
Abstract
β-Glucan is a naturally occurring glucose polysaccharide with immunostimulatory activity in both infection and malignancy. β-Glucan’s antitumor effects have been attributed to the enhancement of complement receptor 3-dependent cellular cytotoxicity, as well as modulation of suppressive and stimulatory myeloid subsets, which in turn enhances antitumor T cell immunity. In the present study, we demonstrate antitumor efficacy of yeast-derived β-glucan particles (YGP) in a model of metastatic-like melanoma in the lung, through a mechanism that is independent of previously reported β-glucan-mediated antitumor pathways. Notably, efficacy is independent of adaptive immunity, but requires inflammatory monocytes. YGP-activated monocytes mediated direct cytotoxicity against tumor cells in vitro, and systemic YGP treatment upregulated inflammatory mediators, including TNFα, M-CSF, and CCL2, in the lungs. Collectively, these studies identify a novel role for inflammatory monocytes in β-glucan-mediated antitumor efficacy, and expand the understanding of how this immunomodulator can be used to generate beneficial immune responses against metastatic disease.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
β-Glucan is a biological response modifier with defined immunomodulatory properties [1]. It is a glucose polymer that naturally occurs in the cell walls of many fungi, yeast, seaweed, some bacteria, and some cereal grains. The most-studied immunoactive structure of β-glucan is a β-1,3 backbone of d-glucose with β-1,6 branches of variable length and complexity (hereafter, β-glucan) [2]. The myriad possible sources and preparations of β-glucan produce a diverse range of reported immunostimulatory activities, including protective effects against both infection [3] and malignancy [2]; however, the immunologic mechanisms that mediate antitumor immunity remain to be fully understood [4].
The most well-characterized mechanism of β-glucan-mediated antitumor efficacy acts through the receptor for complement-component 3 (CR3) to induce neutrophil or NK cell cytotoxic activity against tumors opsonized with inactivated complement component 3b (iC3b) [5, 6]. CR3 is expressed by many myeloid cells and NK cells, and its dual ligation by small, soluble β-glucan fragments and iC3b leads to cytotoxic granule release and/or phagocytosis by neutrophils and/or NK cells [5, 6]. β-Glucan enhances CR3-mediated killing of tumors with bound iC3b from naturally occurring Ab [6], and combining β-glucan with complement-fixing antitumor mAb enhances this activity in mouse tumor models [5] and humans [7]. Apart from one study reporting a failure of therapy in SCID mice [8], the antitumor efficacy of β-glucan has not been assessed in an Ab-deficient setting.
Recently, additional β-glucan-mediated antitumor effects, mediated through the C-type lectin receptor Dectin-1, have been reported; in these studies, β-glucan treatment alters the balance of stimulation vs. immunosuppression in ways that are beneficial to antitumor immune outcomes. For example, yeast β-glucan was shown to: improve the activation of DCs, generating superior T-cell stimulating APC [9]; convert suppressive M2 macrophages (Mφ) into stimulatory M1 Mφ [10]; and modulate the suppressive capacity of granulocytic and monocytic MDSCs [11, 12]. Bacterial β-glucan was shown to stimulate the conversion of Tregs into inflammatory Th17 cells [13, 14]. These studies implicate enhanced T cell immunity, likely an outcome of β-glucan’s signaling and activation of innate cells, as the proximal cause for β-glucan-induced antitumor immune responses; however, prior studies have not evaluated the potential direct role that innate cells may play in mediating adaptive-independent antitumor cytotoxic functions in response to β-glucan stimulation, including the potential role of monocytes (Mo).
Mo have been the focus of a recently described, β-glucan-mediated phenomenon termed trained innate immunity (TII), whereby β-glucan generates a primed state in innate immune cells that renders them more responsive to a subsequent, nonspecific stimulus [15]. TII was shown to mediate protection against infectious challenge and was associated with an altered metabolic state upon β-glucan priming, analogous to the well-characterized metabolic shift in activating lymphocytes [16]. TII’s metabolic shift was shown to be mediated by a Dectin-1-Akt-mTOR-HIF1α pathway, and interference with any of these components disrupted the effect [17]. While TII was elegantly demonstrated in infection models, it is not known whether these pathways are involved in β-glucan’s antitumor efficacy.
Therefore, we used a well-characterized murine tumor model [18] to assess the immunologic mechanisms underlying yeast-derived β-glucan particle (YGP)-mediated anti-tumor immunity. Our studies identify a novel role for inflammatory Mo in β-glucan-mediated antitumor efficacy, and suggest this immunomodulator may be useful as an adjuvant to enhance immune responses against metastatic disease.
Materials and methods
Cell lines
B16.F10 cells (CRL-6475) were obtained from ATCC, then expanded and cryopreserved to create individual aliquots for each study. B16.F10-luciferase cells were a gift from Dr. Yina Huang (Dartmouth); briefly, Click beetle green luciferase was cloned into an MSCV-based retrovirus (pCMV2.1, a gift from Dr. Bill Sha [UC Berkeley]) and packaged in 293T cells; viral supernatants were used to transduce B16.F10 melanoma cells followed by selection with 2 μg/ml puromycin (InvivoGen, San Diego, CA). All tumor cells were maintained in cRPMI [RPMI-1640 medium (Hyclone/GE Healthcare Life Science, Pittsburgh, PA) containing 10% FBS (Gemini Bio Products, Sacramento, CA) and 1.5% HEPES (Thermo Fisher Scientific, Waltham, MA) buffer] at 37 °C and 5% CO2.
Animals and tumor model
C57BL/6J (stock no. 000664) and CCR2−/− mice (stock no. 004999) mice were from Jackson Laboratories (Bar Harbor, ME). CCR2.DTR mice [16] were a gift from Dr. Tobias Hohl (Memorial Sloan Kettering Cancer Center). Myeloid-HIF1α−/− mice (LysM-Cre floxed-HIF1α exon 2) [17, 19] were bred in house at Dartmouth. All animals were maintained in standard, specific pathogen-free housing conditions, and 8–12 week old mice were used for all studies. Metastatic-like melanoma in the lungs was induced by injection of 3 × 105 B16.F10 melanoma cells, as described [20].
