
Abstract
The area along middle and lower reaches of the Yangtze River is one of the biggest Cu belts in China. In the present study, the metallophytes growing in four copper (Cu)-enriched sites along the Yangtze River were surveyed to get detailed information about vegetation composition and their Cu uptake characteristics. In all sampling sites, Cu concentrations of soils were high; whereas the organic matter, acidity and salinity of most soils were on normal levels. Totally 82 plant species belonging to 45 families were recorded. All the species recorded in the present study exhibited high tolerances for Cu although they differed greatly in their abilities to accumulate Cu. Except for Rumex acetosa and Phytolacca acinosa, most species were Cu-excluders and no Cu hyperaccumulator was found. The Cu translocation factors (TFs) and bioconcentration factors (BCFs) of the 12 dominant species were fairly low, indicating low concentrations of Cu were translocated to the shoots of these species. On this basis, the potential utilization of these metallophytes for phytoremediation was discussed.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Metalliferous mining is one of the biggest industries on a global scale and has brought us massive economic benefits; meanwhile, it has also led to numerous contaminated soils. The unfavorable conditions of these heavy metal contaminated sites made themselves the sources of pollution to the ground- and surface-waters as well as the soils in their vicinities (Conesa et al. 2006). Considerable efforts therefore have been made to search cost-effective and environmentally sustainable methods to control the contaminations derived from these mining wastelands (Bradshaw 1997; Wong 2003). Phytoremediation is a plant-based approach to control the heavy metal contaminations (or other pollutants), which was defined as the use of plants to remove heavy metals from the environment or to render them harmless (Salt et al. 1998; Pilon-Smits 2005). In general, phytoremediation can be largely divided into phytoextraction and phytostabilization. The aim of phytoextraction is to remove heavy metals from contaminated soils and an ideal phytoextraction depends on high-biomass hyperaccumulators (Baker et al. 1994; Brooks 1998). Meanwhile, phytostabilization is the use of plants to reduce the bioavailability of pollutants in the environment when the heavy metal contaminations were too heavy to be removed in an acceptably long time (Salt et al. 1998; Wong 2003). Therefore, the selection of suitable plants is of vital importance in the application of phytoremediation.
Metallophytes are plant species that have evolved biological mechanisms to resist, tolerate, or thrive on the toxic metalliferous soils (Whiting et al. 2004). They can exclude, indicate or accumulate toxic levels of heavy metals, which endow them with promising applications in phytoremediation (Baker 1981). Thus autochthonous plant species that naturally grow on metal-contaminated sites may represent an inexpensive long-term solution in coping with heavy metal contaminations (Conesa et al. 2006). Besides the practical applications in phytoremediation, metallophytes are also of theoretical interests because of their long period evolutions under harsh selective pressures (Bradshaw and McNeilly 1981). Evolutionary processes of metallophytes have attracted attentions as classical examples of micro-evolution (Macnair 1993). It was suggested that the populations of metallophytes growing on contaminated soils were genetically distinct from those thriving on uncontaminated sites nearby (Macnair et al. 2000). Therefore, metallophytes provided ideal materials to study the evolutionary processes of organism under heavy metal pressures. However, because of the rapid development of the mining industry and urbanization, which is especially the case in developing countries such as China (Li 2006), many habitats of metallophytes have been destroyed and considerable metallophytes might be extinct before being recorded. Therefore, more precise geobotanical explorations on metal-enriched soils are urgently needed (Whiting et al. 2004).
The area along middle and lower reaches of the Yangtze River is one of the biggest Cu belts in China (Chang et al. 1991). Spoil heaps and mining deposits intensively distributed in this area and several large-scale Cu mines are still in practice. Some metallophytes growing on cupriferous soils in this area have been reported in previous studies (Tang et al. 1999; Shu et al. 2001a; Lou et al. 2004; Weng et al. 2005). However, most of these studies were either focused on a few plant species or restricted in one single cupriferous site; and a comprehensive knowledge of autochthonous metallophytes and corresponding biogeochemical characters is therefore still poorly understood.
