
Abstract
In modern industrial society, molybdenum is one of the important metals for development of the industry of rare metals. It is important to recycle the rare metals from wastes because they are technically and economically difficult to be dug and be purified, and they exist in only a few regions in the world. In this study, ModE protein derived from Escherichia coli, which is a molybdate-dependent transcriptional regulator with the ability to bind molybdate as a form of soluble molybdenum, was displayed on the yeast cell surface by α-agglutinin-based cell surface display system for the adsorption and recovery of molybdate. Displayed ModE, confirmed by immunofluorescence labeling, caught molybdate more preferably at pH 3.0 than at basic pH. Yeast cells displaying C-terminal domain of ModE, which lacks N-terminal DNA binding domain, more effectively adsorbed molybdate than those displaying full-length ModE, suggesting that the deletion of the domain unrelated to metal binding enhanced the binding ability. Our results indicated that the adsorption system on cell surface of yeast cells displaying ModE is effective not only for adsorption of molybdate as a rare metal bioadsorbent but also for the easy recovery of molybdate located on the cell surface.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
“Rare metals” are globally scarce metal elements; the digging and extraction of which from raw materials in the ground are difficult from the viewpoint of technical and economical aspects. Today, rare metals with some useful characteristics are indispensable for high-tech products, such as specific steel, lubricants, spark plugs, electronic materials, pigments, filaments of fluorescent light, and cellular phones, because they have important and prominent roles in realizing downsizing, resistance to heat, weight saving, high performance, and energy saving, although they are not directly used as raw materials. Rare metals are unevenly distributed in only a few regions, such as Africa, China, Russia, and South America. Although they are widely used as new materials of surrounding livingware and important for high-tech industry, the price of rare metals tends to fluctuate owing to their unstable supply. Especially, since autumn of 2004, the international price of these metals has greatly increased and we now face difficultly in securing rare metals (InfoMine Inc, http://www.infomine.com/). Therefore, their recycling is now necessary in modern industrial societies for the additional reasons that the metal resource itself is finite and may reach the limit of storage.
In living organisms, metals are necessary as essential elements. Among them, molybdenum is an essential trace element that is used to provide a redox active center in the molybdopterin cofactor of a number of enzymes (Rajagopalan 1996; Kisher et al. 1997). This transient metal is bioavailable in the form of water-soluble molybdate (MoO 2-4 ). In bacteria and archaea, molybdate is transported by an ABC-type transporter comprising three proteins, ModA (periplasmic binding protein), ModB (membrane protein), and ModC (ATPase; Self et al. 2001). The expression of the modABCD operon of Escherichia coli is controlled by ModE-Mo. In the absence of the high-affinity molybdate transporter, molybdate is also transported by another ABC transporter that transports sulfate/thiosulfate as well as by a nonspecific anion transporter. Thus, ModE is a bacterial transcriptional regulator (McNicholas et al. 1998; Kuper et al. 2003) that orchestrates many aspects of molybdenum metabolism by binding to specific DNA sequences in a molybdate-dependent fashion (Hollenstein et al. 2007). ModE, activated by binding molybdate, acts as both a positive and a negative transcriptional regulator of operons involved in various aspects of molybdenum metabolism. The ModE polypeptide can be subdivided into four structural domains from the N terminus to the C terminus, namely, the N-terminal domain involved in DNA binding, a linker region, and two so-called mop domains (Ulrike et al. 2000). The C-terminal domain of ModE (120–262 amino acid residues) is the molybdate-binding domain, which consists of two tandem repeats of mop domains. These β-rich mop domains found in a number of bacterial proteins have the function of binding molybdate at interdomain interfaces.
Yeast cell surface engineering (Ueda and Tanaka 2000a, b) is an effective strategy for adsorption and recovery of various metal ions on the cell surface. For example, as a tool for bioremediation, heavy metal ions such as cadmium ions are effectively adsorbed by the cell-surface-engineered yeast displaying yeast metallothionein, which is a cysteine-rich metal-binding protein (Kuroda and Ueda 2006). Furthermore, the enhanced bioadsorption of metal ions on the yeast cell surface has been established in recent years (Kuroda and Ueda 2003, 2006). These studies suggest that adsorption of metal ions by a protein displayed on the microbial cell surface has several benefits (Kuroda et al. 2001): high surface area-to-volume ratio for adsorption (Walker et al. 1989), ability to adsorb or recover metals by controlling pH and salt concentration. Adsorption on the cell surface is more advantageous than intracellular adsorption for the reason that cell-surface adsorption enables the recovery and recycling of the metal adsorbed on the cell surface without disintegration of cells, although intracellular accumulation needs extraction after disruption of cells. This system would enable us not only to remove toxic heavy metals but also to adsorb and recover useful metals such as rare metals from the environment.
