
Abstract
Sophora moorcroftiana, an endemic leguminous shrub in Tibet, is found in valleys, slopes, terraces from 3,000 to 3,900 m above sea level along the middle reaches of the Yarlung Zangbo River. S. moorcroftiana is more tolerant of sand burial than other species in the valleys and is regarded as an ideal species for studying acclimatization to climatic factors. Stem basal diameter showed decline as altitude increased from 3,039 to 3,928 m in the Yarlung Zangbo River valley. Population density increased with the rise of altitude. However, seed bank density, seed germination and seed weight of S. moorcroftiana did not change with changing elevation. Plants responded to altitude variances with remarkable sensitivities. Stem basal diameter size declined along the vertical gradient between plateau and mountainside. Both in alpine and plateau areas, plants similarly adapted growth to altitude, but displayed different responses with respect to reproductive parameters, such as seed bank density, dry weight of seed, seed germination, seedling density and sapling density. Sand accretion remarkably reduced seed bank density and seedling density of the total population of S. moorcroftiana, but could promote shoot sprouting and shoot growing on the individual scale that stimulated the overall development of the population. The results suggested that this particular foredune species did not merely tolerate sand burial, but actually responded positively to sand accumulation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Phenotypic adaptation to environment fluctuation is a kind of gradual and reversible adjustment of physiology and morphology to environmental conditions (Crawford 1989). Plants and vegetation display different means of acclimatizing to environment fluctuation caused by altitude (Gutierrez et al. 1998; Hegazy et al. 1998; Kotze and O’Connor 2000). Changes in dominant plant life forms, stand characteristics, seed production, seed weight, and seed germination with altitude have been well documented (Abulfatih 1995; Ares and Fowns 1999; Barclay and Crawford 1984; Crawford 1989). Most previous research focused on alpine areas and little attention was paid to plateau areas.
Sand burial is a frequent environmental stress not only in the coast and lake dune but also in the inland dune environment (Bullard 1997; Houle 1996; Moreno-Casasola 1986; van der Valk 1974; Viles 1990). The risk of burial by sand in the dune environment is high, owing to the spatial and temporal variation in the substrate (Maun 1994). The effects of sand burial on strandline flora and foredune grass species were studied in coastal and lake environment, such as Ammophila, Agropyron psammophilum, Elymus Canadensis, Cakile edentula, Corispermum hyssopifolium (Chen and Maun 1999; Crawford 1989; Lee and Ignaciuck 1985; Maun and Lapierre 1986). As for shrub species, previous studies concentrated on the response of seedlings to sand burial; few have studied the response of mature plants. In general, the ecological and eco-physiological effects of burial on established plant cover have been poorly studied compared with the frequency of such events (Kent et al. 2001).
In the middle Tibet, especially in the middle reaches of Yarlung Zangbo River valley, fragility of the natural environment due to climatic fluctuation and shifting sands determine vegetation development (Dong et al. 1996; Liu and Zhao 2001). It is also an ideal place to study adaptations of natural ecosystems to climatic gradients because the vegetation is relatively less disturbed by human activities, and the South Asian Monsoon produces wide ranges of temperature and moisture (Li and Zhou 1998).
Sophora moorcroftiana, an endemic leguminous shrub in Tibet, which plays an important role in vegetation restoration, is found in valleys, slopes, terraces from 3,000 to 3,900 m above sea level along the middle reaches of the Yarlung Zangbo River. It exhibits stronger adaptability to sand burial than other species in the valleys and is as an ideal candidate species for studying acclimatization to climate. Although S. moorcroftiana has had much attention from researchers (Sheng 1996), particularly for its use in vegetation reestablishment programs to stabilize shifting sand (Liu et al. 1998), little information is available on the adaptability of S. moorcroftiana to elevation as well as to sand burial.
The objectives of this study were to determine: (1) the response of the dominant species S. moorcroftiana to climatic factors along the gradient of altitudes in plateau areas and its adaptation differences in altitude sequence between alpine area and plateau area; (2) the adaptation of S. moorcroftiana to sand burial in the valley area.