In the present study, a large scale field survey was carried out to get detailed information about the metallophytes growing on four typical Cu-enriched sites along the middle and lower reaches of the Yangtze River, and further to evaluate the potential applications of these metallophytes in phytoremediation.
Materials and methods
Site description
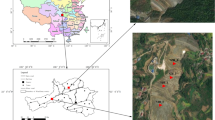
Four Cu-enriched sites were selected for investigation: Dagongshan (DGS) at Nanling county, Fenghuangshan (FHS) and Shizishan (SZS) at Tongling city, and Tongshan (TS) at Chizhou city. All of these sites were located in Anhui Province along the middle and lower reaches of the Yangtze River (Fig. 1). The detailed descriptions of these sites are given in Table 1.

a The map of China; b the map of Anhui Province, showing the Yangtze River and four sampling sites (1 DGS; 2 SZS; 3 FHS; 4 CZ)
Sampling
Sampling was carried out during May–June 2006. Samples of soils and plants were collected from 44 separate 1 × 1 m plots as well as three separate 5 × 5 m plots, which were established based on vegetation, topography and slope at each site. All vascular plants growing on each plot were recorded. The relative abundance of each species within the plots was estimated visually and then described as dominant, occasional, or rare. Plant samples included leaves and roots for tree and bush, and shoots and roots for herbs. At least three individual plants of each plant species were randomly collected within a plot and then were mixed to give a composite plant sample. The soils associated with the rhizosphere of the sampled plants within each plot were collected and mixed to give a composite soil sample; and one soil sample (0–20 cm) was collected at each plot.
Soil analysis
The soil samples were air-dried, grounded into fine powder, and sieved through a 1 mm nylon sieve. Soil pH value and electrical conductivity (EC) were measured in a 1:2.5 (w/v) ratio of soil to distilled water using a pH meter and EC meter, respectively. Soil organic matter (OM) was determined by the method of Walkley and Black (1934). For total Cu determination, the soil samples were digested in 4 ml of ‘‘aqua-regia’’ (HNO3/HCl = 1/3, v/v; McGrath and Cunliffe 1985). The extractable Cu in soils was extracted with a diethylenetetraminepentaacetic acid (DTPA) solution (Lindsay and Norvell 1978). Cu concentrations in soil samples were determined using an atomic absorption spectrophotometer (AAS, Hitachi-Z-5300).
Plant analysis
The plant samples were carefully washed with running tap water, rinsed with deionized water, separated into shoots and roots, and then oven-dried at 70°C for 72 h. Total Cu in plant materials was extracted with a mixture of concentrated HNO3 and concentrated HClO4, at 5:1 (v/v) (Allen 1989). The concentrations of Cu in plant materials were determined by AAS (Hitachi-Z-5300).
Data analysis
All statistical analyses were performed using the statistical package SPSS 10.0 for Windows (SPSS Inc., USA). The translocation factor (TF) and bioconcentration factor (BCF) of dominant species were calculated as follows: TF = Cu concentration in plant shoots/Cu concentration in plant roots; BCF = Cu concentration in plant shoots/Cu concentration in soil (Rotkittikhun et al. 2006; McGrath and Zhao 2003). The TF and BCF values of each dominant species were the means of those calculated separately for different individual samples. The TF and BCF values of different dominant species were separately analyzed and then compared using one way ANOVA followed by the least significant difference (LSD) test with the SPSS statistical package.