Reagents
YGP was generated from Saccharomyces cerevisiae, as described [21]. Dry glucan particles were resuspended to 10 mg/mL in sterile PBS and injected in 100 μL (1 mg) i.p. on days −7 and −4, relative to tumor challenge (day 0).
Depleting antibodies against NK cells (αNK1.1, clone PK136) and neutrophils (αLy6G, clone 1A8) were purchased from Bio X Cell (West Lebanon, NH). For depletions, 50 μg/mouse αNK1.1 was administered i.p. on days − 6 and − 2 in 200µL PBS, or 200 μg/mouse. αLy6G was administered i.p. on days − 8, −6, −4, and − 2 in 200 μL PBS.
Rapamycin (Sigma Aldrich, St Louis, MO; cat no. R0395) was dissolved in vehicle [0.2% carboxymethylcellulose (SigmaC-5013, high viscosity) and 0.25% polysorbate 80 (Sigma P-8074) in PBS]. Rapamycin was administered i.p. in 200 μL of 0.4 mg/mL solution; mice were treated every 2 days beginning day − 9, as described [22]. Diphtheria toxin (DT); (List Biological Laboratories, Campbell, CA; cat no. 150) was administered to mice i.p. on days − 8, − 6, and − 4 at a dose of 2 ng/g of body weight [16].
RNA extraction and tyrosinase qPCR
RNA from the left lung was generated and reverse-transcribed, as described [18]. Tyrosinase gene expression was quantified by qPCR using master mix and pre-validated gene-specific TaqMan primers (Thermo Fisher), then analyzed on a StepOne Plus instrument (Applied Biosystems). Tyrosinase (Thermo Fisher Mm00453201_m1) was normalized to GAPDH (Thermo Fisher Mn99999915_g1) or β-actin (Thermo Fisher Mm02619580_g1), and fold-expression data were analyzed using the ΔΔCt method.
Flow cytometry
Perfused right lungs were dissociated into single-cell suspensions and stained, as described [18]. Prior to staining, cells were Fc blocked with αCD16/32 (clone 93, BioLegend, San Diego, CA), then incubated with indicated antibodies for 30 min at 4 °C in the dark. Antibodies specific for mouse CD45 (clone 30-F11), CD11b (clone M1/70), Ly6C (clone HK1.4), Ly6G (clone 1A8), CD4 (GK1.5), and CD8 (53 − 6.7) were obtained from Biolegend (San Diego, CA). Live/dead fixable violet cell stain was obtained from Thermo-Fisher. Data were acquired with a MacsQuant 10 cytometer (Miltenyi) and analyzed using FlowJo (Ashland, OR) software.
Cytotoxicity assay
Mo were isolated from bulk C57BL/6J BM utilizing the Mouse Monocyte Isolation kit (Miltenyi, cat no. 130-100-629), then were trained in vitro with β-glucan, based on published methods [23]. Briefly, Mo were plated at 1 × 106 cells/mL in 10 mL Mo media (RPMI with 10% FBS, 55 µM 2-mercaptoethanol, 1× HEPES, 1 mM sodium pyruvate, 1x penicillin/streptomycin, and 2 mM l-glutamine) and stimulated for 24 h with RPMI or 5 µg/mL β-glucan. After 24 h, cells were washed and rested for 3 days in Mo media, activated with 10 ng/mL LPS for 24 h, then harvested and washed for plating with target cells. B16.F10-luciferase were plated 2 × 103 cells/well in a 96-well opaque white plate, then rested for 6 h in a 37 °C and 5% CO2 in cRPMI plus 2 µg/mL puromycin. After 6 h, media was changed to Mo media, and Mo were plated at 2 × 105 cells/well, 200 µL total volume per well. Plates were incubated at 37 °C 5% CO2 for 24 h, control lysis wells were lysed with Lysis solution from the CytoTox 96 kit (Promega, Madison, WI; cat no. G1780), and 50 μL of 200 µg/mL luciferin (in PBS) was added to each well, incubated at 37 °C for 30 min, then luminescence was read on a Molecular Devices (Sunnyvale, CA) SpectraMax i3x. Percent specific lysis was calculated using the formula: (1-percent viability) × 100, where percent viability was calculated as the [luminescence of the experimental sample minus background (luminescence of lysed target cells)], divided by [maximum viability (luminescence of unlysed target cells) minus background] [24].
Multiplex protein assays
Perfused lungs were homogenized with a rotor–stator homogenizor in T-PER buffer (Thermo Fisher cat no. 78510) supplemented with protease inhibitor tablets (Roche, Mannheim, DE; cat no. 11697498001) and Y-30 antifoam (Sigma Aldrich cat no. A6457). The T-PER protocol was followed for protein isolation. Blood was collected in heparinized microcentrifuge tubes and centrifuged at 1125×g for 10 min, with the supernatant then centrifuged at 1825×g for 10 min, and the plasma collected again. Cytokine concentrations were normalized to total protein quantified with the Pierce BCA Protein Assay Kit (Thermo Fisher cat no. 23225). The Luminex was performed using the EMD-Millipore (Billerica, MA) mouse 32-plex kit.
Statistics
Graphing and statistical analyses were performed using GraphPad Prism software (La Jolla, CA). Data are presented as means ± standard deviation. Data for experimental groups were compared using the Mann–Whitney U test. Statistical significance is indicated as *p < 0.05, **p < 0.01. ***p < 0.001. ****p < 0.0001. Data are representative of at least 2 independent experiments.