Methods
Study area
To reveal how S. moorcroftiana adapts to altitude, six sites situated between 29°02′ and 29°20′N, and 88°21′ and 93°02′E at an altitude ranging from 3,139 to 3,925 m a.s.l. were selected to investigate the structure characteristics. The six sites were located in Nangxian (Site 1), Gyaca County (Site 2), Zhanang County (Site 3), Qüxü County (Site 4), Xigaze City (Site 5) and Xaitongmin County (Site 6) along the Yarlung Zangbo River in the mid-stream valley (Fig. 1). All sites were on the south side of the river due to topographic condition and traffic limitations.

Location of the study sites and distribution of S. moorcroftiana in Tibet autonomous region, China. Nangxian Site 1, Gyaca County Site 2, Zhangang County Site 3, Qüxü County Site 4, Xigaze City Site 5, Xaitongmin Site 6
Because there are only three meteorological observation stations in the study region and there is no observation station in Nangxian (Site 1), Gyaca County (Site 2) and Xaitongmin County (Site 6), the precipitation and air temperature of the three sites were estimated from isohyet and isothermal based on the literature (TCSTQPCAS 1984). Site location, elevation, mean air temperature and mean precipitation is shown in Table 1.
The soils in the sites are characterized by coarse-textured sand derived by fluvial and wind deposition in terraces of the river (sand: 89.2–98.7%, silt: 12.0–6.4%, clay: 0.1–4.4%). The dominant species in the terrace is S. moorcroftiana with an average coverage of 20–40%.
The investigation in the response of S. moorcroftiana to sand burial was carried out at Jiangdang (29°20′N, 89°21′E, 3820 m a.s.l.) (Site 7) and at Xigaze (Site 5) on a terrace about 6 km wide on the south side of the Yarlung Zangbo River. The mean annual air temperature is about 6.3°C, with the coldest monthly mean temperature being −3.8°C in January and and the warmest being 14.6°C in June. The mean annual accumulated air temperature above 0°C is 2,566 (cumulative warmth measurement which roughly correlates to “total growing days” in other terminology systems.) The mean annual precipitation is 434 mm, nearly 95% of which falls from June to September. The mean annual pan-evaporation is around 2,316 mm. Prevailing wind directions are southwest to west. The mean annual wind speed ranges from 1.1 to 2.9 m/s. The threshold for sand movement, 5 m/s, is exceeded 754.2 h, 90.4% of which occurs in the dry spring and winter seasons. Gales (wind speed ≥ 17 m/s) occur 59.5 days per year, 62% of them from January to May. Because wind and dry seasons occur simultaneously in the study area, sand flow occurs from January to May when deposited material along the river are exposed due to a lowering of the river water level. The geomorphology of this region is characterized by sand dune and sand-covered terrace interspersed with terrace plain. Most of sand dunes move at a rate around 8–25 m/year (Liu and Zhao 2001). The dominant plant in the terrace and sand-covered terrace is also S. moorcroftiana, with an average coverage of about 10–25%.
Investigation methods
The six sites used to investigate S. moorcroftiana’s response to altitude were at least 20 km from towns and villages so the anthropological impact (cutting) could be minimized, about 2 km from the riverside, and set away from sand dunes so the influence of currently blowing sand could be avoided.
