism. When yellow warblers have nestlings, cowbirds pose little threat to their reproductive success in our study population, as they have never been observed to depredate nestlings (McLaren and Sealy 2000). Nevertheless, 6 of 23 females caring for nestlings responded to seet playbacks with nest-protection behaviour, which suggests that while seet calls may still be functionally referential at later nesting stages, the motivation of most females to respond to such calls may be low. The use of contextual information, in this case an individual’s nesting stage, results in a more adaptive response matched to the threat posed by cowbirds.
Response urgency did not affect alarm calling by females, as the rate of seet calling did not differ among cowbird models simulating different risks of parasitism. However, females apparently perceived the models as different dangers because they spent more time in nest-protection behaviour when the model was closest to the nest. There is additional evidence that urgency of response is not reflected in seet calls, as these calls were rarely elicited by avian or mammalian nest predators (Hobson et al. 1988; Gill and Sealy 1996). If seet calls were given in high-urgency situations, then warblers would be expected to use them when confronting nest predators, which lower reproductive success of yellow warblers in our study population to a greater extent than cowbirds (Goossen and Sealy 1982). Although seet calls do not reflect response urgency in the broad sense, urgency may be revealed with a finer scale of analysis. Call structure (Manser 2001), rate and intensity (Blumstein 1995; Warkentin et al. 2001) may vary with risk and communicate response urgency to receivers, possibilities that warrant further investigation in yellow warblers.
What is the adaptive significance of functional reference in the seet calls of yellow warblers? In group-living animals, callers may benefit from warning group members and also may derive selfish benefits from decreased predation risk (e.g. Manser et al. 2001). In socially monogamous yellow warblers, callers may benefit from informing mates about cowbirds in their nesting area, if mates are more likely to return to aid in nest defence. Females are usually the first line of defence against cowbirds because they respond more rapidly and more often than males to models and playbacks, only females perform nest-protection behaviour (Gill and Sealy 1996), and they are more likely to be on the nest at dawn when cowbirds come to lay (Neudorf and Sealy 1994; Sealy et al. 2000). Thus, whereas females are more responsive to seet calling and would be more likely to be recruited for nest defence than their mates, males rarely respond first. Female warblers may also manage (Owings and Morton 1998) the behaviour of cowbirds by giving seet calls, as they are given repetitively until the female enters her nest, following which she becomes silent regardless of whether her mate has responded. This suggests that the cowbird may be an additional receiver of seet calls. Similarly, the long-distance calls of Diana monkeys simultaneously function to warn conspecifics of predators and as perception-advertisement calls to predators that rely on surprise attacks (Zuberbühler et al. 1997). Playback experiments are needed to examine the responses of cowbirds to seet calls and to determine whether they play a role in deterring cowbirds from parasitising nests.
The evolution of referential alarm calls
In contrast to the sympatric population, yellow warblers not exposed to brood parasitism by cowbirds never produced seet calls in response to playbacks, and only rarely to models of a cowbird and an avian nest predator. One female performed nest-protection behaviour during the seet playback. Together with Briskie et al. (1992), our results have documented the nearly complete absence of seet calls (2/30 females gave seet calls) and nest-protection behaviour (1/30 females) to cowbird models in the allopatric population. This contrasts dramatically with our observations in the sympatric population, in which 91% and 74% of females (n=35) gave seet calls and nest-protection behaviour, respectively, to cowbird models placed near their nests (Gill 1995, unpublished data). The different responses to cowbird models and seet calls in allopatric and sympatric populations provide strong evidence that functional referentiality in yellow warbler seet calls evolved as a direct consequence of brood parasitism by cowbirds and not due to other selection pressures, such as avian nest predation.