Results
β-Glucan mediates antitumor efficacy in a model of metastatic melanoma
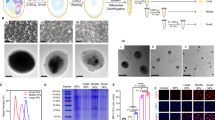
To evaluate the effect of β-glucan priming in our model of metastatic melanoma, we utilized a prophylactic treatment protocol [17] (schema in Fig. 1a) with YGP β-glucan. In lungs 14-days post-tumor challenge, visible tumor burden was reduced by YGP treatment (Fig. 1b). YGP treatment significantly (p < 0.05) reduced tyrosinase expression in lungs [25], compared to PBS-treated controls (Fig. 1c). To discount the possibility that the observed differences in tumor burden were due to a defect in initial tumor engraftment, we evaluated tyrosinase expression 2 h post-tumor challenge, by which time B16 is known to have efficiently engrafted the lung [26]; there was no difference in tyrosinase expression (Fig. 1d), discounting the hypothesis that β-glucan treatment influences initial tumor engraftment.

β-Glucan mediates antitumor efficacy in a model of metastatic melanoma. a Experimental scheme: C57BL/6J mice were given β-glucan (YGP) i.p. on days − 7 and − 4, and challenged with 3 × 105 B16.F10 i.v. on day 0. Tumor burden was evaluated at various timepoints up to 14 days after tumor challenge; b Lung photos at day 14; c Tyrosinase qPCR of total lung RNA of lungs shown in b, plus non-tumor bearing control lungs not pictured; d Mice treated as in a. Tyrosinase qPCR of total lung RNA 2 h after tumor inoculation (as in a), plus non-tumor bearing control lungs; e Tyrosinase qPCR of total lung RNA 48 h after tumor inoculation (as in a), plus non-tumor bearing control lungs
While β-glucan-mediated antitumor efficacy was apparent in a long-term study, it remained uncertain whether β-glucan priming was impacting early stages of tumor development. Therefore, we evaluated tumor burden 48 h post-tumor challenge. We observed a significant (p < 0.05) reduction in tumor burden in YGP-treated mice at 48 h (Fig. 1e), indicating that β-glucan-induced immunologic anti-tumor activity occurs rapidly after tumor engraftment. Consequently, these data suggested an innate immune-mediated mechanism.
β-Glucan induces an early accumulation of myeloid cells in the lung
To gain insight into the immune cells responsible for β-glucan’s antitumor efficacy in our model, we assessed changes in lung immune cellularity. Our gating strategy for flow cytometry (Fig. 2a) discriminated singlets and live CD45+ cells, which we analyzed for either CD4 and CD8 expression to evaluate T cell populations, or CD11b and subsequently Ly6C and Ly6G expression to evaluate myeloid populations. Consistent with prior reports, we defined CD11b+Ly6G−Ly6Chi cells as inflammatory Mo, CD11b+Ly6G+Ly6Cmid cells as neutrophils, and CD11b+Ly6C−Ly6G− cells as Ly6C− Mo/Mφ [27].

β-Glucan induces an early expansion of myeloid cells in the lung. Mice were treated as in Fig. 1a, and lungs were harvested at the indicated time point; the right lung was perfused and processed into single-cell suspensions and stained for flow cytometry. a Gating strategy to identify CD4 and CD8 T cells (live singlet CD45+CD11b+CD45+CD4+ or 8+), neutrophils (live singlet CD45+CD11b+Ly6C−Ly6G+), inflammatory monocytes (live singlet CD45+CD11b+Ly6ChiLy6G−), and Ly6C− Mo/Mφ (live singlet CD45+CD11b+Ly6C−Ly6G−); b Total right lobe numbers of the indicated populations, without tumor inoculation; c Total right lobe numbers of each population at 2 h post B16 i.v. tumor challenge; d Total right lobe numbers of each population at 14 days post-tumor challenge. Each panel includes representative data from one of several similar experiments with comparable results
YGP treatment alone (no tumor) induced an accumulation of neutrophils (p < 0.05), a strongly trending increase in inflammatory Mo (p = 0.057), and a trend towards increased Ly6C−Mo/Mφ, in the lungs compared to PBS-treated mice. In contrast, we observed no differences in CD4 or CD8 T cell numbers in this treatment group (Fig. 2b). Introduction of tumor did not substantially change the pattern of cellular accumulation at 2 h post-tumor challenge, with significant increases (p < 0.05) in neutrophils, inflammatory Mo, and Ly6C−Mo/Mφ, and no changes in T cell numbers, in YGP-treated mice (Fig. 2c and Supplemental Fig. 1a). At both 0 h (without tumor) and 2 h post-tumor, cellularity in the spleen mirrored the lung (data not shown), indicating that β-glucan’s effect on myeloid cells was not restricted to the lung. By the terminal 14-day time point, treatment-induced changes in myeloid populations had contracted, and no differences in neutrophils or Mo were observed (Fig. 2d and Supplemental Fig. 1b). However, CD4 and CD8 T cell numbers were significantly (p < 0.05) decreased (Fig. 2d), perhaps consequent to reduced tumor burden. Together, these data demonstrate an early and transient β-glucan-induced accumulation of myeloid cells in the lung, with comparatively little effect on T cells.
β-Glucan stimulates inflammatory mediator expression in the lung and circulation
We next investigated the broad immune impact of β-glucan treatment, both in the lung environment and systemically, using a multiplex cytokine assay (Luminex) on lung homogenates and plasma of YGP-treated mice, both before and 24 h-post tumor challenge (Fig. 3). YGP treatment increased the levels of TNFα (Fig. 3a, p < 0.05) and CCL2 (Fig. 3b, trending) in the lung and plasma, notably only in the presence of tumor. This is suggestive that β-glucan primes immune cells for enhanced activity in the presence of tumor. YGP treatment increased some cytokines, regardless of tumor, including IL-6 in plasma (Fig. 3c) and G-CSF in plasma and lung tissue (Fig. 3d). Interestingly, YGP decreased circulating levels of M-CSF regardless of tumor presence, whereas M-CSF was increased in the lungs of tumor-bearing, YGP treated mice (Fig. 3e). YGP treatment had no impact on the immunomodulatory cytokine IL-10 in the lungs (Fig. 3f); IL-10 in serum was below the limit of detection. These data support the concept that β-glucan primes the lung compartment with proinflammatory cytokines that may enable rapid and effective innate responses to engrafting tumor cells, likely through the induction of Mo-mediated antitumor mechanisms.