Results and discussion
Physicochemical properties of the four Cu-enriched sites
The physicochemical properties of the four Cu-enriched sites are shown in Table 1. All of these sites contained high concentrations of total and DTPA-extractable Cu, and each soil sample contained at least 1,500 mg/kg total Cu. However, there were great variations in Cu concentrations among and within different sites. For example, the average Cu concentrations in SZS and DGS were 8,226 and 7,005 mg/kg, which were significantly higher than those of FHS and TS (3,090 and 4,120 mg/kg, respectively). Meanwhile, although the abandoned periods in DGS and SZS were longer than those of the other two sites, it was noticed that there were large numbers of waste rocks and slag dumps distributed in DGS and SZS, which might result in the higher levels of Cu contamination in these site (Table 1). Values of pH varied from 5.08 (Plot: TS13) to 7.39 (Plot: SZS07), and those of EC varied from 0.40 (Plot: TS01) to 2.90 (Plot: DGS11) dS/m. It was noticed that the EC values of DGS and TS were distinctly higher than those of FHS and SZS. Soil matrix could be easily dissolved under acidic conditions, and thus resulted in high salt contents (Shu et al. 2001b). Therefore, the variance in EC values might be a reflection of varied pH values in different sites. The soil OM varied from 0.71% (Plot: TS01) to 6.23% (Plot: SZS08) with an average of 2.66%. It was reported that the soil OM of typical farmlands in the Yangtze River Delta ranged from 1.38 to 1.96% in 2002 (Zhu et al. 2006), which was slighter lower than those obtained in the present study (Table 1), and thus indicated normal levels of organic matter in surveyed sites. On the other hand, soil heavy metals pollution could change the mineralization rate of soil organic matter and increase the amount of particular organic matter (Zhang and Wang 2007), which might explain the high contents of organic matter in DGS and SZS when compared with the other two sites.
Copper is an essential nutrient for plants, but excessive Cu is highly phytotoxic (Weng et al. 2005). In general, phytotoxicities have been associated with 60–125 mg/kg total soil Cu (Ross 1994). However, the Cu concentrations determined in the present study (Table 1) were much higher than the threshold value, indicating that these Cu-enriched sites were the harsh habitats for plants growth. Acidity and salinity in soil can affect plant growth directly and indirectly. Generally, plant can achieve normal growth in the pH range of 5–7 and in the EC range of 0–2 dS/m except for those tolerant species (Gemmell 1977; Shu et al. 2001b). The pH values (5.08–7.39) and EC values (0.40–2.90) of the four Cu-enriched sites were, therefore, mostly in the ranges for normal growth of plants.
The overall results of physicochemical analysis indicated that some soil properties such as acidity, salinity and organic matter were not the limiting factors for plant growth. However, the Cu concentrations in these sites were extremely high, which were distinctly phytotoxic and might be the major constraint on vegetation composition and plants growth on these Cu-enriched sites.
Vegetation composition
The four sites were mostly comprised of long-abandoned mining spoil heaps and deposits that have been naturally colonized by native plant species. Totally 82 plant species belonging to 45 families was collected, including 50 herbs, 10 shrubs, 11 climbers and 11 trees (Table 2). A total of 12 dominant plants were recorded, including three Composite species, three Polygonaceae species and six species from other six families (Table 2). During field investigation, it was noticed that several families were frequently represented on different sites. These families included Caryophyllaceae, Polygonaceae, Rosaceae, Leguminosae, Compositae and Gramineae. Meanwhile, based on the data of dominant species in each plot, several communities composed of same single dominant species or same combinations of dominant species were frequently recorded. These frequently recorded communities and their associated soil Cu concentrations are shown in Table 3.
