
Abstract
The aim of this study was to investigate the relationship of m6A RNA methylation to CaOX-induced renal tubular injury. Microarray analysis was performed to detect the difference in mRNA expression and m6A methylation between the injurious groups and controls. We established injurious renal tubular epithelial cell model induced by calcium oxalate crystals (CaOX), and we validated that CaOX could increase the overall m6A methylation levels. By microarray analysis, we identified 5967 differentially expressed mRNAs (2444 were up-regulated and 3523 were down-regulated in the injurious groups) and 6853 differentially methylated mRNAs (4055 were in hypermethylation and 3688 were in hypomethylation in the injurious groups). Four clusters (hyper-up, hyper-down, hypo-up and hypo-down) were further identified via conjoint analysis. Functional analysis revealed that m6A methylation played a crucial role in the development of CaOX through participating multiple processes covering inflammation, oxidative stress, apoptosis, crystal–cell adhesion. We delineated the first transcriptome-wide m6A landscape of injurious renal tubular cells in high-CaOX environment. We identified a series of mRNAs of renal tubular epithelial cells with differential expression and m6A methylation between the CaOX-treated groups and controls.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Nephrolithiasis is a common metabolic disease and its prevalence in all age groups is on the rise worldwide [1]. Calcium oxalate (CaOX) crystals, which are the main ingredient of kidney stones, can lead to intrarenal inflammatory response and oxidative stress injury, and consequentially facilitate crystal aggregation, adhesion and stone formation [2]. Alleviating CaOX-induced injury of renal tubular epithelial cells has been identified as a promising strategy for the treatment of nephrolithiasis [3, 4]. However, the pathological process of crystal–cell interaction has not been well elucidated.
N6-methyladenosine (m6A) represents the most prominent internal RNA modification in eukaryotes [5]. m6A is a dynamic reversible post-transcriptional modifying process, which is regulated by methyltransferases (writers), demethylases (erasers) and recognition proteins (readers) [6]. The m6A labels can affect the fate of methylated RNAs via regulating its splicing, nuclear export, decay and translation [7]. It has been demonstrated that aberrant m6A methylation is crucial for the development of multiple human diseases, such as cancer, cardiovascular and brain diseases [8,9,10]. But so far, little progress has been made to investigate the relationship of m6A regulation to CaOX-induced renal tubular injury and stone formation. In our study, we selected human kidney proximal tubular cells (HK-2) as models due to its major role of oxalate handling [11], and delineated the first m6A methylation landscape of injurious HK-2 cells in high-CaOX environment via microarray analysis. This study firstly uncovered the pathophysiological process of crystal–cell interaction from the perspective of epigenomics.
Methods
Cell culture and CaOX exposure
HK-2 cell line was cultured in DMEM supplemented with 10% fetal bovine serum (FBS) in an incubator at 37 °C with 5% CO2. Commercial calcium oxalate monohydrate crystals were purchased from Sigma Aldrich (C0350000, USA). The adherent cells were treated with CaOX suspension at a concentration of 100 µg/mL for 24 h, and these conditions had been proven to be effective to induce oxidative stress injury [3]. Finally, RNA was collected for microarray sequencing.
Lactate dehydrogenase (LDH) release
HK-2 cells were seed into 96-well plates (5×103 cells/well). After receiving the appropriate treatments, the medium was collected and centrifuged to remove crystals and cellular debris. We measured the absorbance of culture medium at 450 nm as the results of LDH activity. The detailed process could be consulted in the instructions of LDH Activity Assay Kit (Solarbio).
Tunel assays
HK-2 cells were seed into cell climbing films and treated after adherence as mentioned above. And then, cell films were put into 4% paraformaldehyde for cell fixation. After enhancing membrane permeability, we added TdT Enzyme reaction solution and Streptavidin-TRITC as instructed by One-step TUNEL Cell Apoptosis Detection Kit (KeyGEN). Relative cell apoptosis was assessed with fluorescence microscope.
RNA extraction and quantitative real-time PCR (qRT-PCR)
Total RNA was extracted by TRIzol™ Reagent and then reverse transcribed using the TransScript® First-Strand cDNA Synthesis SuperMix (TransGen). qPCRs were conducted on Applied Biosystems 7500 system with TransStart® Green qPCR SuperMix (TransGen). Gene expression was normalized to GAPDH.