β-Glucan stimulates inflammatory mediator expression in the lung and circulation. Mice were treated with β-glucan as in Fig. 1a, and plasma and total lung protein were harvested at day 0 (no tumor), and 24 h post B16 challenge. Samples were analyzed using a Millipore mouse multiplex kit on the Luminex platform. Normalized protein concentrations shown for a TNFα; b CCL-2; c IL-6; d G-CSF; e M-CSF; and f IL-10. Measurements were normalized to total protein values obtained from a BCA total protein assay
β-Glucan-mediated protection against metastatic melanoma does not require adaptive immunity, NK cells or neutrophils
While much of the literature reporting an antitumor effect of β-glucan has implicated Ab [5, 6, 28] or T cell activation [11, 29], no study has evaluated whether the therapeutic effect is maintained in animals lacking B and T cells. To determine whether adaptive immunity contributes to β-glucan anti-tumor efficacy in our model, we assessed tumor development in YGP-treated RAG-deficient animals. Strikingly, the strong protective effect of YGP treatment was maintained (Fig. 4a), and the quantification of tumor burden confirmed the reduction (Fig. 4b, p < 0.05), demonstrating an innate-intrinsic antitumor effect of β-glucan in our model.

β-Glucan-mediated protection against metastatic melanoma does not require adaptive immunity, NK cells or granulocytes. Mice were treated as in Fig. 1a, and lungs were harvested 14 days after i.v. B16 tumor challenge; a Lung photos from RAG−/− mice; b Tyrosinase qPCR of total lung RNA from RAG−/− lungs shown in a; c C57BL/6J mice were treated as in Fig. 1a, with some also given αNK1.1 Ab i.p. on days − 6 and − 2; d Tyrosinase qPCR of total lung RNA from lungs shown in c; e C57BL/6J mice were treated as in Fig. 1a, with some also given αLy6G Ab on days − 8, -6, -4, and − 2; f Tyrosinase qPCR of total lung RNA from lungs shown in e
NK cells have also been implicated in β-glucan-mediated antitumor effects, including studies that evaluated metastatic melanoma [30, 31]. To assess the role of NK cells in our model, we used specific depleting Ab, and confirmed depletion by flow cytometry (data not shown). Although control mice had a lower baseline tumor engraftment in this study, the protective effect of YGP treatment was still apparent (Fig. 4c, d). Consistent with prior work [32], NK depletion dramatically increased baseline tumor engraftment, but YGP treatment reduced the visible tumor burden in NK-depleted mice (Fig. 4c) and modestly but consistently reduced tyrosinase expression (Fig. 4d), suggesting that NK cells are not major contributors to the anti-tumor efficacy of β-glucan treatment.
Finally, neutrophils have been implicated in the antitumor mechanism of β-glucan [33]. To assess their requirement for protective effect in our model, we depleted neutrophils using a Ly6G-depleting Ab, and confirmed depletion with flow cytometry (data not shown). YGP was equally protective against lung metastatic-like lesions, regardless of the presence of neutrophils (Fig. 4e, f). Thus, adaptive immunity is dispensable, at least in part, for β-glucan-mediated antitumor efficacy in our model of metastatic melanoma.
β-Glucan’s antitumor efficacy is independent of the mTOR-HIF1α trained innate immunity (TII) mechanism
Recently, β-glucan has been shown to induce a primed state of innate immunity that responds more vigorously to a subsequent, non-specific stimulus. This phenomenon, termed TII, has been shown to be protective in infection models, though the potential role in β-glucan-induced antitumor immunity remains unknown. Therefore, we assessed whether β-glucan’s anti-tumor efficacy against metastatic-like lung tumors was mediated through a classical TII mechanism. First, we repeated our YGP treatment and tumor challenge in mice with a myeloid-specific knockout of HIF1α (mHIF1α−/−) [19], shown to be required for Candida albicans β-glucan-induced TII in infection models [17]. We note here that our model utilized YGP from Saccharomyces cerevisiae [21]. Fourteen days after tumor challenge, YGP treatment reduced tumor burden in mHIF1α−/− mice, both by gross visual assessment and by tyrosinase expression (p < 0.05) (Supplemental Fig. 2a, b). Because mHIF1α−/− mice showed an impaired baseline engraftment compared to wild-type mice, we compared the average YGP-induced reduction in tumor burden between experiments with wild-type and mHIF1α−/− mice, and found that the fold-reduction in tumor burden in mHIF1α−/− mice exceeded that seen in wild-type mice (Supplemental Fig. 2c). Thus, there was no impairment of protection, despite the lower baseline engraftment, in mHIF1α−/− mice.
As HIF1α is downstream of mTOR, we assessed rapamycin treatment as an alternative for the blockade of TII activity. Rapamycin treatment failed to diminish β-glucan’s protective effect (Supplemental Fig. 2d, e), in conceptual concordance with the mHIF1α−/− data. Thus, we exclude classical TII mechanisms from a role in β-glucan’s anti-tumor efficacy in our model.
