
Abstract
Worker-reproduction is rare in queenright honey-bee colonies. When workers do lay eggs, their eggs are normally eaten by other workers presumably because they lack the queen's egg-marking signal. Workers use the absence of this queen signal to enforce the queen's reproductive monopoly by policing any worker-laid eggs. In contrast, in anarchistic colonies, the majority of the males arise from worker-laid eggs. Anarchistic worker-laid eggs escape policing because workers perceive anarchistic eggs as queen-laid. However, in this study, we show that eggs laid by queenless anarchistic workers do not escape policing and have very similar removal rates to worker-laid eggs from queenless wild-type (i.e. non-anarchistic) colonies. This suggests that, under queenless conditions, eggs laid by anarchistic workers lose their chemical protection and are therefore no longer perceived as queen-laid. Hence, the egg-marking signal seems to be only applied to eggs when queen and brood are present. This suggests that in the absence of queen and brood, the biosynthetic pathway that produces the egg-marking signal is switched off.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Social insects have long been seen as a classic example of a harmonious society. However, since Hamilton's (1964) seminal work, we know that insect societies are the scene of numerous potential and actual conflicts. Conflicts occur because insect societies are non-clonal and this leads to potential conflict among individuals or groups of individuals over colony reproduction (Ratnieks and Reeve 1992). One such conflict is that over production of male-producing eggs.
In the majority of social-insect species, worker reproduction is facultative; that is, workers have functional ovaries but often refrain from reproducing in the presence of the queen (Bourke 1988). The extent to which worker reproduction occurs in queenright colonies is determined by the interplay of selective forces working both at the level of the individual worker and the workers collectively. In polyandrous species like honey bees, selection acts at the level of the collective workers to decrease worker reproduction, thereby minimising conflict among workers over which individual(s) will produce males (Ratnieks 1988). At the same time, selection acts at the individual level to increase worker reproduction, as any worker that can harness the brood-production system of its colony for personal reproduction is at an enormous fitness advantage over its non-reproductive peers (Oldroyd and Osborne 1999; Barron et al. 2001).
In honey bees, the outcome of these divergent selective forces is strongly biased in favour of functional worker sterility. In the vast majority of colonies, 99.9% of adult male offspring are derived from queen-laid eggs (Ratnieks 1993; Visscher 1996). Functional worker sterility is the result of two different mechanisms. First, workers normally refrain from reproduction because they respond to pheromonal signals emanating mainly from the brood (Trouiller et al. 1991; Arnold et al. 1994; Mohammedi et al. 1998; Oldroyd et al. 2001b). Second, any eggs that are laid by workers are removed by other workers in a process known as worker policing (Ratnieks 1988; Ratnieks and Visscher 1989).
The selective removal of worker-laid eggs is presumably made possible because the queen marks her eggs, thereby enabling the workers to discriminate between queen-laid (those with the mark) and worker-laid (those lacking the mark) eggs. Although the nature of the egg-marking signal is yet unknown (Katzav-Gozansky et al. 2001, 2002; Oldroyd et al. 2002), a chemical difference between queen-laid and worker-laid eggs seems the most likely means by which workers discriminate between both type of eggs (Katzav-Gozansky et al. 2001). Both the receivers (the workers) and the sender (the queen) benefit from the presence of such a queen-produced egg-marking signal. Workers benefit from the signal because their indirect fitness is increased by the removal of worker-laid eggs to which they are less related than male-producing eggs laid by the queen. The queen benefits because the signal allows her to monopolise the colony's male production (Ratnieks 1988).
Despite the colony-level benefits of workers refraining from reproducing, some Apis mellifera workers do lay eggs in queenright colonies, but nearly all of these eggs are quickly eaten by other workers (Ratnieks 1993; Visscher 1996; Martin et al. 2002). In the Asian honey bee, A. cerana, however, worker-ovary activation is relatively common (1–5%) although successful reproduction is rare (Oldroyd et al. 2001a). The fact that workers do try to reproduce despite the presence of the queen and brood shows that actual conflict over male production still occurs in honey-bee colonies. At the same time, the rarity of worker-produced males shows that worker policing is an effective mechanism that prevents workers from reproducing.