Measurement of m6A levels
The m6A levels were measured using EpiQuik m6A RNA Methylation Quantification Kit according to the manufacturer’s instructions (Epigentek). In addition, we added the m6A Dot Blot assays to validate the reliability. The 2 μg total RNAs were spotted onto a nylon membrane, and then incubated with m6A antibody (A19841, ABclonal) at 4 °C overnight. The m6A dots were analyzed using an imaging system (Bio-Rad).
Western blot
Cells were washed with PBS and lysed with RIPA buffer. An equal amount of protein was separated by SDS-PAGE and then transferred to PVDF membrane, and blotted with related antibodies in 4 °C overnight. HRP-labeled secondary antibody for 2 h at room temperature and protein bands were revealed by enhanced chemiluminescence (ECL). Antibodies used in this study were purchased from Abcam: rabbit anti-METTL3 antibody (ab195352), rabbit anti-METTL14 antibody (ab98166), rabbit anti-ALKBH5 antibody (ab195377); Cell Signaling Technology: rabbit anti-WTAP antibody (#56,501); ABclonal: rabbit anti-FTO antibody (A1438).
m6A Immunoprecipitation
3–5 μg total RNA and m6A spike-in control mixture were added to 300 μL 1 × IP buffer containing 2 μg anti-m6A rabbit polyclonal antibody (Synaptic Systems). The reaction was incubated with head-over-tail rotation at 4 °C for 2 h. 20 μL Dynabeads™ M-280 Sheep Anti-Rabbit IgG suspension per sample was blocked with freshly prepared 0.5% BSA at 4 °C for 2 h, washed three times with 300 μL 1 × IP buffer, and resuspended in the total RNA-antibody mixture prepared above. The RNA binding to the m6A-antibody beads was carried out with head-over-tail rotation at 4 °C for 2 h. The beads were then washed three times with 500 μL 1 × IP buffer and twice with 500 μL Wash buffer. The enriched RNA was eluted with 200 μL Elution buffer at 50 °C for 1 h. The RNA was extracted by acid phenol–chloroform and ethanol precipitated.
Labeling and hybridization
The “IP” RNAs and “Sup” RNAs were added with equal amount of calibration spike-in control RNA, separately amplified and labeled with Cy3 (for “Sup”) and Cy5 (for “IP”) using Arraystar Super RNA Labeling Kit. 2.5 μg of Cy3 and Cy5 labeled cRNAs were mixed. The cRNA mixture was fragmented by adding 5 μL 10 × Blocking Agent and 1 μL of 25 × Fragmentation Buffer, heated at 60 °C for 30 min, and combined with 25 μL 2 × Hybridization buffer. 50 μL hybridization solution was dispensed into the gasket slide and assembled to the m6A-mRNA&lncRNA Epitranscriptomic Microarray slide. The slides were incubated at 65 °C for 17 h in an Agilent Hybridization Oven. The hybridized arrays were washed, fixed and scanned using an Agilent Scanner G2505C.
Statistical analysis
Agilent Feature Extraction software was used to analyze acquired array images. Raw intensities of IP (immunoprecipitated, Cy5-labeled) and Sup (supernatant, Cy3-labeled) were normalized with average of log2-scaled Spike-in RNA intensities. After Spike-in normalization, the probe signals having Present (P) or Marginal (M) QC flags in a certain proportion were retained for further “m6A quantity” analyses. “m6A quantity” was calculated for the m6A methylation amount based on the IP (Cy5-labeled) normalized intensities. Differentially m6A-methylated mRNAs based on "m6A quantity" passing fold change and statistical significance cutoffs were identified and compiled. Differentially m6A-methylated RNAs and differentially expression genes between two groups were identified by filtering with fold change > 2 and p < 0.05. And then, GO enrichment and KEGG pathway enrichment analyses of these genes were performed, respectively, to explore their molecular function. By conjoint analysis, four classifications were identified and named with methylation status (hyper or hypo)-expression status (up-down). Hierarchical clustering was performed using the R software.