In this study, we attempted to construct the cell-surface-engineered yeast with molybdate-binding ability as a recycle system for rare metals by displaying the molybdate-binding protein (ModE) on the yeast cell surface. The adsorption and recovery of MoO 2-4 by ModE-displaying yeast were evaluated, and the usability of cell-surface adsorption in the collection of rare metals is discussed.
Materials and methods
Strains and media
E. coli DH5α [F−, ϕ80dlacZ∆M15, ∆ (lacZYA-argF)U169, hsdR17 (r− K , m+ K ), recA1, endA1, deoR, thi-1, supE44, gyrA96, relA1, λ−] was used as a host cell for recombinant DNA manipulation. Saccharomyces cerevisiae Δsed1 mutant strain derived from BY4741 (MATa, his3, leu2, met15, ura3) was used as the host for construction of the cell surface display of proteins (Kuroda et al. 2009). E. coli was grown in Luria–Bertani medium [1% (w/v) tryptone, 0.5% (w/v) yeast extract, 0.5% (w/v) sodium chloride] containing 100 μg ml−1 ampicillin. The yeast was precultivated aerobically in synthetic dextrose (SD) medium [0.67% (w/v) yeast nitrogen base without amino acids (Difco, Detroit, USA), 2% (w/v) glucose, 0.5% (w/v) casamino acids, 0.002% (w/v) l-histidine, 0.003% (w/v) l-leucine, and 0.003% (w/v) l-methionine] and then cultivated in the same medium.
Construction of plasmids for cell-surface display
ModE can be subdivided into four structural domains, from the N terminus to the C terminus, as described previously (Hall et al. 1999). The regions encoding the full-length ModE (263 amino acids) and C-terminal 154 amino acids of ModE were amplified from E. coli DH5α genomic DNA by PCR using two oligonucleotides: ModE-forward or C-terminal-forward and ModE-reverse (Table 1), respectively. The amplified products, whose sequences were confirmed by the dideoxynucleotide chain termination method (Sanger et al. 1977), were digested with BglII and XhoI, and then inserted into the pULD1 vector (Kuroda et al. 2009) that enhanced the efficiency of displaying proteins on the cell surface. The constructed plasmids were named pULD1-ModE and pULD1-ModE-Ct, respectively. As a control, the pULD1-s vector for displaying only strep-tag was used (Kuroda et al. 2009).
Transformation of S. cerevisiae
The lithium acetate method (Ito et al. 1983) was used for transformation of S. cerevisiae cells. The transformants were isolated by incubation on a plate of selective SD medium [0.002% (w/v) l-histidine, 0.003% (w/v) l-leucine, and 0.003% (w/v) l-methionine] containing 0.5% (w/v) casamino acids at 30°C for 48 h.
Immunofluorescence microscopy
To confirm the display of full-length ModE and C-terminal domain of ModE on the yeast cell surface, immunofluorescence labeling of cells was performed using the FLAG epitope tag as described below. Cells were fixed with 3.7% (v/v) formaldehyde/phosphate-buffered saline (PBS; pH 7.4) for 1.5 h and incubated in PBS (pH 7.4) containing 1% (w/v) bovine serum albumin for 30 min prior to immunostaining. Mouse monoclonal anti-FLAG M2 antibody (Sigma Chemical Co., St. Louis, MO, USA) was used as the primary antibody at a dilution rate of 1:300. A mixture of cells and the antibody was incubated using a rotator for 1.5 h at room temperature. The cells were then washed with PBS (pH 7.4). Alexa Fluor 488-conjugated goat anti-mouse IgG antibody (Molecular Probes Inc., OR, USA) diluted at 1:300 was then reacted with the cells using a rotator for 1.5 h at room temperature. After washing with PBS (pH 7.4), the cells were suspended in 30 μl of PBS (pH 7.4) and observed by microscopy. Fluorescence was detected using an inverted microscope IX71 (Olympus, Tokyo, Japan) through a U-MNIBA2 mirror unit with a BP470-490 excitation filter, DM505 dichroic mirror, and a BA510-550 emission filter (Olympus). Live images were obtained using Aqua-Cosmos 2.0 software (Hamamatsu Photonics, Shizuoka, Japan) controlling a digital charge-coupled device camera (C4742-95-12ER, Hamamatsu Photonics).