Inflammatory monocytes are required for β-glucan’s efficacy against metastatic melanoma, and β-glucan enhances monocyte cytotoxicity against B16 in vitro
Inflammatory Mo were increased in the lungs of β-glucan-treated mice, but no study has assessed the role of these cells in mediating the antitumor effects of β-glucan. Further, although Mo (and their derivative Mφs) are usually ascribed a pro-tumor role [31], a recent paper reported an antitumor effector role for inflammatory Mo in a murine melanoma model [32], further intriguing us as to the possible role of these cells. Therefore, we first utilized CCR2−/− mice [34]; Mo from these donors are known to be deficient in egress from BM and trafficking to peripheral sites of inflammation. Whereas YGP treatment significantly (p < 0.05) reduced tumor burden in wild-type mice, it had no effect in CCR2−/− animals, apparent visually and quantified by tyrosinase expression (Fig. 5a, b). To confirm the role of inflammatory Mo, we utilized inflammatory Mo-depleter mice, which express the DT receptor under the control of the CCR2 promoter [16]. We confirmed DT-mediated depletion of Ly6Chi Mo by flow cytometry (data not shown). In accord with data from the CCR2−/− model, β-glucan’s efficacy was abrogated in Mo-depleted mice. Curiously, Mo depletion resulted in higher baseline tumor engraftment in the lung; nevertheless, the tumor burden was unchanged in β-glucan treated animals (Fig. 5c, d).

Inflammatory monocytes are required for β-glucan’s efficacy against metastatic melanoma, and β-glucan enhances monocyte cytotoxicity against B16 in vitro. Mice were treated as in Fig. 1a, and lungs were harvested at day 14 post B16 i.v. a Day 14 lung photos from WT C57BL/6J or CCR2−/− mice treated as in Fig. 1a, b Tyrosinase qPCR of total lung RNA of lungs shown in a; c WT C57BL/6J or CCR2.DTR+/− mice were treated as in Fig. 1a, with the CCR2.DTR mice also receiving DT i.p. on days − 8, − 6, and − 4. Day 14 lung photos shown; d Tyrosinase qPCR of total lung RNA from lungs shown in c; e BM-derived monocytes were isolated and stimulated with either RPMI or β-glucan for 24 h, followed by a 3-day resting period, activated with LPS for 24 h, then plated with B16-luciferase for 24 h at a monocyte:B16 ratio of 100:1. After 24 h of co-culture luciferin was added and luminescence intensity determined. See methods for % lysis calculation
We recognized that the anti-tumor activity of β-glucan-induced Mo may be direct or indirect (including chemotactic factor or proinflammatory cytokine production). To address whether Mo may be directly mediating antitumor effector functions, we adapted an in vitro cytotoxicity assay [35], using luciferase-expressing B16 (B16.F10-luciferase) as target cells, to quantify Mo-mediated cytotoxicity. In vitro YGP-stimulated BM-derived Mo (characterized as CD45+CD11b+Ly6G−Ly6Chi by flow cytometry, data not shown) exhibited roughly twofold greater cytotoxicity toward B16 cells, compared to control-stimulated Mo (Fig. 5e). These data demonstrate that β-glucan enhances Mo cytotoxicity toward B16 in vitro and support a previously unappreciated mechanism for β-glucan-induced tumor control, through Mo accumulation and activation in situ.
Discussion
We demonstrate a striking antitumor effect of particulate β-glucan (YGP) treatment in a murine model of pulmonary-metastatic melanoma, independent of previously reported β-glucan antitumor mechanisms. These findings define a novel Mo-dependent mechanism by which β-glucan mediates antitumor efficacy against metastatic disease in the lungs; further, whereas Mo and Mφ in the tumor microenvironment are usually characterized as pro-tumor, we demonstrate the capacity of inflammatory Mo as antitumor effectors.
Prior literature has reported that β-glucan “decreased engraftment” or reduced metastasis [36], when in fact, the experiments (assessment of terminal tumor load) cannot distinguish between changes in engraftment and immune-mediated tumor control. Our studies, showing that prophylactic β-glucan treatment led to a striking reduction in pulmonary tumor burden but no changes in engraftment, suggest an early innate immune-driven mechanism.
Of the reported mechanisms for β-glucan-mediated antitumor efficacy, two prevail, and our data are inconsistent with both. First is the well-established mechanism by which β-glucan binds to and primes CR3 for enhanced phagocytosis and cytotoxicity of iC3b-opsonized target cells [28, 37]. The primary CR3-expressing cell types reported to be responsible for this are NK cells and neutrophils, both of which were likely dispensable for the induction of antitumor immunity in our model. In addition to CR3-expressing effector cells, this mechanism requires deposition of iC3b on tumor cells, and this is reported to require tumor-specific, complement-fixing antibody, either naturally occurring or administered as mAb [6, 28]. β-glucan’s efficacy in our studies is unaffected in RAG−/− mice that lack functional mature B cells. Unless B16.F10 has a ligand for Mannose Binding Lectin, which to our knowledge is not known, and iC3b deposition is being triggered in that manner, the CR3-iC3b mechanism of β-glucan antitumor efficacy is not likely to be involved in our model. It should be noted that the lectin-binding site of CR3 binds soluble β-glucan fragments, while we used particulate YGP in our study; prior work has shown that glucan particles are phagocytosed and processed into smaller soluble fragments in the BM, and thereafter available to bind to and prime CR3 in the same manner as soluble fragments [38]. Thus, our use of particulate glucan cannot alone explain why the CR3-iC3b mechanism and its cellular effectors are not responsible for anti-tumor efficacy.
The other mechanism implicated in β-glucan-mediated anti-cancer efficacy is stimulation of antitumor T cell immunity. Many studies have documented the activation or enhancement of T cell immunity through the release of suppression from MDSC, M2 Mφ, or Tregs, and the stimulation of DC to be more efficient APC [9,10,11,12, 39]. However, as with the requirement for Ab, our RAG−/− experiments clearly demonstrate the dispensability of T cells for β-glucan’s efficacy in our model. To our knowledge, this is the first study reporting antitumor efficacy of β-glucan in RAG−/− mice, an important result given the central role that Ab and T cells play in the previously-reported mechanisms of β-glucan’s antitumor effects.