In contrast to the normal condition of functional worker sterility, in a few very rare "anarchistic" colonies, the majority of adult males arise not from the queen, but from eggs laid by the workers of a limited number of patrilines (Oldroyd et al. 1994; Montague and Oldroyd 1998; Châline et al. 2002). The production of worker-laid males in anarchistic colonies arises from two key characteristics of their workers. First, anarchistic workers have the ability to activate their ovaries despite the presence of brood (Barron and Oldroyd 2001). Second, anarchistic workers lay eggs that evade policing (Oldroyd and Ratnieks 2000). These two traits have been fixed in the anarchistic line maintained at Sydney University (Oldroyd and Osborne 1999), such that up to 41% of 14-day-old workers have active ovaries (Oldroyd et al. 2001b) and all or nearly all of the males in these colonies arise from worker-laid eggs. Hereafter we use the term "anarchistic" to describe workers of this line.
Anarchy is rare in naturally occurring honey-bee colonies and yet the fitness pay-off for an individual anarchistic worker is substantial because she produces her own sons. Similarly, the males arising from anarchistic workers are at an advantage over queen-derived males because they not only father female offspring, but are likely to produce male offspring via their reproductive worker-daughters (Oldroyd et al. 1994; Montague and Oldroyd 1998). Thus the selective forces that keep worker-reproduction so rare must be very strong. Surprisingly, however, significant fitness costs related to behaviour of individual anarchistic workers have not been found in that anarchistic workers perform worker tasks normally (Oldroyd et al. 1999) albeit at a slightly slower rate (Dampney et al. 2003). Notwithstanding the above, high rates of worker reproduction, as occur in the selected anarchistic line, are unsustainable at the colony level (Barron et al. 2001).
Here we investigate the fate of eggs laid by anarchistic workers from queen- and broodless (hereafter called "queenless") and queenright anarchistic colonies. We transferred anarchistic worker-laid eggs from queenright and queenless anarchistic colonies into queenright wild-type discriminator colonies. The results show that eggs laid by queenless anarchistic workers are removed almost at a similar rate as eggs laid by wild-type worker-laid eggs, whereas eggs laid by queenright anarchistic workers are removed at a much slower rate. This suggests that when queenless, eggs laid by anarchistic workers do not carry the signal that normally prevents them from being policed.
Methods
Sources of eggs
Seven days before assays commenced, we divided two anarchistic colonies into two halves, one containing the original queen and young brood and the other containing no queen, and no young brood from which the workers could rear a replacement queen. Each colony was placed in a two-box hive, each box containing four combs, and the top and bottom boxes separated by a queen excluder that prevents the queen but not the workers from entering the top box. Egg-source combs from the queenright anarchist colonies were only taken from the box above the queen excluder, so that all eggs were known to be worker-laid (Montague and Oldroyd 1998).
All assays were performed on unfertilised eggs less than 24 h old. To obtain freshly laid eggs, we introduced combs containing drone-sized cells into the egg-source colonies 24 h before the assay. Queen-laid eggs were obtained from three randomly selected wild-type colonies. These wild-type colonies were unrelated to the discriminator colonies (see below). Wild-type worker-laid eggs were obtained from two randomly selected queenless laying-worker colonies. These colonies were made queen- and broodless 17 days before assays commenced.
Bioassays
Experiments were performed at Pearl Beach, New South Wales, Australia, during January and February 2002. Foraging conditions were poor at this time due to the drought and bush fires of December 2001, so all colonies were provided with supplementary sugar syrup and ground bee-collected pollen daily throughout the experiment.
To assay the removal rates of eggs from various sources, we used three wild-type discriminator colonies. Each discriminator colony was of standard commercial type, comprising two boxes separated by a queen excluder that confined the queen to the lower box. Each discriminator colony contained 17 combs covered by bees, 8 of which contained brood, and a sugar syrup feeder.
To perform the assay, we transferred 20 eggs from each of the 4 different sources (wild-type queen-laid, wild-type worker-laid, queenright and queenless anarchist worker-laid) into drone cells of clean test combs using standard policing assay techniques (Ratnieks and Visscher 1989; Oldroyd and Ratnieks 2000). Eggs were removed from their original cell either by using modified forceps or a blunt needle. Eggs of each egg-source were then carefully placed in random rows in the test comb such that there were 4 rows of 20 eggs per test comb. The test combs had been left overnight in their respective discriminator colonies. After egg transfers, we sandwiched the test comb between two brood combs that we had previously placed in the centre of the upper chamber of each discriminator colony. The number of eggs remaining was determined 1, 2, 4 and 6 h after the introduction of the test combs. Assays were performed on 7 separate days, using a total of 420 eggs of each egg-source; these were 9, 11, 12, 13, 19, 22 and 23 days after the queenless anarchistic splits were made.