Results
CaOX crystals dysregulated the expression of m6A regulators and overall m6A levels of renal tubular epithelial cells
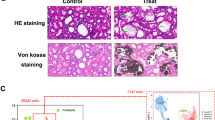
We utilized HK-2 cell line co-incubated with CaOX crystals to simulate the injurious renal tubular epithelial cells in high-CaOX environment. TUNEL and LDH release assays proved that the cell model was successfully established (Fig. 1A, B). We firstly determined whether CaOX crystals enabled to change its overall m6A levels via Dot blot and fluorometric methods. The results consistently showed that CaOX increased intracellular m6A methylation levels (Fig. 1C, D). To further explore whether m6A variations were affected by dysregulated expression of m6A regulators, we focused on three writers (METTL3, METTL14 and WTAP) and two erasers (FTO and ALKBH5), who played a major role in installing and removing m6A labels. The qPCR results showed that FTO and METTL3 were down-regulated when stimulated by CaOX crystals, and WTAP was significantly up-regulated (Fig. 1E). Western blots demonstrated that FTO, METTL14 and WTAP were all down-regulated in stimulated cells (Fig. 1F). This section indicated that the regulation of m6A methylation might be influenced by CaOX stimulation and m6A was involved in the process of crystal–cell interaction.

CaOX crystals induced renal tubular cellular injury and increased overall m6A levels. Cellular injury was measured using TUNEL assays (A) and LDH release (B). Overall m6A levels were measured using Dot Blot (C) and EpiQuik m6A RNA Methylation Quantification Kit (D). The expression of three writers (METTL3, METTL14 and WTAP) and two erasers (FTO and ALKBH5) was detected by qPCR (E) and Western Blot (F)
Differentially expressed mRNAs were identified between the injurious group and controls
Volcano plot demonstrated that a total of 5967 DEGs were screened out, of which 2444 were up-regulated and 3523 were down-regulated in the injurious groups (Fig. 2A). Hierarchical clustering and Functional enrichment analysis showed that up-regulated genes were mainly distributed in intracellular organelle and involved in cellular macromolecule metabolic process, regulation of cellular metabolic process and biosynthetic process (Fig. 3, Supplement Figure 1 and Supplement Table 1). In contrast, down-regulated genes were mainly distributed in cytoplasm and participated in cellular metabolic process, organonitrogen compound metabolic process and transport. Pathway analysis revealed that up-regulated genes were enriched in Herpes simplex virus 1 infection, Thyroid hormone signaling pathway and Ubiquitin-mediated proteolysis, while down-regulated genes were mainly enriched in ribosome, Huntington disease, NAFLD and oxidative phosphorylation (Fig. 4 and Supplement Table 2).

Differentially expressed mRNAs and methylated mRNAs were identified between the injurious group and controls. (A) Differentially expressed genes. (B) Differentially methylated genes

Heatmap and dot plots showed the results of GO analysis of differentially expressed mRNAs

Bar plots showed the enriched pathways in KEGG analysis of differentially expressed mRNAs
Differentially methylated mRNAs were identified between the injurious group and controls
To further depict RNA methylated profiling, m6A quantity, sites and percentage of total 37,015 transcripts were identified via microarray analysis. 6853 differentially methylated transcripts were selected based on the filter conditions described in Methods, among which 4055 were in hypermethylation and 3688 were in hypomethylation in the injurious groups (Fig. 2B). Surprisingly, functional analysis revealed that hypermethylated genes were also mainly distributed in intracellular organelle and involved in cellular macromolecule metabolic process and regulation of cellular metabolic process as DEGs, while hypomethylated genes were mainly distributed in cytoplasm and participated in location, organonitrogen compound metabolic process, establishment of location and transport (Fig. 5, Supplement Figure 2 and Supplement Table 3). Besides those biologic processes, we found that differentially methylated genes were also associated with cellular response to stress, cell cycle and oxidation–reduction process, which represented its preventive response to crystalline injury. Pathway analysis showed that hypermethylated genes were also mainly enriched in Herpes simplex virus 1 infection, Fanconi anemia pathway and apoptosis, while hypomethylated genes were enriched in systemic lupus erythematosus and apoptosis (Fig. 6 and Supplement Table 4).

Heatmap and dot plots showed the results of GO analysis of differentially methylated mRNAs

Bar plots showed the enriched pathways in KEGG analysis of differentially methylated mRNAs
Conjoint analysis of differentially m6A methylation and differentially gene expression
We performed a combined analysis to identify the underlying differentially expressed mRNAs regulated by m6A dynamic modification. The 4-quadrant results demonstrated that 1663 up-regulated and 20 down-regulated mRNAs were in hypermethylation status, while 674 up-regulated and 1140 down-regulated mRNAs were in hypomethylation status (Fig. 7 and Supplement Table 5). The top 10 m6A foldchange mRNAs of four classifications (hyper-up, hyper-down, hypo-up and hypo-down) are shown in Table 1. Finally, we achieved synchronous functional analysis of four classifications via Metascape. Regulation of cellular response to stress and Nervous system development were the shared Enriched Ontologies (Fig. 7 and Supplement Figure 3). Moreover, a few of annotations related to stone development were also significantly enriched, including cell junction organization, immune system development, regulation of cell adhesion, cellular response to DNA damage stimulus and cell cycle in hyper-up cluster, cytokine signaling in immune system, regulation of apoptotic signaling pathway, regulation of ion transport, cell proliferation, response to oxidative stress and signaling by NOTCH in hypo-down cluster.