β-Glucan is known to prime cells for an enhanced response to nonspecific stimuli in a TII-like manner [40]. This is known to be protective against infection, and the mechanism was recently shown to involve a metabolic shift to aerobic glycolysis in trained cells, requiring elements of the Dectin-1-Akt-mTOR-HIF1α pathway, specifically in myeloid cells [17]. This mechanism of β-glucan-mediated protection has not been evaluated for involvement in β-glucan-mediated antitumor efficacy. Our study demonstrated the maintenance of β-glucan’s antitumor efficacy in mHIF1α−/− mice, and in mice treated systemically with rapamycin, both of which abrogated β-glucan’s protective effect in infection models. Thus, we found no evidence that the metabolic shift of TII is involved in our tumor model, however, we note the use of Saccharomyces cerevisiae-derived β-glucan [21] in our studies and Candida albicans-derived β-glucan in the TII models [17]. Dectin-1 is, however, known to activate distinct signaling pathways, including the canonical Syk-CARD9 pathway and the more recently described non-canonical Raf-1-Syk-independent pathway, in addition to the PI3K-Akt-mTOR-HIF1α signaling discussed above as the basis for TII [41, 42]. Thus, the mTOR-HIF1α TII pathway is not required for antitumor efficacy in our model, necessitating further investigation to determine the requisite signaling for the effects observed.
Mo are among the innate cells known to be responsive to β-glucan, though tumor studies have not specifically evaluated the role of Mo in mediating β-glucan’s antitumor effects. Rather, the immunotherapy literature generally reports Mo and Mφ in the context of immunosuppression, for example via M2 Mφ and MDSC [10, 12]. CCR2 is a chemokine receptor that is well established as the means of inflammatory Mo egress from bone marrow, as well as a defining marker for inflammatory Mo, along with high Ly6C expression. We show a failure of therapy in CCR2−/− mice, whose primary immunodeficiency is impaired Mo egress from BM and trafficking to peripheral sites of inflammation [43]. To bolster this finding with another Mo-interfering approach, we utilized CCR2.DTR mice, allowing us to deplete inflammatory Mo during the β-glucan-treatment period, and we again observed an abrogation of β-glucan’s efficacy. While both of these approaches have limitations (i.e., collateral effects on other CCR2-expressing populations), these well-accepted models of inflammatory Mo deficiency demonstrate a requirement for inflammatory Mo for β-glucan’s effect. Although our models are yet to establish a direct effect of the Mo against tumor in vivo, our in vitro cytotoxicity data demonstrate that β-glucan increases the cytotoxic capacity of Mo towards B16.F10. The potential of inflammatory Mo as antitumor effector cells is not without precedent: a recent paper described inflammatory Mo as essential effector cells in the mouse MT/ret melanoma model, independent of T cell- or NK cell-mediated immunity. They further demonstrated in vitro cytotoxicity of inflammatory Mo, which was most sensitive to ROS inhibition, and less so to TNFα inhibition [32]. Another study demonstrated an increase in circulating inflammatory Mo numbers and activation markers in advanced breast cancer patients after taking oral β-glucan for 2 weeks, which is a small but important clinical correlate supporting the potential role of inflammatory Mo in β-glucan therapy [43].
Our studies demonstrate a previously undescribed role for inflammatory Mo in mediating the antitumor effects of β-glucan, wholly or partly independent of known antitumor mediators of β-glucan including adaptive immunity, NK cells and neutrophils. We show in vitro evidence that β-glucan enhances the cytotoxic potential of Mo, and identify potential inflammatory mediators in vivo including CCL2, TNFα and M-CSF. We do not exclude the possibility of direct β-glucan effects on tumor cells, or multi-parameter interactions between primed monocytes and tumors, which collectively limit tumor growth. However, the present data are an important addition to the understanding of how particulate β-glucan mediates antitumor efficacy, and future immunotherapeutic applications of β-glucan may be able to harness the antitumor potential of inflammatory Mo, particularly in the context of advanced and often refractory metastatic disease.
Abbreviations
- β-Glucan:
-
β-1,3 Backbone of d-glucose with β-1,6 branches of variable length and complexity
- CR3:
-
Complement receptor 3
- DT:
-
Diphtheria toxin
- iC3b:
-
Inactivated complement component 3b
- Mφ:
-
Macrophages
- Mo:
-
Monocytes
- mHIF1α−/− :
-
Myeloid-specific knockout of HIF1α
- TII:
-
Trained innate immunity
- YGP:
-
Yeast-derived β-glucan particles
References
Aleem E (2013) β-Glucans and their applications in cancer therapy: focus on human studies. Anticancer Agents Med Chem 13:709–719
Vannucci L, Krizan J, Sima P et al (2013) Immunostimulatory properties and antitumor activities of glucans. Int J Oncol 43:357–364. https://doi.org/10.3892/ijo.2013.1974 (review)
Rahar S, Swami G, Nagpal N et al (2011) Preparation, characterization, and biological properties of β-glucans. J Adv Pharm Technol Res 2:94–103. https://doi.org/10.4103/2231-4040.82953
Chan G, Chan W, Sze D (2009) The effects of β-glucan on human immune and cancer cells. J Hematol Oncol 2:25. https://doi.org/10.1186/1756-8722-2-25
Hong F, Yan J, Baran JT et al (2004) Mechanism by which orally administered beta-1,3-glucans enhance the tumoricidal activity of antitumor monoclonal antibodies in murine tumor models. J Immunol 173:797–806. https://doi.org/10.4049/jimmunol.173.2.797