Statistical analysis
Where possible, we varied the egg-source colonies used on each day, but this was irregular. For this reason, we do not analyse the differences between source colonies within each egg-source.
Data were analysed using a Cox regression survival analysis (Collett 1994) as implemented in SPSS. We compared the survival of eggs laid by queens, queenless and queenright anarchists, and wild-type workers using exact failures (i.e. a transferred egg was removed at hours 1, 2, 4 or 6) and right censoring (for those eggs still remaining at hour 6). The null hypothesis in a Cox regression model assumes that the hazard rate (i.e. probability of failure) at any given time for an individual (here an egg) in one group is proportional to the hazard at that time for a similar individual (egg) in the other group(s). The model we constructed included egg-source, day and discriminator as variables. We used this model to calculate the likelihood ratio for the survival rate of the different egg-sources after allowing for the effects of day and discriminator. We present the data graphically as the untransformed mean proportion of eggs remaining at each time period rather than the hazard function (Collett 1994), which is less informative.
Results
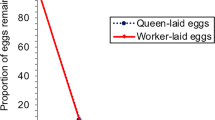
In all three discriminator colonies, eggs laid by queenright and queenless anarchistic workers were treated differently: eggs laid by queenless anarchistic workers were removed quickly whereas eggs laid by queenright anarchistic workers were removed at a much slower rate (Fig. 1).

Removal rates of eggs laid by wild-type workers and queens, and queenright and queenless anarchistic workers when introduced into three (1–3) wild-type queenright discriminator colonies. The fourth panel (All) shows the mean of all the discriminators. The bars represent the standard errors of the means (seven trials per colony). In some cases the error bars are very small and so are obscured by the data point
A Cox regression analysis (Table 1) shows that there were statistically significant differences in removal rates for the four different egg-sources ("Source of eggs": P<0.001). The day of the assay did not have a significant effect on removal rates ("Day": P=0.4) nor did the discriminator colony used ("Discriminator": P=0.2). Table 1 also shows the results of pair-wise comparisons of the removal rates using pooled data of all 7 days and all three discriminators (Fig. 1 "All" panel). The pair-wise comparisons show that eggs laid by queenless anarchists are removed at the same rate as eggs laid by wild-type workers (P=0.06) whereas eggs laid by queenright anarchists are removed much more slowly than wild-type worker eggs (P<0.001). Queenright and queenless anarchistic eggs are also treated differently (P<0.001).
Discussion
The removal rates of eggs laid by queenless anarchistic workers appear to be more similar to those of wild-type worker-laid eggs, whereas removal rates of eggs laid by queenright anarchistic workers are intermediate to those of queen- and worker-laid eggs. Hence, when anarchistic workers are queen- and broodless, their eggs do not seem to carry the egg-marking signal and, as a result, are policed. Our results also confirm the findings of Oldroyd and Ratnieks (2000), who showed that eggs laid by queenright anarchistic workers have survival rates intermediate between queen-laid and wild-type worker-laid eggs. Because of the poor foraging conditions caused by the extreme drought, a relatively large number of queen-laid eggs were removed in the bioassays. However, because we compare relative removal rates, this does not affect our conclusions.
How can we explain the decline in the acceptability of eggs laid by queenless anarchistic workers? We suggest that three hypotheses are plausible.
Under the first hypothesis, it is suggested that preselection of eggs occurs in anarchistic colonies, and that this preselection is reduced or eliminated when anarchistic colonies are queenless. It is proposed that eggs of a range of acceptability are produced by queenright anarchistic workers. Those eggs with low acceptability (i.e. those not properly marked) are removed by policing so that only those eggs that do carry the egg-marking pheromone remain for transfer by experimenters. Under this scenario, transferred eggs are likely to have a higher acceptability than the majority of eggs laid in anarchistic colonies and will have low removal rates in policing assays. If rates of policing are lowered in queenless anarchistic colonies, then preselection would be reduced thereby decreasing the average acceptability of eggs in policing assays.
We suggest that differential preselection rates (arising from lower policing rates in queenless colonies) between queenright and queenless anarchistic colonies are an unlikely cause of our results for three reasons. First, rates of policing are known to be reduced in queenright anarchistic colonies (Oldroyd and Ratnieks 2000), so preselection, if it exists, is likely to be of little consequence. Second, Miller and Ratnieks (2001) have shown that egg-removal rates (of eggs laid by both queens and workers) increase rather than decrease in queenless colonies. An indiscriminate increase in egg-eating in our queenless anarchistic colonies cannot explain our results because it would not lead to a proportional increase of eggs of low acceptability (i.e. those eggs that would be policed by wild-type workers). Third, the survival rates of anarchist-laid eggs in situ appear to be high, with less than 7.3% of eggs being removed within 2 h of oviposition (Ratnieks et al. 2003), again suggesting that preselection is not important.