Considerable field surveys were carried out on Cu-enriched sites outside China and that was especially the case in Africa, from which a large number of Cu tolerant plants were documented (Brooks 1998; Leteinturier et al. 1999; Malaisse et al. 1999). Although metal tolerance has been demonstrated in many families of vascular plants throughout the world (Antonovics et al. 1971), several families such as Compositae and Gramineae were frequently represented in metal-contaminated habitats and were suggested to have more tolerances to unfavorable habitats (Shu et al. 2005; Rotkittikhun et al. 2006). Metallophytes for Cu were suggested to be concentrated in the families Labiatae, Scrophulariaceae, Polygonaceae, Amaranthaceae, Leguminosae, Caryophyllaceae, Gramineae, Cyperaceae, Convolvulaceae, Compositae and Tiliaceae (Tang et al. 1999), all of which were represented in the present study except for Convolvulaceae. Meanwhile, Gramineae and Compositae exhibited the maximum species in all families growing on the four Cu-enriched sites, which further indicated their universal high tolerances and adaptations to unfavorable habitats. Several indicators that have been used by Chinese geologists for ore exploration were intensively thriving on Cu-enriched sited investigated, e.g. Elsholtzia haichowensis, Commelina communis, Silene aprica and Rumex acetosa. Additionally, three interesting phenomena were noticed: firstly, as a famous “copper-flower”, E. haichowensis could hardly be found on normal soils; secondly, when compared with those from normal soils, the individuals of C. communis in Cu-enriched sites appeared rather dwarfish with intensive hairs on back surface of leaves; thirdly, the individuals of R. acetosa usually possessed purple leaves and stems rather than green leaves or stems of those from normal soils. The appearance of distinct physiological races such as metallicolous populations is traditionally considered as the first step in speciation, and the final result of that process might be endemic metallicolous taxon (Lefèbvre and Vernet 1990). Therefore, the vegetations established on Cu-enriched sites in the present study indicated that some plant taxa, e.g. Compositae, Gramineae, Polygonaceae and several copper-flowers might possess universal affinities for Cu. Meanwhile, considering the persistent harsh Cu pressures in these sites, it seemed that some species with distinct morphological properties have evolved into new ecotypes or metalloendemics.
Cu concentrations in plants
Average Cu concentrations in plants sampled in the present study and in their associated soils are shown in Table 2. In general, a large number of plants could accumulate considerable Cu in their roots; however, the Cu concentrations in shoots were fairly low and seldom exceed 100 mg/kg. The concentrations of Cu in plants were quite different: Cu concentrations in shoots of single sample were from 1.26 mg/kg in Miscanthus sinensis from TS to 317 mg/kg in Polygonum perfoliatum from DGS; and those in roots of single sample were from 12.2 mg/kg in Smilax china in TS to 2,309 mg/kg in Pteridium aquilium var. latiusulum in DGS. As to the average values, the maximum Cu concentrations in shoot and root were obtained in Arenaria serpyllifolia (117 mg/kg) and Cyrtomium fortunei (1,410 mg/kg), respectively (Table 2).
As to dominant species, the mean Cu concentrations and the ranges (minimum–maximum) in shoots, roots and associated soils were listed in Table 4. The TF and BCF values of each dominant species were the means of those calculated separately for different individual samples as shown in Fig. 2. The Cu concentrations in most of these dominant species varied greatly, and the uptakes of Cu in shoots could hardly exceed those in roots (Table 4). The TF values were from 0.038 in Setaria viridis to 3.896 in R. acetosa. Most dominant plants have TF values of less than 1 except for R. acetosa and Phytolacca acinosa. As to BCF values, the highest value (0.053) was found in Polygonum hydropiper and the lowest (0.002) was found in Boehmeria nivea. Whereas the TF and BCF values were generally low, P. acinosa presented the best performance in both TF and BCF values.

The translocation factor (TF, a) and bioconcentration factor (BCF, b) of dominant species (mean ± SD). Numbers 1–12 indicate the dominant species. 1 Elsholtzia haichowensis; 2 Commelina communis; 3 Rumex acetosa; 4 Polygonum perfoliatum; 5 Artemisia capillaries; 6 Phytolacca acinosa; 7 Silene aprica; 8 Erigeron annuus; 9 Artemisia sylvatica; 10 Setaria viridis; 11 Boehmeria nivea; 12 Polygonum hydropiper. The error bars represent standard deviation (SD). Different letters on the error bars indicate a significant difference at P < 0.05 (LSD test)
Although Cu is essential to plants, it is required in very small amounts: 5–20 mg/kg in plant tissue (dry weight) is adequate for normal growth and over 20 mg/kg is considered toxic (Adriano 1986). Based on this, all plants except for Melia azedarach collected in the present study contained toxic levels of Cu (Table 2), which further indicated the high Cu tolerance in these plants.