Measurement of fluorescence intensity
To quantify the amount of ModE and C-terminal domain of ModE displayed on the yeast cell surface, fluorescence intensity was evaluated. After immunofluorescence labeling, the cells were precipitated by centrifugation at 800 × g and the concentration of yeast was adjusted to an OD600 of 3.0 with PBS (pH 7.4). The fluorescence intensity was measured using Fluoroskan Ascent FL (Labsystems, Helsinki, Finland) in a tissue-culture plate (353072 Multiwell 96-well; Becton-Dickinson Labware, Franklin Lakes, NJ, USA). These values of fluorescence intensity correspond to the amount of ModE or C-terminal domain of ModE displayed on the yeast cell surface.
Bioadsorption and recovery of molybdate by cell-surface-engineered yeast cells
Prior to adsorption, the cell-surface-engineered yeasts (Δsed1 mutant strain harboring pULD1-ModE or pULD1-ModE-Ct, and pULD1-s for control) were precultivated and cultivated at 30°C in SD medium with appropriate amino acids for 48 h. The cells were collected by centrifugation at 800 × g for 5 min and then washed with 10 ml of sterile water. The cells were incubated to an OD600 of 20 in 10 ml of Na2MoO4 solution containing 100 μM molybdenum using a rotator for 2 h at 30°C and centrifuged at 800 × g. The amount of residual molybdenum in the supernatant was measured using Miracle Kit (Katayama Chemical, Osaka, Japan) and by inductively coupled plasma mass spectrometry (ICP-MS; Agilent 7500cx, Agilent Technologies, Tokyo, Japan). Adsorption of copper ion was also performed as described above, although copper ion adsorption was not detected. The colorimetric method to measure molybdenum concentration using Miracle Kit was performed as follows. The supernatant (5 ml) obtained after adsorption was transferred into a conical flask and diluted with 5 ml of ion-exchanged water. The solution was boiled for 5 min after adding 1 g of ammonium persulfate. The boiled solution was diluted up to 30 ml by adding ion-exchanged water and cooled to room temperature. Then, 2 ml of the solution was transferred into a test tube, and 1 ml of 20% (v/v) hydrochloric acid and 1 ml of coloring solution [0.1% (w/v) toluene-3, 4-dithiol solution dissolved in the solution containing 4.85% (w/v) NaOH, 0.995% (v/v) ethanol, and 2% (v/v) thioglycolic acid] were added into the test tube. The mixture was intensively shaken and incubated at room temperature for the formation of the dithiol-molybdenum complex, which was observed as a change into a green color. To extract the colorimetric complex, 2 ml of chloroform was finally added and the mixture was shaken intensively. Residual molybdate was measured by the degree of the green color (A565). The obtained cell pellet was treated with PBS (pH 7.4) or 50 mM piperazine-N,N′-bis (2-ethanesulfonic acid) (PIPES) buffer (pH 7.5) or 50 mM N-tris (hydroxymethyl) methyl-3-aminopropanesulfonic acid (TAPS) buffer (pH 9.0) containing 5 mg/ml papain (Wako Pure Chemical Industries, Ltd., Osaka, Japan) to recover cell-surface-bound MoO 2−4 . A mixture of cells and papain was incubated at 30°C with gentle shaking using a rotator for 2 h. The resulting concentration of MoO 2−4 released into the buffer was also measured using Miracle Kit and by ICP-MS. Adsorption and recovery efficiencies were calculated as follows.
- Ci:
-
initial metal concentration
- Cs:
-
metal concentration of supernatant after bioadsorption experiment
- Cr:
-
metal concentration after recovery experiment
Results
Construction of plasmids for cell-surface display of ModE and C-terminal domain of ModE
The expression-enhanced plasmids pULD1-ModE and pULD1-ModE-Ct were constructed for cell-surface display of ModE and the C-terminal domain of ModE by fusion with α-agglutinin and FLAG tag for the confirmation of the display, as described in “Materials and methods” (Fig. 1). The constructed multicopy plasmids have the fusion gene composed of the glyceraldehydes-3-phosphate dehydrogenase promoter, glucoamylase signal sequence, full-length ModE or C-terminal domain of ModE, FLAG-tag, linker, and 3′-half of α-agglutinin as the cell-wall anchoring domain. As a control, the multicopy plasmid pULD1-s for display of strep-tag only was used in this study (Kuroda et al. 2009).