Referential alarm calls may evolve when a species is exposed to predators that exhibit different hunting styles that require disparate escape strategies (Macedonia and Evans 1993). Yellow warblers may respond uniquely to cowbirds versus nest predators because successful defence against each requires vastly different responses. Yellow warblers perform behaviours that appear to prevent cowbirds from entering and parasitising nests as well as removing host eggs (Sealy et al. 1998; Tewksbury et al. 2002), whereas nest predators elicit distraction displays that may lure them away from nests (Gill and Sealy 1996). It is unlikely that cowbirds could be lured away from nests by what appears to be easy but relatively large prey, whereas returning to sit in their nests to defend against predators would probably render females both helpless to prevent predation and very likely to be injured and possibly killed themselves. Thus, yellow warblers have evolved defensive strategies well matched to the different threats posed by brood parasitism and nest predation.
Yellow warblers are remarkable because, to our knowledge, they are the only passerine species that displays unique behavioural responses to brood parasites. Studies on nest defence by hosts of cowbirds and cuckoos have revealed that many species react more strongly to brood parasites during the laying cycle of the host than at other nesting stages (e.g. Neudorf and Sealy 1992), that host responses to brood parasites and nest predators or non-threatening species are quantitatively, but not qualitatively, different (e.g. Duckworth 1991; Neudorf and Sealy 1992; Hobson and Villard 1998; Prather et al. 1999), and that defensive responses are less developed or absent in populations allopatric or recently sympatric with brood parasites (e.g. Lindholm and Thomas 2000; Røskaft et al. 2002; but see, e.g., Prather et al. 1999). These studies show that many species recognise the unique threat posed by brood parasites, but none, other than the studies on yellow warblers, have shown that hosts respond with unique behaviours to brood parasites. Why yellow warblers, and not other passerines, have evolved both a unique functionally referential call as well as unique defensive behaviour to cowbirds is unknown, because presumably individuals of many species would benefit from similarly specific responses. The interactions of yellow warblers with cowbirds have been more thoroughly studied than almost any passerine, and we may yet discover cowbird-specific responses in other species. However, the apparent absence of unique responses to cowbirds in other species is likely more than just sampling bias because, in addition to seet calls and nest-protection behaviour, only yellow warblers regularly bury cowbird eggs (Sealy 1995). This behaviour also evolved as a direct consequence of cowbird parasitism, as it is not a by-product of conspecific brood parasitism and warblers in populations allopatric with or recently exposed to cowbirds do not express it (reviewed in Sealy 1995). Thus, yellow warblers employ a suite of anti-parasite strategies unparalleled by other passerines studied to date.
Recently, Rothstein (2001) provided persuasive evidence that most potential hosts of cowbirds and cuckoos retain the ability to eject parasitic eggs in the absence of current brood parasitism. This “relic” behaviour may be expressed in the absence of brood parasitism because there are few or no costs in retaining egg ejection (Rothstein 2001). Such retention contrasts sharply with observations of nest defence in yellow warblers (Briskie et al. 1992; this study) and other host species (Lindholm and Thomas 2000; Roskaft et al. 2002; but see Prather et al. 1999), as well as observations of some predator-prey systems (e.g. Foster 1994; Berger et al. 2003; but see Coss 1993; Byers 1997), in which defensive behaviours apparently have been lost. Many of these studies point to the potentially high costs of exhibiting anti-parasite or anti-predator behaviours in the absence of these risks. We have argued that yellow warbler responses to cowbirds and predators differ because if females reacted to nest predators as they do to cowbirds (by sitting in their nests), they would very likely risk serious injury and possibly death. Thus, the cost to exhibiting these unique behavioural responses to nest predators may have favoured their almost complete loss in the allopatric population. At present, however, we are unable to infer whether the expression of seet calls and nest-protection behaviour by a small number of females in the allopatric population is due to the retention of these behaviours or to gene flow from parasitised populations (Briskie et al. 1992). Further experimentation on additional yellow warbler populations allopatric and sympatric with cowbirds in conjunction with documentation of their historical relationships is needed to understand more fully the evolution of these remarkable behaviours.