The value of BCF is an index of element soil–plant transfer that may favor the understanding of the heavy metal uptake characteristics (Mingorance et al. 2007). According to BCF values, metallophytes could be classified into two types: excluders were those with BCF < 1; and accumulators were those with BCF > 1 (Baker 1981). As a special type of accumulator, some plants contain unusually large concentrations of certain elements in aerial parts on a dry mass basis and thus termed hyperaccumulators (Boyd 2004); and hyperaccumulation threshold for Cu is 1,000 mg/kg (Reeves and Baker 2000). According to these criteria, most of plant species in the present study were Cu excluders, and no Cu hyperaccumulator could be identified.
Some plant species reported here have been intensively studied before. For example, E. haichowensis was the most famous Cu flower in China (Xie and Xu 1953). The Cu tolerance and mechanism of this species were intensively reported in several studies, from which the applications for phytoremediation were discussed (Jiang et al. 2004; Lou et al. 2004; Song et al. 2004). In the present study, E. haichowensis was found dominant growing on all cupriferous sites; and thus might present as a typical Cu excluder with low Cu concentration in shoot (51.3 mg/kg, on average) and relatively high Cu content in root (481 mg/kg, on average). Additionally, C. communis and P. perfoliatum were respectively reported to exceed the threshold of Cu hyperaccumulator in two former studies in other Cu-enriched sites (Shu et al. 2001a; Liu et al. 2005). However, in the present study none of the two species could hyperaccumulate Cu, even though several of these species exhibited stronger capabilities of Cu uptake than most of the other species did. The same species that could hyperaccumulate Cu reported in previous studies might represent different ecotypes under different Cu contaminations.
Phytoremediation of heavy metal contaminated soils has gained acceptance in the last decade (Pilon-Smits 2005). Considering its advantage of being environmental friendly and cost-effective, this technique might be especially applicable to developing countries such as China. To ensure a successful phytoremediation of mine degraded soils, the selection of appropriate plant species would be of vital importance (Wong 2003). The plant species selected should not only possess the tolerance of high heavy metal levels, but also distribute in a wide range of environmental conditions (Pauwels et al. 2008). In the present study, all of the 12 dominant species listed in Tables 2 and 4 were widely distributed in China and presented high Cu tolerance. Thus these species might be employed in phytoremediation of Cu-contaminated soils, and their applications could be further divided into phytoextractraction and phytostablization. R. acetosa and P. acinosa were the only two species that possessed the characteristics of Cu accumulator (Table 2, Fig. 2). To date, no Cu hyperaccumulator could be available in China. Considering that R. acetosa and P. acinosa possessed several biological and ecological features such as high dominance, high biomass and erect growth, they might be prior choices for phytoextraction of soils slightly contaminated by Cu. Except these two species, all the other ten dominant species were Cu excluders, which widely distribute in China and have high dominances on cupriferous sites. Therefore, they could be utilized in phytostabilization of heavily Cu-contaminated soils just like the sites surveyed in the present study. Furthermore, combining using of two or more of those species listed in Table 3 might be especially effective in constructing stable vegetative covers on Cu-contaminated sites and hence decreased the risk of Cu entering surrounding environments and food chains.
Conclusions
Our results indicated that the four Cu-enriched sites along the Yangtze River of China contained extremely high Cu concentrations, which provided harsh habitats for vegetation development. However, 82 plant species were recorded and several species exhibited high dominance on the four investigated sites. Combined with the results from previous studies, some families, such as Compositae, Commelinaceae, Gramineae, Labiatae and Polygonaceae, might possess high affinities for Cu. All of the species recorded presented high Cu tolerance although no Cu hyperaccumulator could be recognized. There were 12 dominant species in these Cu-enriched sites, most of which were typical Cu-excluders except for R. acetosa and P. acinosa. In sum, the detailed information on different metallophytes thriving on Cu-enriched sites obtained in the present study might likely provided researchers more choice in selecting plants for phytoremediation of Cu-contaminated soils. Considering their wide distributions, high tolerances and dominances, the dominant species recorded in the present study might be employed in phytoremediation for Cu-contaminated soils in China, and the combining using of two or more of these species could be especially effective.