Plasmid pULD1-ModE or -ModE-Ct constructed for display of ModE on yeast cell surface. GAPDH glyceraldehyde-3-phosphate dehydrogenase, GA s.s. glucoamylase signal sequence. The FLAG-encoding gene was fused for detection of the successful display of ModE on the yeast cell surface. Other markers were described previously (Kuroda et al. 2009)
Cell-surface display of ModE and C-terminal domain of ModE
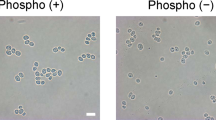
To confirm whether the ModE protein was displayed on the cell surface, FLAG tag fused with ModE or C-terminal domain of ModE was labeled by immunofluorescence. The transformants grown to the stationary phase were labeled using anti-FLAG antibody as the primary antibody and Alexa Flour 488-conjugated goat anti-mouse IgG antibody as the secondary antibody (Fig. 2). Cells harboring pULD1-ModE or pULD1-ModE-Ct for display of ModE fused with FLAG-tag showed green fluorescence on their surface. In contrast, cells harboring pULD1-s showed no fluorescence. The results of immunofluorescence labeling indicated that two types of ModE were displayed correctly on the yeast cell surface. Furthermore, to evaluate the amount of ModE or C-terminal domain of ModE displayed on the yeast cell surface, fluorescence intensity was measured after immunofluorescence labeling, as described in “Materials and methods”. Fluorescence intensity was obtained by subtracting the fluorescence intensity of the negative control BY4741 (ΔSED1) strain harboring pULD1-s. As a result, C-terminal domain of ModE was displayed more effectively than ModE on the yeast cell surface (Fig. 3).

Immunofluorescence labeling of yeast cells using anti-FLAG antibody and Alexa488-conjugated goat anti-mouse IgG antibody. Phase contract micrographs (a, c, e) and immunofluorescence micrographs (b, d, f) of S. cerevisiae harboring pULD1-ModE for display of full-length ModE (a, b); pULD1-ModE-Ct for display of C-terminal domain of ModE (c, d); pULD1-s for negative control (e, f)

Immnofluorescence intensity of full-length ModE or C-terminal domain of ModE-displaying yeast cells. After the cells were incubated in the medium at 30°C for 48 h, fluorescence intensity was measured by immunefluorescence labeling as described in the “Materials and methods”. These data are the averages ± standard deviations of three independent data
Bioadsorption and recovery of molybdate by cell-surface-engineered yeasts
The adsorption of molybdate from metal-containing water (100 μM Na2MoO4) was confirmed to observe the effect of ModE on the cell surface, as described in “Materials and methods”. As a result of colorimetric assay, lighter green indicating residual molybdate was observed after adsorption by yeast cells displaying full-length ModE and C-terminal domain of ModE, although the molybdate solution prior to adsorption showed dark green (Fig. 4a). Results shown in Fig. 4a indicated that the yeast cells displaying ModE on the cell surface could adsorb molybdate. The enhancement of molybdate adsorption indicated that displayed full-length ModE and C-terminal domain of ModE have functional ability to adsorb molybdate on the cell surface. Moreover, the adsorption ability of cell-surface-engineered yeasts at various pHs was measured by ICP-MS (Fig. 4b). The results indicated that different pH affects the molybdate-adsorption efficiency of ModE. The adsorption efficiency of C-terminal domain of ModE was little affected by pH. However, full-length ModE showed no adsorption ability at pH 13. The bioadsorption percentage of molybdate from 100 μM Na2MoO4 solution was evaluated by the formula described in “Materials and methods” (Table 2). After the adsorption, yeast cells were treated with papain to recover molybdate adsorbed on the cell surface. Molybdate released from the cells was quantified by ICP-MS and the recovery percentage, that is, the ratio of the amount of released molybdate to that of adsorbed molybdate was evaluated by the formula described in “Materials and methods” (Fig. 5). The amount of molybdate recovered from cell surface by papain was 52% of the amounts of molybdate adsorbed on C-terminal domain of ModE-displaying yeast in the case of using PBS (pH 7.4).