Conjoint analysis identified four clusters and representative functional enrichment. (A) The 4-quadrant identified four classifications (hyper-up, hyper-down, hypo-up and hypo-down). (B) and (C) selected a subset of representative terms from the full cluster and converted them into a network layout. More specifically, each term is represented by a circle node, where its size is proportional to the number of input genes fall under that term, and its color represent its cluster identity (nodes of the same color belong to the same cluster)
Discussions
m6A RNA methylation is the most abundant mRNA modification and has emerged as a crucial gene expression regulatory mechanism related to cellular differentiation, development and cancer progression. Unlike with DNA methylation, RNA methylation takes place at post-transcriptional levels and m6A-tagged mRNAs can be recognized by “readers”, which in turn influences the fate of mRNAs on mRNA splicing, translation, stability and degradation [12]. Although RNA methylation has become the current hot topic, the relationship between m6A and urinary calculi is still a blank field. To make up for gaps, we implemented the first transcriptome-wide m6A mapping study on kidney injury induced by CaOx crystals.
Dysregulation of m6A methylation has been discovered in many pathologic processes of disease. Mathiyalagan et al. [9] reported that increased m6A levels were detected in human failing hearts, and myocardial FTO demethylase gene transfer rescued cardiac function in mouse models of myocardial infarction. Chen et al. [13] reported a negative association between m6A and autophagy in Leydig cells during testosterone synthesis and provided new insights for treating azoospermatism by exploiting m6A regulation. Niu et al. [14] found that demethylase FTO sustained the lower m6A levels of breast tumor and induced the degradation of BNIP3 (a pro-apoptosis gene), which mediated tumor growth and metastasis. In our study, we observed that intracellular overall m6A modification was up-regulated when being stimulated by CaOx crystals. And this phenomenon could only be explained by the down-regulation of demethylase FTO. This section indicated that m6A was involved in the process of crystal-cell interaction, while the key role of FTO was required to be further investigated. For other regulators, we found the unmatched results between qPCR and Western Blot, and it can be explained that the process of translation might be regulated by other mechanisms and the expression levels were also affected by different inducing conditions.
Increased intracellular generation of reactive oxygen species (ROS) can cause DNA damage and cell apoptosis [15]. High concentrations of CaOX crystals can induce oxidative stress and inflammation, and accelerate CaOX stone formation by mediating renal tubular apoptosis and crystal–cell adhesion [16, 17]. Furthermore, the functional impairment of tubular reabsorption and secretion disturbs molecular homeostasis in the tubules and promotes the supersaturation of stone-forming substances. Yu et al. [18] reported that demethylase ALKBH5 protected cells from DNA damage and apoptosis via modulating m6A modifications related to DNA repair process during ROS-induced stress. In our study, we found that m6A dysregulation had widespread influence on molecular metabolism and various cellular processes, indicating the internal complicated mechanisms of cellular response to crystalline injury. Our differentially methylated results demonstrated that herpes simplex virus 1 infection was identified as the main enrichment pathway of differential up-regulated and hypermethylated genes. Besides viral proteins, we noticed that pro- and anti-apoptotic genes were also enriched in this pathway such as Fas, Bax, Bcl-2, CASP8, BIRC2 and BIRC3, etc. Meanwhile, we found that both hypermethylated and hypomethylated genes could be enriched in apoptosis pathway. In addition to apoptosis, we also noticed that nucleotide excision repair and mismatch repair were enriched in hypermethylated genes. In the field of crystal–cell interaction, the relationship of oxidative stress to cellular injury has been most frequently investigated, and we explored the underlying mechanisms afresh at the post-transcriptional levels.