References
Adriano DC (1986) Trace elements in the terrestrial environment. Springer, New York, 533 pp
Allen SE (1989) Chemical analysis of ecological materials, 2nd edn. Blackwell Scientific Publications, Oxford. 368 pp
Antonovics J, Bradshaw AD, Turner RG (1971) Heavy metal tolerance in plants. Adv Eco Res 7:1–85
Baker AJM (1981) Accumulators and excluders-strategies in the response of plants to heavy metals. J Plant Nutr 3:643–654
Baker AJM, McGrath SP, Sidoli CMD, Reeves RD (1994) The possibility of in situ heavy metal decontamination of polluted soils using crops of metal-accumulating plants. Resour Conserv Recy 11:41–49
Boyd RS (2004) Ecology of metal hyperaccumulation. New Phytol 162:563–567
Bradshaw AD, McNeilly T (1981) Evolution and pollution. Edward Arnold Limited, London, 76 pp
Bradshaw AD (1997) Restoration of mined lands—using natural processes. Ecol Eng 8:255–269
Brooks RR (1998) Plants that hyperaccumulate heavy metals. Wallingford CAB International, New York, p 380
Chang YF, Liu XP, Wu YC (1991) The copper and ore belt in the middle and lower reaches of the Yangtze River. Geology Publishing House, Beijing, p 379 (in Chinese)
Conesa HM, Faz A, Arnaldos R (2006) Heavy metal accumulation and tolerance in plants from mine tailings of the semiarid Cartagena-La Union mining district (SE Spain). Sci Total Environ 366:1–11
Gemmell RP (1977) Colonization of industrial wasteland. Edward Arnold, London, p 75
Jiang LY, Yang XE, He ZL (2004) Growth response and phytoextraction of copper at different levels in soils by Elsholtzia splendens. Chemosphere 55:1179–1187
Lefèbvre C, Vernet P (1990) Microevolutionary processes on contaminated deposites. In: Shaw J (ed) Heavy metal tolerance in plants: evolutionary aspects. CRC, Boca Raton, FL, pp 286–297
Leteinturier B, Baker AJM, Malaisse F (1999) Early stages of natural revegetation of metalliferous mine working in South Central Africa: a preliminary survey. Biotechnol Agron Soc Environ 3(1):28–41
Li MS (2006) Ecological restoration of mineland with particular reference to the metalliferous mine wasteland in China: a review of research and practice. Sci Total Environ 357:38–53
Lindsay WL, Norvell WA (1978) Development of a DTPA test for zinc, iron, manganese, and copper. Soil Sci Soc Am J 42:421–428
Liu XH, Xue Y, Zhou DM, Wu LH, Si YB, Shen ZG, Luo YM (2005) Primary study on copper tolerant plants in mine areas. J Agro-Environ Sci 24:50–54 (in Chinese)
Lou LQ, Shen ZG, Li XD (2004) The copper tolerance mechanisms of Elsholtzia haichowensis, a plant from copper-enriched soils. Environ Exp Bot 51:111–120
Macnair MR (1993) The genetic of metal tolerance in vascular plants. New Phytol 124:541–559
Macnair MR, Tilstone GH, Smith SE (2000) The genetics of metal tolerance and accumulation in higher plants. In: Terry N, Banuelos G (eds) Phytoremediation of contaminated soil and water. Lewis Publishers, FL, pp 235–250
Malaisse F, Baker AJM, Ruelle S (1999) Diversity of plant communities and leaf heavy metal content at Luiswishi copper/cobalt mineralization, Upper Katanga, Dem. Rep. Congo. Biotechnol Agron Soc Environ 3(2):104–114
McGrath SP, Cunliffe CH (1985) A simplified method for the extraction of the metals Fe, Zn, Cu, Ni, Cd, Pb, Cr, Co and Mn from soils and sewage sludges. J Sci Food Agr 36:794–798
McGrath SP, Zhao FJ (2003) Phytoextraction of metals and metalloids from contaminated soils. Curr Opin Biotech 14:277–282
Mingorance MD, Valdé B, Rossini Oliva S (2007) Strategies of heavy metal uptake by plants growing under industrial emissions. Environ Int 33:514–520
Pilon-Smits E (2005) Phytoremediation. Annu Rev Plant Biol 56:15–39