Cell-surface adsorption of molybdate by cell-surface-engineered yeast harboring pULD1-s (control), pULD1-ModE (Full-length display), and pULD1-ModE-Ct (C-terminal domain display). a The result of colorimetric assay of A 100 μM molybdate solution, B residual molybdate after adsorption by yeast cells displaying C-terminal domain of ModE, and C residual molybdate after adsorption by yeast cells displaying full-length ModE. b The number of molybdate ions adsorbed on a single cell was calculated by dividing into the number of adsorbed molybdate ions by the number of yeast cells. These data are the averages ± standard deviations of three independent data

Recovery of molybdate by papain dissolved in various buffers. In water, PBS (pH 7.4), 50 mM PIPES buffer (pH 7.5), and 50 mM TAPS buffer (pH 9.0), the enzyme reaction for recovery was performed using yeast cells harboring pULD1-ModE-Ct after adsorption. These data are the averages ± standard deviations of three independent data
Discussion
Surface-engineered yeast cells can be repeatedly used as bioadsorbents because the cells themselves are not damaged by metal ions in the environment for the reason that adsorbed metal ions are easily released from the cell surface. This recycle is important for the sustainability of natural resources in modern societies. The metal-adsorption ability of surface-engineered yeasts depends on the feature of the displayed metal-binding proteins. We focused on ModE as a molybdate-binding protein and attempted to display ModE on the yeast cell surface by cell surface engineering.
As shown in Fig. 4b, ModE displayed on the cell surface was effective for adsorption of molybdate. Furthermore, the displayed ModE did not show specific binding to other metals except for tungstate, an analog of molybdate. The results suggested that C-terminal domain of ModE is more effective than full-length ModE for adsorption. From the measurement of fluorescence intensity, the amount of C-terminal domain of ModE displayed on the yeast cell surface was about four times that of full-length ModE. This result indicates that the transport and targeting of full-length ModE on the cell surface may become less efficient than those of C-terminal domain during the expression of fusion genes. Western blot analysis of the cell-surface protein showed that the intensity of a single band of displayed full-length ModE was lower than that of the C-terminal domain of ModE (data not shown). The full-length ModE includes a DNA-binding domain, a linker domain, and a molybdate-binding domain. C-terminal domain includes a part of the linker domain and a molybdate-binding domain. Both types have a stretched three-dimensional structure, so that the structure formation including folding does not seem to affect display efficiency. A difference in the amount of displayed proteins between full-length ModE and C-terminal domain of ModE due to the efficiency of synthesis might affect adsorption ability. Further, C-terminal domain of ModE has the higher adsorption ability than full-length ModE. In full-length ModE, the accessibility of the molybdate-binding domain might be affected by N-terminal DNA-binding domain, because N terminus of displayed proteins is directed toward the outside of cells in α-agglutinin-based yeast cell surface display system. However, the molybdate-binding domain is exposed directly to molybdate ions in C-terminal domain of ModE displaying because of having only molybdate-binding domain. Therefore, it may be easier for cell-surface-engineered yeast cells to bind to molybdate ions. In full-length ModE, the ability of adsorption at pH 13.0 becomes lower than that at pH 7.0 (Fig. 4b). However, in C-terminal domain of ModE, the dependence of adsorption ability on pH was not found. This may explain why the structural change of a DNA-binding domain in full-length ModE has an influence on the ability of adsorption.
For recovery of the adsorbed molybdate from yeast cells, the cells were washed with solutions of various pHs (Kuroda and Ueda 2003, 2006), detergents, and denaturing agents, but no enhanced recovery of molybdate was observed. The recovery experiment was next performed by treating the molybdate-bound yeast cells with papain dissolved in various buffers [water, PBS (pH 7.4), 50 mM PIPES buffer (pH 7.5), and 50 mM TAPS buffer (pH 9.0)] (Fig. 5). The recovery efficiency was highest in use of PBS (pH 7.4). For the reuse of cell-surface-engineered yeast cells harboring pULD1-ModE-Ct and readsorption of molybdate, yeast cells treated with papain were recultivated. Recultivated yeast cells grew as well as cells before papain treatment, and the readsorption efficiency was comparable to initial adsorption efficiency (data not shown). This finding indicated that cell-surface-engineered yeast cells are available for recultivation as a novel bioadsorbent (totally, so-called “recycling”).