Investigation of growth and reproduction of S. moorcroftiana with altitude
Five 10 × 10 m plots located at least 100 m from each other were marked randomly in each of the six sites. Each plot was located at a terrace with similar landforms. At each site, latitude, longitude and elevation were recorded. Within each plot, the background population density, the density of the 2-year-old S. moorcroftiana seedlings, and the tiller density were surveyed. In addition, heights, basal diameter of individual and seed bank density were investigated. Data of individual height were based on the average of 30 individuals selected randomly, and the data of basal diameter were based on the average of 30 largest twigs from 30 individuals. In each plot, 30 cores (20 × 20 cm in size and 10 cm in depth) were chosen randomly to survey the soil seed bank density. The lowest limit of 10 cm was considered adequate since the decline in seed bank with depth is usually exponential (Leck 1989). The field investigation showed that the seed banks are within 10 cm (Liu and Zhao 2002). The soil samples were sieved through a set of 1, 0.5 and 0.2 mm meshes, and seeds were counted under a stereoscopic microscope (magnification, 10–40×). Seeds obtained from the samples appeared to be intact, but because no germination tests were performed, viability could not be presumed. More than 5,000 seeds at each site were collected to determine the weight of a 1,000 seeds after air-drying at the laboratory.
Investigation of growth and reproduction of S. moorcroftiana under different burial depths
Fifty 10 × 10 m plots were selected randomly in a terrace covered with sand accretion at Site 7 to establish the relationship between seedling density, seed bank density and the depth of sand accretion (usually less than 150 cm). The methods of seedling density and seed bank density determination were similar to the methodology outlined above. The depth of sand accretion was determined by three profiles for each plot. In addition, 60 groups consisting of three individuals per group were chosen at approximately every 50 m on a transect to investigate individual height, number of shoots, height of shoots, and depth of sand accretion. The data from each group were subjected to regress analysis.
Statistics analysis
Statistics analysis was performed using Micro-origin 5.0 software. One-way ANOVA was used to compare the mean population parameters. To detect the differences between the means of two groups of samples, two sample t-tests were used. These statistics then were used to compare the population density, individual height, current annual increment of shoot, stem basal diameter, seedling density, tiller density, seed bank density, and a thousand seed weight.
A correlation matrix of r-values was constructed to establish the interrelationships among the variables. Data on number of shoots were natural logarithm transformed before regression. Then appropriate non-liner regression analysis was established to describe the function relationship among seedling density, seed bank density and depth of sand accretion. The function relationships between depth of sand accretion and individual height, shoot height, number of shoots, respectively, were also constructed.
Results
Stand structure and growth status
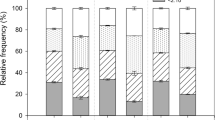
The population density increased with altitude while the stem basal diameters deceased (Table 2). The two sites with elevations of around 3,200 m, Nangxian (Site 1) and Gyaca (Site 2), had significantly lower population density and significantly greater stem basal diameters of S. moorcroftiana individuals compared with the other four sites with elevations of over 3,500 m. Differences also existed between the sites when Nangxian (Site 1) and Gyaca (Site 2) were compared with the sites in Zhanang (Site 3), Qüxü (Site 4), Xigaze (Site 5), and Xaitongmin (Site 6). Individual height had a maximum of 68.3 cm in Nangxian (Site 1) and a minimum of 22.9 cm in Xigaze (Site 5). Current annual increments of twigs showed high variation along the altitude sequence (Table 2), with a maximum of 26.3 cm in Qüxü (Site 4) and a minimum of 5.8 cm in Xigaze (Site 5).
Seed bank density, dry weight of 1,000 seeds, seedling density, and sapling density
Seed bank density, dry weight of 1,000 seeds, seedling density, and sapling density at the sites in different elevation are shown in Table 3. The seed bank density ranged from 6.4 to 12.3 grains m−2. No significant difference in seed bank density was found among the six sites. The dry weight of a 1,000 seeds fluctuated from 35.39 to 43.54 g, with the maximum of 43.54 g at Gyaca (Site 2) and the minimum of 35.39 g at Xaitongmin (Site 6). Seedling density ranging from 0.068 to 0.548 individuals m−2 differed nearly by one order of magnitude between Qüxü (Site 4) and Xigaze (Site 5).
Sapling density at Zhanang (Site 3), Qüxü (Site 4) and Xigaze (Site 5) were 0.032, 0.140, 0.086 individuals m−2, respectively, not significant differences. No sapling was found in Nangxian (Site 1), Gyaca (Site 2) and Xaitongmin (Site 6). Overall, seed bank density, dry weight of 1,000 seed, seedling density, and sapling density showed no obvious trend with altitude gradient.