Pauwels M, Willems G, Roosens N, Frérot H, Saumitou-Laprade P (2008) Merging methods in molecular and ecological genetics to study the adaptation of plants to anthropogenic metal-polluted sites: implications for phytoremediation. Mol Ecol 17:108–119
Reeves RD, Baker AJM (2000) Metal-accumulating plants. In: Raskin I, Ensley BD (eds) Phytoremediation of toxic metals: using plants to clean up the environment. Wiley, New York, USA, pp 193–229
Ross SM (1994) Sources and forms of potentially toxic metals in soil-plant systems. In: Ross SM (ed) Toxic metals in soil-plant systems. Wiley, Chichester, UK, pp 3–26
Rotkittikhun P, Kruatrachue M, Chaiyarat R, Ngernsansaruay C, Pokethitiyook P, Paijitprapaporn A, Baker AJM (2006) Uptake and accumulation of lead by plants from the Bo Ngam lead mine area in Thailand. Environ Pollut 144:681–688
Salt DE, Smith RD, Raskin I (1998) Phytoremediation. Annu Rev Plant Biol 49:643–668
Shu WS, Yang KY, Zhang ZQ, Yang B, Lan CY (2001a) Flora and heavy metals in dominant plants growing on an ancient copper spoil heap on Tonglushan in Hubei province, China. Chin J Appl Environ Biol 7:7–12 (in Chinese)
Shu WS, Ye ZH, Lan CY, Zhang ZQ, Wong MH (2001b) Acidification of lead/zinc mine tailings and its effect on heavy metal mobility. Environ Int 26:389–394
Shu WS, Ye ZH, Zhang ZQ, Lan CY, Wong MH (2005) Natural colonization of plants on five Lead/Zinc mine tailings in Southern China. Restor Ecol 13:49–60
Song J, Zhao FJ, Luo YM, McGrath SP, Zhang H (2004) Copper uptake by Elsholtzia splendens and Silene vulgaris and assessment of copper phytoavailability in contaminated soils. Environ Pollut 128:307–315
Tang SR, Wilke BM, Huang CY (1999) The uptake of copper by plants dominantly growing on copper mining spoils along the Yangtze River, the People’s Republic of China. Plant Soil 209:225–232
Walkley A, Black CA (1934) An examination of degradation method for determining soil organic matter: a proposed modification of the chromic acid titration method. Soil Sci 37:29–35
Weng GY, Wu LH, Wang ZQ, Luo YM, Christie P (2005) Copper uptake by four Elsholtzia ecotypes supplied with varying levels of copper in solution culture. Environ Int 31:880–884
Whiting SN, Reeves RD, Richards D et al (2004) Research priorities for conservation of metallophyte biodiversity and their potential for restoration and site remediation. Restor Ecol 12:106–116
Wong MH (2003) Ecological restoration of mine degraded soils, with emphasis on metal contaminated soils. Chemosphere 50:775–780
Xie XJ, Xu BL (1953) Elsholtzia haichowensis: an indicator plant for copper ore. Acta Geologica Sin 32:360–367 (in Chinese)
Zhang MK, Wang LP (2007) Impact of heavy metals pollution on soil organic matter accumulation. Chin J Appl Ecol 18(7):1479–1483 (in Chinese)
Zhu J, Huang B, Sun WX et al (2006) Soils, tempo-spatial variation of soil organic matter of farmland and its affecting factors in a typical area of the Yangtze River Delta Region. Soils 38(2):158–165 (in Chinese)
Acknowledgements
This work was funded by the National Science Foundation of China (No. 40471117, No. 30400053), the National ‘‘863’’Project of China (No. 2006AA06Z359) and Team Work Project of the Natural Science Foundation of Guangdong Province (No. 06202438).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Ye, M., Li, J.T., Tian, S.N. et al. Biogeochemical studies of metallophytes from four copper-enriched sites along the Yangtze River, China. Environ Geol 56, 1313–1322 (2009). https://doi.org/10.1007/s00254-008-1229-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00254-008-1229-9