References
Berger J, Monfort SL, Roffe T, Stacey PB, Testa JW (2003) Through the eyes of prey: how the extinction and conservation of North America’s large carnivores alter prey systems and biodiversity. In: Festa-Bianchet M, Apollonio M (eds) Animal behavior and wildlife conservation. Island Press, Washington, DC, pp 133–155
Blumstein DT (1995) Golden-marmot alarm calls: I. the production of situationally specific vocalizations. Ethology 100:113–125
Blumstein DT, Armitage KB (1997) Alarm calling in yellow-bellied marmots: I. The meaning of situationally variable alarm calls. Anim Behav 53:143-171
Briskie JV, Sealy SG, Hobson KA (1992) Behavioral defenses against avian brood parasitism in sympatric and allopatric host populations. Evolution 46:334–340
Byers JA (1998) American pronghorns: social adaptations and the ghosts of predators past. University of Chicago Press, Chicago
Coss RG (1993) Evolutionary persistence of ground squirrel antisnake behavior: reflections on Burton’s commentary. Ecol Psychol 5:171–194
Duckworth JW (1991) Responses of breeding reed warblers Acrocephalus scirpaceus to mounts of sparrowhawk Accipter nisus, cuckoo Cuculus canorus and jay Garrulus glandarius. Ibis 133:68–74
Evans CS (1997) Referential signals. In: Owings DH, Beecher MD, Thompson NS (eds) Perspectives in ethology. Plenum, New York, pp 99–143
Evans CS, Evans L, Marler P (1993) On the meaning of alarm calls: functional reference in an avian vocal system. Anim Behav 46:23–38
Falls JB (1982) Individual recognition by sounds in birds. In: Kroodsma DE, Miller EH (eds) Acoustic communication in birds. Vol. 2 Academic Press, New York, pp 237–278
Fichtel C, Kappeler PM (2002) Anti-predator behavior of group-living Malagasy primates: mixed evidence for a referential alarm call system. Behav Ecol Sociobiol 51:262–275
Ficken MS (1990) Acoustic characteristics of alarm calls associated with predation risk in chickadees. Anim Behav 39:400–401
Ficken MS, Ficken RW (1965) Comparative ethology of the chestnut-sided warbler, yellow warbler, and American redstart. Wilson Bull 77:363–375
Ficken MS, Witkin SR (1977) Responses of black-capped chickadee flocks to predators. Auk 94:156–157
Foster SA (1994) Inference of evolutionary pattern: diversionary displays of three-spined sticklebacks. Behav Ecol 5:114–121
Gill SA (1995) Information transfer, function, and evolution of yellow warbler alarm calls. MSc thesis, University of Manitoba, Canada
Gill SA, Sealy SG (1996) Nest defence by yellow warblers: recognition of a brood parasite and an avian nest predator. Behaviour 133:263–282
Gill SA, Sealy SG (2003) Tests of two functions of alarm calls given by yellow warblers during nest defence. Can J Zool 81:1685–1690
Gill SA, Neudorf DL, Sealy SG (1997) Host responses to cowbirds near the nest: cues for recognition. Anim Behav 53:1287–1293
Godfrey WE (1986) The birds of Canada. National Museum of Natural Sciences, Ottawa
Goossen JP, Sealy SG (1982) Production of young in a dense nesting population of yellow warblers, Dendroica petechia, in Manitoba. Can Field-Nat 96:189–199
Gyger M, Marler P, Pickert R (1987) Semantics of an avian alarm call system: the male domestic fowl, Gallus domesticus. Behaviour 102:15–40
Hobson KA, Sealy SG (1989a) Responses of yellow warblers to the threat of cowbird parasitism. Anim Behav 38:510–519
Hobson KA, Sealy SG (1989b) Mate guarding in the yellow warbler Dendroica petechia. Ornis Scand 20:241–249
Hobson KA, Villard M-A (1998) Forest fragmentation affects the behavioral response of American redstarts to the threat of cowbird parasitism. Condor 100:389–394