Vetvicka V, Thornton BP, Wieman TJ, Ross GD (1997) Targeting of natural killer cells to mammary carcinoma via naturally occurring tumor cell-bound iC3b and beta-glucan-primed CR3 (CD11b/CD18). J Immunol 159:599–605
Cheung N-KV, Modak S, Vickers A, Knuckles B (2002) Orally administered beta-glucans enhance anti-tumor effects of monoclonal antibodies. Cancer Immunol Immunother 51:557–564. https://doi.org/10.1007/s00262-002-0321-3
Yan J, Vetvicka V, Xia Y et al (1999) Beta-glucan, a “specific” biologic response modifier that uses antibodies to target tumors for cytotoxic recognition by leukocyte complement receptor type 3 (CD11b/CD18). J Immunol 163:3045–3052
Li B, Cai Y, Qi C et al (2010) Orally administered particulate beta-glucan modulates tumor-capturing dendritic cells and improves antitumor T-cell responses in cancer. Clin Cancer Res 16:5153–5164. https://doi.org/10.1158/1078-0432.CCR-10-0820
Liu M, Luo F, Ding C et al (2015) Dectin-1 activation by a natural product β-glucan converts immunosuppressive macrophages into an M1-like phenotype. J Immunol 195:5055–5065. https://doi.org/10.4049/jimmunol.1501158
Tian J, Ma J, Ma K et al (2013) β-Glucan enhances antitumor immune responses by regulating differentiation and function of monocytic myeloid-derived suppressor cells. Eur J Immunol 43:1220–1230. https://doi.org/10.1002/eji.201242841
Albeituni SH, Ding C, Liu M et al (2016) Yeast-derived particulate β-glucan treatment subverts the suppression of myeloid-derived suppressor cells (MDSC) by Inducing polymorphonuclear MDSC apoptosis and monocytic MDSC differentiation to APC in cancer. J Immunol 196:2167–2180. https://doi.org/10.4049/jimmunol.1501853
Osorio F, LeibundGut-Landmann S, Lochner M et al (2008) DC activated via dectin-1 convert Treg into IL-17 producers. Eur J Immunol 38:3274–3281. https://doi.org/10.1002/eji.200838950
LeibundGut-Landmann S, Osorio F, Brown GD, Reis e Sousa C (2008) Stimulation of dendritic cells via the dectin-1/Syk pathway allows priming of cytotoxic T-cell responses. Blood 112:4971–4980. https://doi.org/10.1182/blood-2008-05-158469
Netea MGG, Quintin J, van der Meer JWM, van der Meer JWM (2011) Trained immunity: a memory for innate host defense. Cell Host Microbe 9:355–361. https://doi.org/10.1016/j.chom.2011.04.006
Hohl TM, Rivera A, Lipuma L et al (2009) Inflammatory monocytes facilitate adaptive CD4 T cell responses during respiratory fungal infection. Cell Host Microbe 6:470–481. https://doi.org/10.1016/j.chom.2009.10.007
Cheng S-C, Quintin J, Cramer RA et al (2014) mTOR- and HIF-1α-mediated aerobic glycolysis as metabolic basis for trained immunity. Science 345:1250684. https://doi.org/10.1126/science.1250684
Clancy-Thompson E, Perekslis TJ, Croteau W et al (2015) Melanoma induces, and adenosine suppresses, CXCR3-cognate chemokine production and T-cell infiltration of lungs bearing metastatic-like disease. Cancer Immunol Res. https://doi.org/10.1158/2326-6066.CIR-15-0015
Cramer T, Yamanishi Y, Clausen BE et al (2003) HIF-1alpha is essential for myeloid cell-mediated inflammation. Cell 112:645–657
Clancy-Thompson E, Perekslis TJ, Croteau W et al (2015) Melanoma induces, and adenosine suppresses, CXCR3-cognate chemokine production and T-cell infiltration of lungs bearing metastatic-like disease. Cancer Immunol Res 3:956–967. https://doi.org/10.1158/2326-6066.CIR-15-0015
Huang H, Ostroff GR, Lee CK et al (2012) Relative contributions of dectin-1 and complement to immune responses to particulate β-glucans. J Immunol 189:312–317. https://doi.org/10.4049/jimmunol.1200603
Wang X, Wang W, Xu J, Le Q (2013) Effect of rapamycin and interleukin-2 on regulatory CD4+CD25+Foxp3+ T cells in mice after allogenic corneal transplantation. Transplant Proc 45:528–537. https://doi.org/10.1016/j.transproceed.2012.06.064
Bekkering S, Blok BA, Joosten LAB et al (2016) In vitro experimental model of trained innate immunity in human primary monocytes. Clin Vaccine Immunol 23:926–933. https://doi.org/10.1128/CVI.00349-16
Liu C, Yu S, Kappes J et al (2007) Expansion of spleen myeloid suppressor cells represses NK cell cytotoxicity in tumor-bearing host. Blood 109:4336–4342. https://doi.org/10.1182/blood-2006-09-046201
Butler KL, Clancy-Thompson E, Mullins DW (2017) CXCR3+ monocytes/macrophages are required for establishment of pulmonary metastases. Sci Rep 7:45593. https://doi.org/10.1038/srep45593
Sorensen MR, Pedersen SR, Lindkvist A et al (2014) Quantification of B16 melanoma cells in lungs using triplex Q-PCR—a new approach to evaluate melanoma cell metastasis and tumor control. PLoS One 9:e87831. https://doi.org/10.1371/journal.pone.0087831
Misharin AV, Morales-Nebreda L, Mutlu GM et al (2013) Flow cytometric analysis of macrophages and dendritic cell subsets in the mouse lung. Am J Respir Cell Mol Biol 49:503–510. https://doi.org/10.1165/rcmb.2013-0086MA
Hong F, Hansen RD, Yan J et al (2003) Beta-glucan functions as an adjuvant for monoclonal antibody immunotherapy by recruiting tumoricidal granulocytes as killer cells. Cancer Res 63:9023–9031
Yoon TJ, Kim TJ, Lee H et al (2008) Anti-tumor metastatic activity of β-glucan purified from mutated Saccharomyces cerevisiae. Int Immunopharmacol 8:36–42. https://doi.org/10.1016/j.intimp.2007.10.005
Ballas ZK, Buchta CM, Rosean TR et al (2013) Role of NK cell subsets in organ-specific murine melanoma metastasis. PLoS One 8:e65599. https://doi.org/10.1371/JOURNAL.PONE.0065599
Noy R, Pollard JW (2014) Tumor-associated macrophages: from mechanisms to therapy. Immunity 41:49–61. https://doi.org/10.1016/j.immuni.2014.06.010