Under the second hypothesis, individual egg-laying rates differ for queenright and queenless workers. It could be that when queenright, anarchistic workers only lay a few eggs per day, which enables them to produce sufficient amounts of the egg-marking signal to prevent their eggs from being policed. When queenless, however, individual egg-laying rates may go up to such an extent that workers may not be able to produce sufficient egg-marking signal to protect the larger number of eggs laid. There is some evidence for a difference in individual egg-laying rates under queenright and queenless conditions in wild-type honey-bee colonies. Ratnieks (1993), studying worker reproduction in queenright wild-type colonies, reported that the two workers that had activated ovaries only had a single fully developed egg in an ovary. This is in stark contrast to the 19–32 eggs laid per day per worker under queenless conditions (see references in Ratnieks 1993). Studying differences in egg-laying rates of anarchistic workers under queenless and queenright conditions is feasible, and could be used to test this hypothesis.
The third hypothesis for the differences in removal rates of eggs laid by queenright and queenless anarchistic workers is that few, if any, anarchistic workers mark their eggs when queenless. We surmise that the majority of workers that activate their ovaries when queenless do not establish the biochemical mechanism(s) to produce the egg-marking signal. There may be variance in the amount of egg-marking signal placed on eggs or in its quality, leading to a rapid decline in the acceptability of eggs as more and more workers become reproductively active, such that the survival of eggs laid by queenless anarchists approaches that of wild-type laying workers (Fig. 1). We do not argue that this is an adaptive switching-off of the production of the egg-marking signal. Rather, we suggest that it is an epiphenomenon of the other physiological and behavioural changes that occur in queenless workers (reviewed in Morgan et al. 1998; Simon et al. 2001), with the biochemical machinery used by queenright anarchistic workers being directed to the production of other compounds.
Our postulated hypotheses for the absence of the egg-marking signal in queenless anarchistic colonies remain speculative. To be able to fully understand its causes, we need a better understanding of the chemical nature of the signal, and how it is synthesised. We need to know how the nature and composition of egg-marking signals change as the influence of brood pheromones declines in a queenless colony. At present, we are far from this goal (Katzav-Gozansky et al. 2002; Oldroyd et al. 2002).
References
Arnold G, LeConte Y, Trouiller J, Hervet H, Chappe B, Masson C (1994) Inhibition of worker honeybee ovaries development by a mixture of fatty acid esters from larvae. C R Acad Sci Ser III Sci Vie 317:511–515
Barron AB, Oldroyd BP (2001) Social regulation of ovary activation in 'anarchistic' honey-bees (Apis mellifera). Behav Ecol Sociobiol 49:214–219
Barron AB, Oldroyd BP, Ratnieks FLW (2001) Worker reproduction in honey-bees (Apis) and the anarchistic syndrome: a review. Behav Ecol Sociobiol 50:199–208
Bourke AFG (1988) Worker reproduction in the higher eusocial Hymenoptera. Q Rev Biol 63:291–311
Châline N, Ratnieks FLW, Burke T (2002) Anarchy in the UK: detailed genetic analysis of worker reproduction in a naturally-occurring British anarchistic honeybee, Apis mellifera, colony using DNA microsatellites. Mol Ecol 11:1795–1803
Collett D (1994) Modelling survival data in medical research. Chapman & Hall, London
Dampney JR, Barron AB, Oldroyd BP (2003) Measuring the cost of worker reproduction in honey bees: work tempo in an 'anarchistic' line. Apidologie (in press)
Hamilton WD (1964) The genetical evolution of social behaviour. J Theor Biol 7:1–32
Katzav-Gozansky T, Soroker V, Ibarra F, Francke W, Hefetz A (2001) Dufour's gland secretion of the queen honeybee (Apis mellifera): an egg discriminator pheromone or a queen signal? Behav Ecol Sociobiol 51:76–86