Relationships between sand burial depth and seed bank, seedling density, shoot number, shoot increment and stem height
The seed bank density decreased with increasing depth of sand accretion, following a parabola function, as did seedling density. Although both decreased with depth of sand accretion, there was remarkable contrast in corresponding depth to minimum. Regression analysis indicated that the minimum seed bank density occurred at sand accretion depth of about 145 cm, while the minimum of seedling density was at around 75 cm.
The stem height, the shoot number per individual and the length of shoot changed with depth of sand accretion (Fig. 2), following the parabola function. The corresponding sand depth accretions to their maximum value were 40, 117 and 130 cm, respectively. Regression analysis indicated that individual height increased with sand accretion to an average of 39 cm at a sand depth of 40 cm, the shoot number per individual plant increased to an average of 148 at a sand depth of 117 cm, and shoot height increased to an average of 22 cm when depth was 130 cm.

Regression equation between depth of sand burial and some parameters of S. moorcroftiana. a Number of shoot. b Height of individual. c Shoot height
Discussions
The present study showed that the stem basal diameter of S. moorcroftiana individual displayed a trend of decline with increasing altitude from 3,039 to 3,928 m a.s.l. in the Yarlung Zangbo River valley (Table 2). The results are in line with previous studies on forest basal diameter (Fralish 1987). Stem density rose with increasing altitude, in agreement with the result on Nothofagus pumilio in Tierra del Fuego, Argentinano (Barrera et al. 2000). Crawford (1989) pointed out that the number of seeds per m2, dry weight per 1,000 seeds, and germination rate decreased with increasing altitude. There was a weak relationship between the stem density and stem basal diameter (r = 0.15, P > 0.1). Whether basal diameter of S. moorcroftiana could be useful in characterizing site potential needs further research.
The results showed that seed bank density had no evident trend along the gradient of elevation. Although the seed production has not been investigated, the seed bank density could be used to judge the trend of seed production along the altitude gradient. The seed production of S. moorcroftiana had no obvious trend along the gradient. However, it has been reported that the quantity of seed produced per hectare of Spruce forest varies with altitude, following the negative exponential function within an altitude gradient between 1,860 to 2,060 m a.s.l. on a Scottish mountainside (Barclay and Crawford 1984). Seed bank variation patterns in this current study are not consistent with those earlier findings in mountainous terrain. In the current study, seedling density had a fluctuation between 0.07 and 0.55 m−2, but no increase or decrease with elevation. Seedling density had a significant correlation with seed bank density (r = 0.59, P < 0.001). Seed germination of S. moorcroftiana did not increase or decrease with elevation, unlike anticipated conclusions drawn from Barley and Crawford. The seed weight also did not exhibit an obvious trend with increasing elevation.
Although stem basal diameter exhibited a similar trend with altitude in the plateau area to that on mountainside, there were remarkable differences in other population parameters between plateau and mountainside communities, such as seed bank, seedling density, and seed weight.
These differences are due to the following reasons. There is a natural difference in thermal and precipitation with altitude between the plateau area and the mountainside. The rate of mean annual temperature decline in the plateau area is similar to the mountainous area (about 0.50°C per 100 m in plateau and 0.5–0.65°C per 100 m in mountainside). Precipitation, however, decreases as altitude increases in the plateau area, but increases as altitude increases along the mountainside.
Temperature can act upon plants in a variety of ways depending on the particular species. S. moorcroftiana showed significant correlation between mean annual temperature and basal diameter (r = 0.78, P < 0.05). There were significant correlations between mean annual precipitation and individual height (r = 0.77, P < 0.05), and basal diameter (r = 0.90, P < 0.01). Temperature and precipitation are the main factors acting upon plant growth status in this study.