In this study, our results suggested that cell-surface-engineered yeast cells displaying ModE are effective for molybdate adsorption, which is the oxyanion form in environmental water. Our adsorption technique is important in modern industrial societies because molybdenum is a rare metal utilized in various fields such as those requiring new materials required for high-tech devices, including electronic materials. The design of cell-surface adsorption by yeast cell surface engineering would be advantageous for adsorption and recovery of various rare metal ions because metal-binding proteins are effectively displayed on the cell surface and can contribute to the effective recycling of rare metal ions from metal-containing water in many industrial processes.
References
Hall DR, Gourley DG, Leonard GA, Duke EM, Anderson LA, Boxer DH, Hunter WN (1999) The high-resolution crystal structure of the molybdate-dependent transcriptional regulator (ModE) from Escherichia coli: a novel combination of domain folds. EMBO J 18:1435–1446
Hollenstein K, Frei DC, Locher KP (2007) Structure of an ABC transporter in complex with its binding protein. Nature 446:213–216
Ito H, Fukuda Y, Murata K, Kimura A (1983) Transformation of intact yeast cells treated with alkali cations. J Bacteriol 153:163–168
Kisher C, Schindelin H, Rees DC (1997) Molybdenum-cofactor-containing enzymes: structure and mechanism. Annu Rev Biochem 66:233–267
Kuper J, Meyer zu Berstenhorst S, Vödisch B, Mendel RR, Schwarz G, Boxer DH (2003) In vivo detection of molybdate-binding proteins using a competition assay with ModE in Escherichia coli. FEMS Microbiol Lett 154:187–193
Kuroda K, Ueda M (2003) Bioadsorption of cadmium ion by cell surface-engineered yeasts displaying metallothionein and hexa-His. Appl Microbiol Biotechnol 63:182–186
Kuroda K, Ueda M (2006) Effective display of metallothionein tandem repeats on the bioadsorption of cadmium ion. Appl Microbiol Biotechnol 70:458–463
Kuroda K, Shibasaki K, Ueda M, Tanaka A (2001) Cell surface-engineered yeast displaying a histidine oligopeptide (hexa-His) has enhanced adsorption of and tolerance to heavy metal ions. Appl Microbiol Biotechnol 57:697–701
Kuroda K, Matsui K, Higuchi S, Kotaka A, Sahara H, Hata Y, Ueda M (2009) Enhancement of display efficiency in yeast display system by vector engineering and gene disruption. Appl Microbiol Biotechnol 82:713–719
McNicholas PM, Mazzotta MM, Rech SA, Gunsalus RJ (1998) Functional dissection of the molybdate-responsive transcription regulator, ModE, from Escherichia coli. J Bacteriol 180:4638–4643
Rajagopalan KV (1996) Biosynthesis of the molybdenum cofactor in Escherichia coli and Salmonella: Cellular and Molecular Biology, 2nd edn. American Society of Microbiology Press, Washington DC, pp 674–679
Sanger F, Nicklen S, Coulson AR (1977) DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci U S A 74:5463–5467
Self WT, Grunden AM, Hasona A, Shanmugam KT (2001) Molybdate transport. Res Microbiol 152:311–321
Ueda M, Tanaka A (2000a) Genetic immobilization of proteins on the yeast cell surface. Biotechnol Adv 18:121–140
Ueda M, Tanaka A (2000b) Cell surface engineering of yeast: construction of arming yeast with biocatalyst. J Biosci Bioeng 90:125–136
Ulrike G, Erhard S, Christoph K (2000) Structure of the molybdate/tungstate binding protein mop from Sporomusa ovata. Structure 8:1127–1136
Walker SG, Flemming CA, Ferris FG, Beveridge TJ, Bailey GW (1989) Physicochemical interaction of Escherichia coli cell envelopes and Bacillus subtilis cell walls with two clays and ability of the composite to immobilize heavy metals from solution. Appl Environ Microbiol 55:2976–2984
Acknowledgment
This study was supported by the Institute for Fermentation, Osaka.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Nishitani, T., Shimada, M., Kuroda, K. et al. Molecular design of yeast cell surface for adsorption and recovery of molybdenum, one of rare metals. Appl Microbiol Biotechnol 86, 641–648 (2010). https://doi.org/10.1007/s00253-009-2304-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-009-2304-1