Katzav-Gozansky T, Soroker V, Hefetz A (2002) Evolution of worker sterility in honey bees: egg-laying workers express queen-like secretions in Dufour's gland. Behav Ecol Sociobiol 51:588–589
Martin SJ, Beekman M, Wossler TC, Ratnieks FLW (2002) Parasitic Cape honeybee workers, Apis mellifera capensis, evade policing. Nature 415:163–165
Miller DG, Ratnieks FLW (2001) The timing of worker reproduction and breakdown of policing behaviour in queenless honey bee (Apis mellifera L.) societies. Insectes Soc 48:178–184
Mohammedi A, Paris A, Crauser D, LeConte Y (1998) Effect of aliphatic esters on ovary development of queenless bees (Apis mellifera L.). Naturwissenschaften 85:455–458
Montague CE, Oldroyd BP (1998) The evolution of worker sterility in honey bees: an investigation into a behavioral mutant causing failure of worker policing. Evolution 52:1408–1415
Morgan SM, Huryn VMB, Downes SR, Mercer AR (1998) The effects of queenlessness on the maturation of the honey bee olfactory system. Behav Brain Res 91:115–126
Oldroyd BP, Osborne KE (1999) The evolution of worker sterility in honeybees: the genetic basis of failure of worker policing. Proc R Soc Lond B 266:1335–1339
Oldroyd BP, Ratnieks FLW (2000) Evolution of worker sterility in honeybees (Apis mellifera): how anarchistic workers evade policing by laying eggs that have low removal rates. Behav Ecol Sociobiol 47:268–273
Oldroyd BP, Smolenski AJ, Cornuet J-M, Crozier RH (1994) Anarchy in the beehive. Nature 371:749
Oldroyd BP, Halling L, Rinderer TE (1999) Development and behaviour of anarchistic honeybees. Proc R Soc Lond B 266:1875–1878
Oldroyd BP, Halling LA, Good G, Wattanachaiyingcharoen W, Barron AB, Nanork P, Wongsiri S, Ratnieks FLW (2001a) Worker policing and worker reproduction in Apis cerana. Behav Ecol Sociobiol 50:371–377
Oldroyd BP, Wossler TC, Ratnieks FLW (2001b) Regulation of ovary activation in worker honey-bees (Apis mellifera): larval signal production and adult response thresholds differ between anarchistic and wild-type bees. Behav Ecol Sociobiol 50:366–370
Oldroyd BP, Ratnieks FLW, Wossler TC (2002) Egg-marking pheromones in honey-bees Apis mellifera. Behav Ecol Sociobiol 51:590–591
Ratnieks FLW (1988) Reproductive harmony via mutual policing by workers in eusocial Hymenoptera. Am Nat 132:217–236
Ratnieks FLW (1993) Egg-laying, egg-removal, and ovary development by workers in queenright honey bee colonies. Behav Ecol Sociobiol 32:191–198
Ratnieks FLW, Reeve HK (1992) Conflict in single-queen hymenopteran societies: the structure of conflict and processes that reduce conflict in advances eusocial species. J Theor Biol 158:33–65
Ratnieks FLW, Visscher PK (1989) Worker policing in the honeybee. Nature 342:796–797
Ratnieks FLW, Wossler TC, Neumann P, Oldroyd BP, Moritz RFA (2003) Egg laying and egg removal in honey bee colonies with different levels of anarchy. Insectes Soc (in press)
Simon UE, Moritz RFA, Crewe RM (2001) The ontogenetic pattern of mandibular gland components in queenless worker bees (Apis mellifera capensis Esch.). J Insect Physiol 47:735–738
Trouiller J, Arnold G, LeConte Y, Masson C, Chappe B (1991) Temporal pheromonal and kairomonal secretion in the brood of honeybees. Naturwissenschaften 78:368–370
Visscher PK (1996) Reproductive conflict in honey bees: a stalemate of worker egg-laying and policing. Behav Ecol Sociobiol 39:237–244
Acknowledgements
We wish to thank Michael Duncan for his beekeeping assistance, and the School of Biological Sciences for the use of the Crommelin Research Station. Financial support was obtained from Australian Research Council grants to M.B. and B.P.O. We thank Stephen Martin for his comments on the manuscript and members of the Social Insects Laboratory for stimulating discussions. The experiments described in this paper comply with the current laws of Australia.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by L. Keller
Rights and permissions
About this article
Cite this article
Beekman, M., Oldroyd, B.P. Different policing rates of eggs laid by queenright and queenless anarchistic honey-bee workers (Apis mellifera L.). Behav Ecol Sociobiol 54, 480–484 (2003). https://doi.org/10.1007/s00265-003-0647-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-003-0647-7