Hobson KA, Bouchart ML, Sealy SG (1988) Responses of naive yellow warblers to a novel nest predator. Anim Behav 36:1823–1830
Le Roux A, Jackson TP, Cherry MI (2001) Does Brants’ whistling rat (Parotomys brantsii) use an urgency-based alarm call system in reaction to aerial and terrestrial predators? Behaviour 138:757–773
Leger DW, Owings DH, Boal LM (1979) Contextual information and differenitial responses to alarm whistles in California ground squirrels. Z Tierpsychol 49:142–155
Lindholm AK, Thomas RJ (2000) Differences between populations of reed warblers in defences against brood parasitism. Behaviour 137:25–42
Lorenzana JC, Sealy SG (1999) A meta-analysis of the impact of parasitism by the brown-headed cowbird on its hosts. Stud Avian Biol 18:241–253
McLaren CM, Sealy SG (2000) Are nest predation and brood parasitism correlated in yellow warblers? A test of the cowbird predation hypothesis. Auk 117:1056–1060
Macedonia JM (1990) What is communicated in the antipredator calls of lemurs: evidence from playback experiments with ringtailed and ruffed lemurs. Ethology 86:177–190
Macedonia JM, Evans CS (1993) Variation among mammalian alarm call systems and the problem of meaning in animal signals. Ethology 93:177–197
Manser MB (2001) The acoustic structure of suricates’ alarm calls varies with predator type and the level of response urgency. Proc R Soc Lond B 268:2315–2321
Manser MB, Bell MB, Fletcher LB (2001) The information that receivers extract from alarm calls in suricates. Proc R Soc Lond B 268:2185–2491
Marler P, Evans CS, Hauser MD (1992) Animals signals: motivational, referential, or both? In: Papousek H, Jürgens U, Papousek M (eds) Nonverbal vocal communication: comparative and developmental approaches. Cambridge University Press, Cambridge, pp 66–86
Naguib M, Mundry R, Ostreiher R, Hultsch H, Schrader L, Todt D (1999) Cooperatively breeding Arabian babblers call differently when mobbing in different predator-induced situations. Behav Ecol 10:636–640
Neudorf DL, Sealy SG (1992) Reactions of four passerine species to threats of predation and cowbird parasitism: enemy recognition or generalized responses? Behaviour 123:84–105
Neudorf DL, Sealy SG (1994) Sunrise nest attentiveness in cowbird hosts. Condor 96:162–169
Ortega CP (1998) Cowbirds and other brood parasites. University of Arizona Press, Tucson
Owings DH, Morton ES (1998) Animal vocal communication: a new approach. Cambridge University Press, Cambridge
Pereira ME, Macedonia JM (1991) Ringtailed lemur anti-predator calls denote predator class, not response urgency. Anim Behav 41:543–544
Post W, Nakamura TK, Wiley JW (1990) Patterns of shiny cowbird parasitism in St. Lucia and southwestern Puerto Rico. Condor 92:461–469
Prather JW, Ortega CP, Cruz A (1999) Aggressive responses of red-winged blackbirds (Agelaius phoeniceus) toward brown-headed cowbirds (Molothrus ater) in areas of recent and long-term sympatry. Bird Behav 13:1–7
Regosin, JV (2002) Alarm calling and predator discrimination in the Arabian babbler (Turdoides squamiceps) Bird Behav 15:11–19
Reid ML, Sealy SG (1986) Behavior of a polygynous yearling yellow warbler. Wilson Bull 98:315–317
Røskaft E, Moksnes A, Stokke BG, Bicík V, Moskát C (2002) Aggression to dummy cuckoos by potential European cuckoo hosts. Behaviour 139:613–628
Rothstein SI (2001) Relic behaviours, coevolution and the retention versus loss of host defences after episodes of avian brood parasitism. Anim Behav 61:95–107
Sealy SG (1994) Observed acts of egg destruction, egg removal, and predation on nests of passerine birds at Delta Marsh, Manitoba. Can Field Nat 108:41–51