Responses of seed production, seedling density and sapling density to altitude in the plateau area are different from those on the mountainside because seed production and seed germination were controlled by localized micro-environment conditions within the parameters of the plot design.
Overall, plants in both alpine and plateau areas had similar means to adapt to altitude with respect to growth. Results from plateau areas coincided with the hypothesis that changes in growth form, forest structure and physiognomy were an adaptation to increasing elevation in alpine areas (Mattheck 1991). However, this species displayed different ways to respond to elevation changes between alpine and plateau areas with respect to reproductive parameters, such as seed bank density, dry weight of seed, seed germination, seedling density and sapling density.
Dunes are generally accepted as a unique and extreme habitat deserving special consideration. The main challenges to plant life in this habitat involve sand movement-accretion over aboveground parts and sand removal exposing roots. Plant response to sand burial is species-specific and also depends on the magnitude and frequency of sand burial. The total germination of strandline species decreased with increasing depth of sand burial (Chen and Maun 1999; Lee and Ignaciuck 1985; Maun and Lapierre 1986). Ammophila species responsible for the greatest fixation of sand in the yellow dune systems required a fresh deposition of sand for continuous vigorous growth (Crawford 1989). With respect to effects of sand burial on S. moorcroftiana, sand accretion reduced seed bank density and seedling density at population scale; however, it could stimulate shoot sprouting and shoot growth in individual plants. Sand burial stimulated S. moorcroftiana’s growth, consistent with the results obtained from by Crawford (1989) when studying Ammophila.
Seed bank density is reduced by burial under the wind-driven sand and the scouring effect of the airborne particles. Flowering usually occurs during the windy season and flower buds are easily damaged by the sand blasts. Mature aerial shoots on which the flower buds develop (usually shoots three years old or older) are buried by sand to some extent, even if the burial is relatively infrequent and less severe in the valley areas.
The sand burial stimulates the shoot sprout and growth under certain maximum depths of sand accretion. The texture of the sand creates higher porosity and improved water availability in comparison with habitats without sand burial. Photosynthetic capacity is improved. Sand burial can stimulate growth of S. moorcroftiana (Zhang 1996). The current finding that sand burial stimulates S. moorcroftiana’s growth in plateau and mountainous areas is consistent with the findings of Maze and Whalley (1992a, b) that foredune species such as the members of genus of Ammophila do not merely tolerate sand burial, but respond positively to sand accumulation.
References
Abulfatih HA (1995) Seed germination in Acacia species and their relation to altitude gradient in south-western Saudi Arabia. J Arid Environ 31:171–178
Ares A, Fowns JH (1999) Water supply regulates structure, productivity, and water use efficiency of Acacia koa forest in Hawaii. Oecologia 121:458–466
Barclay AM, Crawford RMM (1984) Seedling emergence in the roman Sorbus aucuparia from an altitudinal gradient. J Ecol 72:627–636
Barrera MD, Frangi JL, Richter LL (2000) Structural and functional changes in Nothofagus pumilio forests along an altitudinal gradient in Tierra del Fuego, Argentina. J Veg Sci 11:179–188
Bullard JE (1997) Vegetation and dryland geomorphology. In: Thomas DSG (ed) Arid zone geomorphology-process, form and change in dry lands. Wiley, Chichester, pp 109–131
Chen H, Maun MA (1999) Effects of sand burial depth on seed germination and seedling emergence of Cirsium picheri. Plant Ecol 140:53–60
Crawford RMM (1989) Study in plant survival. Blackwell, London