Pommier A, Audemard A, Durand A et al (2013) Inflammatory monocytes are potent antitumor effectors controlled by regulatory CD4+ T cells. Proc Natl Acad Sci 110:13085–13090. https://doi.org/10.1073/pnas.1300314110
Vetvicka V, Vetvickova J (2015) Glucan supplementation has strong anti-melanoma effects: role of NK cells. Anticancer Res 35:5287–5292
Boring L, Gosling J, Chensue SW et al (1997) Impaired monocyte migration and reduced type 1 (Th1) cytokine responses in C–C chemokine receptor 2 knockout mice. J Clin Invest 100:2552–2561. https://doi.org/10.1172/JCI119798
Fu X, Tao L, Rivera A et al (2010) A simple and sensitive method for measuring tumor-specific T cell cytotoxicity. PLoS One 5:e11867. https://doi.org/10.1371/journal.pone.0011867
Yamamoto K, Kimura T, Sugitachi A, Matsuura N (2009) Anti-angiogenic and anti-metastatic effects of β-1,3-D-glucan purified from Hanabiratake, Sparassis crispa. Biol Pharm Bull 32:259–263. https://doi.org/10.1248/bpb.32.259
Cooper PD, Sim RB (1984) Substances that can trigger activation of the alternative pathway of complement have anti-melanoma activity in mice. Int J Cancer 33:683–687. https://doi.org/10.1002/ijc.2910330520
Qi C, Cai Y, Gunn L et al (2011) Differential pathways regulating innate and adaptive antitumor immune responses by particulate and soluble yeast-derived β-glucans. Blood 117:6825–6836. https://doi.org/10.1182/blood-2011-02-339812
Baran J, Allendorf DJ, Hong F, Ross GD (2007) Oral beta-glucan adjuvant therapy converts nonprotective Th2 response to protective Th1 cell-mediated immune response in mammary tumor-bearing mice. Folia Histochem Cytobiol 45:107–114
Netea MG, Quintin J, van der Meer JWM (2011) Trained Immunity: a memory for innate host defense. Cell Host Microbe 9:355–361. https://doi.org/10.1016/j.chom.2011.04.006
Goodridge HS, Wolf AJ, Underhill DM (2009) β-Glucan recognition by the innate immune system. Immunol Rev 230:38–50. https://doi.org/10.1111/j.1600-065X.2009.00793.x
Quintin J, Saeed S, Martens JHA et al (2012) Candida albicans infection affords protection against reinfection via functional reprogramming of monocytes. Cell Host Microbe 12:223–232. https://doi.org/10.1016/j.chom.2012.06.006
Demir G, Klein HO, Mandel-Molinas N, Tuzuner N (2007) Beta glucan induces proliferation and activation of monocytes in peripheral blood of patients with advanced breast cancer. Int Immunopharmacol 7:113–116. https://doi.org/10.1016/j.intimp.2006.08.011
Acknowledgements
We thank Brent L. Berwin, Constance E. Brinkerhoff, Marc S. Ernstoff, John Frelinger, and Irene M. Mullins for helpful suggestions and feedback. We thank Jennifer L. Vella and Yina H. Huang for assistance with B16.F10-luciferase cells.
Funding
Matthew P. Alexander was supported, in part, through the Joanna M. Nicolay Melanoma Foundation. David W. Mullins was supported, in part, through USPHS R03 CA188418. Flow cytometry and multiplex assays were carried out in DartLab, the Immune Monitoring and Flow Cytometry Shared Resource, supported by a National Cancer Institute Cancer Center Support Grant to the Norris Cotton Cancer Center (P30CA023108-37) and an Immunology COBRE Grant (P30GM103415-15) from the National Institute of General Medical Sciences.
Author information
Authors and Affiliations
Contributions
MPA performed all in vitro and mouse model studies, participated in data and statistical analyses, and contributed to the drafting of the manuscript. SNF contributed to study design and data analysis, and participated in drafting of the manuscript. GRO generated YGP β-glucan, contributed to study design and data analysis, and participated in drafting of the manuscript. RAC maintained and provided Myeloid-HIF1α−/− mice, contributed to study design and data analysis, and participated in drafting of the manuscript. DWM participated in in vitro and mouse model studies, oversaw study design and data/statistical analyses, and contributed to the drafting of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
David W. Mullins is a senior scientific advisor and has received research grants, unrelated to the current work, from Qu Biologics (Vancouver, BC). The remaining authors declare no conflict of interest.
Animal studies
C57BL/6J (stock no. 000664) and CCR2−/− mice (stock no. 004999) mice were obtained from Jackson Laboratories (Bar Harbor, ME). CCR2.DTR mice were a gift from Dr. Tobias Hohl (Memorial Sloan Kettering Cancer Center). Myeloid-HIF1α−/− mice (LysM-Cre floxed-HIF1α exon 2) were generated and bred in house at Dartmouth by Dr. Robert A. Cramer.
Ethical approval
All animal care procedures and experimental protocols were performed in accordance with all applicable international, national, and/or institutional guidelines for the care and use of animals. All animal care procedures and experimental protocols were approved by the Institutional Animal Care and Use Committee (IACUC) of Dartmouth College (protocol 1141).
Cell line authentication
B16-F10 were newly acquired from ATCC, then expanded and cryopreserved to create individual aliquots that were used for each study. B16.F10-luciferase cells were generated from new aliqouts of B16-F10. Cells were confirmed to be free of Mycoplasma infection using the Hek Blue Mycoplasma Detection Kit from Invivogen.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Alexander, M.P., Fiering, S.N., Ostroff, G.R. et al. Beta-glucan-induced inflammatory monocytes mediate antitumor efficacy in the murine lung. Cancer Immunol Immunother 67, 1731–1742 (2018). https://doi.org/10.1007/s00262-018-2234-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00262-018-2234-9