Sealy SG (1995) Burial of cowbird eggs by parasitized yellow warblers: an empirical and experimental study. Anim Behav 49:877–889
Sealy SG, Neudorf DL, Hill DP (1995) Rapid laying by brown-headed cowbirds Molothrus ater and other parasitic birds. Ibis 137:76–84
Sealy SG, Neudorf DL, Hobson KA, Gill SA (1998) Nest defence by potential hosts of the brown-headed cowbird : methodological approaches, benefits of defence, and coevolution. In: Rothstein SI, Robinson SK (eds) Parasitic birds and their hosts. Oxford University Press, New York, pp 194–211
Sealy SG, McMaster DG, Gill SA, Neudorf DL (2000) Yellow warbler nest attentiveness before sunrise: antiparasite strategy or onset of incubation? In: Smith JNM, Cook TL, Rothstein SI, Robinson, SK, Sealy SG (eds) Ecology and management of cowbirds and their hosts. University of Texas Press, Austin, pp 169–177
Seddon N, Tobias JA, Alvarez A (2002) Vocal communication in the pale-winged trumpeter (Psophia leucoptera): repertoire, context and functional reference. Behaviour 139:1331–1359
Seyfarth R, Cheney D (1990) The assessment by vervet monkeys of their own and another species’ alarm calls. Anim Behav 40:754–764
Seyfarth RM, Cheney DL, Marler P (1980) Vervet monkey alarm calls: semantic communication in a free-ranging primate. Anim Behav 28:1070–1094
Studd, M, Robertson RJ (1985) Sexual selection and variation in reproductive strategy in male yellow warblers (Dendroica petechia). Behav Ecol Sociobiol 17:101–109
Tewksbury JJ, Martin TE, Hejl SJ, Kuehn MJ, Jenkins JW (2002) Parental care of a cowbird host: caught between the costs of egg-removal and nest predation. Proc R Soc Lond B 269:423–429
Warkentin KJ, Keeley ATH, Hare JF (2001) Repetitive calls of juvenile Richardson’s ground squirrels (Spermophilus richardsonii) communicate response urgency. Can J Zool 79:569–573
Wiley JW (1985) Shiny cowbird parasitism in two avian communities in Puerto Rico. Condor 87:165–176
Yezerinac SM, Weatherhead PJ (1997) Extra-pair mating, male plumage coloration and sexual selection in yellow warblers (Dendroica petechia). Proc R Soc Lond B 264:527–532
Zuberbühler K (2000) Referential labelling in Diana monkeys. Anim Behav 59:917–927
Zuberbühler K, Noë R, Seyfarth RM (1997) Diana monkey long-distance calls: messages for conspecifics and predators. Anim Behav 53:589–604
Acknowledgements
We thank the Portage Country Club for permission to work on their property, and the staff of the Delta Marsh Field Station (University of Manitoba) and Churchill Northern Studies Centre for logistical support during the fieldwork. K. Caldwell and D.G. McMaster were instrumental in the field. Constructive comments were made by J.F. Hare, W.A. Searcy, M.J. Vonhof and two anonymous reviewers. This study was supported by scholarships and grants from NSERC, Faculty of Science (University of Manitoba), Frank M. Chapman Memorial Fund (American Museum of Natural History), and Northern Studies Training Program to S.A.G., and by a NSERC grant (A9556) to S.G.S.. Our research was conducted with the approval of the Canadian Council on Animal Care.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by W.A. Searcy
Rights and permissions
About this article
Cite this article
Gill, S.A., Sealy, S.G. Functional reference in an alarm signal given during nest defence: seet calls of yellow warblers denote brood-parasitic brown-headed cowbirds. Behav Ecol Sociobiol 56, 71–80 (2004). https://doi.org/10.1007/s00265-003-0736-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-003-0736-7