Dong GR, Dong YX, Li S, Liu YZ (1996) Study on the planning of land desertification control in the middle “Yarlung Zangbo River and its two tributaries basin in Tibet. Environment Science Press, China
Fralish JS (1987) Forest stand basal area and its relationships to individual soil and topographic factors in the Shawnee Hills. Trans Ill Acad Sci 80:183–194
Gutierrez JR, Lopez-Cortes F, Marquet PA (1998) Vegetation in an altitude gradient along the Rio Loa in the Atacama Desert of northern Chile. J Arid Environ 40:383–399
Hegazy AK, EI-Demerdash MA, Hosni HA (1998) Vegetation, species diversity and floristic relations along an altitudinal gradient in South-west Saudi Arabia. J Arid Environ 38:3–13
Houle G (1996) Environmental filters and seedling recruitment on a coastal dune in subartic Quebec (Canada). Can J Bot 74:1507–1513
Kent M, Owen NW, Dale P, Newnham RM, Giles TM (2001) Studies of vegetation burial: a focus for biogeography and biogeomorphology? Prog Phys Geog 25:455–482
Kotze DC, O’Connor TG (2000) Vegetation variation within and among palustrine wetland along an altitude gradient in KwaZulu-Natal, South Africa. Plant Ecol 146:77–96
Leck MA (1989) Wetland seed banks. In: Leck MA, Park VT, Simpson RL (eds) Ecology of soil seed banks. Academic, London
Lee JA, Ignaciuck R (1985) The physiological ecology of strandline plants. Vegetatio 62:319–326
Li WH, Zhou XM (1998) Ecosystems of Qinghai-Xizang (Tibetan) plateau and approach for their sustainable management. Guangdong Science and Technology Press, China
Liu ZM, Zhao WZ (2001) Shifting sand control in central Tibet. Ambio 30:376–380
Liu Z, Zhang J, Cheng H, Wei X (1998) Study on comprehensive rehabilitation of desertified land in the middle Basins of Yarlung Zangbo, Lhasa and Nianchu rivers in Tibet. China Environmental Sciences Press, China
Liu ZM, Zhao WZ (2002) Characteristics of the seed bank of S. moorcroftiana population in the middle reach of Yarlung Zangbo River, Tibet. Acta Ecol Sin 22:715–722
Mattheck C (1991) Trees. The mechanical design. Springer, New York
Maun MA (1994) Adaptations enhancing survival and establishment of seedling’s on coastal dune systems. Vegetatio 111:59–70
Maun MA, Lapierre J (1986) Effects of burial by sands on seed germination and seedling emergence of foredune species. Am J Bot 73:450–455
Maze KM, Whalley RDB (1992a). Effects of salt spray and sand burial on Spinifex sericeus R.Br (Poaceae). Aust J Ecol 17:9–19
Maze KM, Whalley RDB (1992b) Germination, seedling occurrence and seedling survival of Spinifex sericeus R.Br (Poaceae). Aust J Ecol 17:189–194
Moreno-casasola P (1986) Sand movement as a factor in the distribution of plant communities in a coastal sand dune system. Vegetatio 65:67–76
Sheng WS (1996) Floristic features of sand vegetation in the middle reaches of the Yarlung Zangbo, Tibet. Acta Phyto Sin 34:276–281
Team for Comprehensive Surveying of Tibet, Qinghai Plateau from Chinese Academy of Sciences (TCSTQPCAS) (1984) Climate of Tibet. Science Press, Beijing
van der Valk AG (1974) Environmental factors controlling the distribution for forbs on coastal foredune’s in Cape Hatteras national seashore. Can J Bot 52:1057–1073
Viles H (1990) ‘The agency of organic beings’: a selective review of recent work in biogeomorphology. In: Thornes JB (ed) Vegetation and erosion-processes and environments. Wiley, Chichester, pp 5–24
Zhang JH (1996) Interactive effects of soil nutrients, moisture and sand burial on the development, physiology, biomass and fitness of Cakile endentula. Ann Bot 78:591–598
Acknowledgments
This work was financially supported by project 40571026 of the National Natural Science Foundation of China. Special thanks are owed to anonymous reviewers for their fruitful suggestions to the improvement of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Zhao, W., Zhang, Z. & Li, Q. Growth and reproduction of Sophora moorcroftiana responding to altitude and sand burial in the middle Tibet. Environ Geol 53, 11–17 (2007). https://doi.org/10.1007/s00254-006-0613-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00254-006-0613-6