In conjoint analysis, we filtered out these genes without differential expression and differential methylation between the two omics and obtained four classifications as described above. As expected, the functional analysis indicated that m6A methylation regulated the expression of those genes associated with response to DNA damage, immune system and apoptosis. Besides these, two gene sets (cell junction and cell adhesion) should also be stressed. Cellular tight junction in the tubular cells serves as a “paracellular barrier” to limit the passage of ions and pathogens, and maintain the cell polarity and physiological function, and Peerapen et al. [19] reported that CaOX crystals could lead to tight junction disruption in distal tubular epithelial cells via p38 MAPK activation. Crystal–cell adhesion is another key point in the onset and progression of stone formation. Qi et al. [20] showed that adhesion molecules (osteopontin, hyaluronic acid and CD44) were up-regulated in CaOX-treated cells. In our study, we observed that these two gene sets (regulation of cell adhesion and cell junction organization) were also significantly enriched in three clusters, including hyper-up, hypo-up and hypo-down. These results revealed that m6A methylation exerted great impact on the development of CaOX through participating multiple processes covering inflammation, oxidative stress, apoptosis, crystal–cell adhesion, etc.
Conclusions
We delineated the first transcriptome-wide m6A landscape of injurious renal tubular cells in high CaOX environment via microarray analysis. We identified a series of mRNAs of renal tubular epithelial cells with differential expression and m6A methylation between the CaOX-treated groups and controls. In addition, we uncovered the underlying mechanisms by which m6A methylation regulated crystal-cell interaction and tubular cellular injury induced by CaOX.
References
Denburg MR, Koepsell K, Lee JJ, Gerber J, Bittinger K, Tasian GE (2020) Perturbations of the gut microbiome and metabolome in children with calcium oxalate kidney stone disease. J Am Soc Nephrol 31:1358–1369
Khan SR, Canales BK, Dominguez-Gutierrez PR (2021) Randall’s plaque and calcium oxalate stone formation: role for immunity and inflammation. Nat Rev Nephrol 17(6):417–433
Jiang K, Hu J, Luo G, Song D, Zhang P, Zhu J, Sun F (2020) miR-155–5p promotes oxalate- and calcium-induced kidney oxidative stress injury by suppressing MGP expression. Oxid Med Cell Longev 2020: 5863617
Zhu W, Zhao Z, Chou FJ, Zuo L, Liu T, Bushinsky D, Chang C, Zeng G, Yeh S (2019) The protective roles of estrogen receptor β in renal calcium oxalate crystal formation via reducing the liver oxalate biosynthesis and renal oxidative stress-mediated cell Injury Oxid Med Cell Longev 2019:5305014
Liu XM, Zhou J, Mao Y, Ji Q, Qian SB (2019) Programmable RNA N(6)-methyladenosine editing by CRISPR-Cas9 conjugates. Nat Chem Biol 15:865–871
Oerum S, Meynier V, Catala M, Tisné C (2021) A comprehensive review of m6A/m6Am RNA methyltransferase structures. Nucleic Acids Res 49:7239–7255
Zhang H, Shi X, Huang T, Zhao X, Chen W, Gu N, Zhang R (2020) Dynamic landscape and evolution of m6A methylation in human. Nucleic Acids Res 48:6251–6264
Wang X, Ma R, Zhang X, Cui L, Ding Y, Shi W, Guo C, Shi Y (2021) Crosstalk between N6-methyladenosine modification and circular RNAs: current understanding and future directions. Mol Cancer 20: 121
Mathiyalagan P, Adamiak M, Mayourian J, Sassi Y, Liang Y, Agarwal N, Jha D, Zhang S, Kohlbrenner E, Chepurko E, Chen J, Trivieri MG, Singh R, Bouchareb R, Fish K, Ishikawa K, Lebeche D, Hajjar RJ, Sahoo S (2019) FTO-Dependent N(6)-Methyladenosine Regulates Cardiac Function During Remodeling and Repair. Circulation 139:518–532
Yu J, Zhang Y, Ma H, Zeng R, Liu R, Wang P, Jin X, Zhao Y (2020) Epitranscriptomic profiling of N6-methyladenosine-related RNA methylation in rat cerebral cortex following traumatic brain injury. Mol Brain 13: 11
Wang Z, Li MX, Xu CZ, Zhang Y, Deng Q, Sun R, Hu QY, Zhang SP, Zhang JW, Liang H (2020) Comprehensive study of altered proteomic landscape in proximal renal tubular epithelial cells in response to calcium oxalate monohydrate crystals. BMC Urol 20:136
Zaccara S, Ries RJ, Jaffrey SR (2019) Reading, writing and erasing mRNA methylation. Nat Rev Mol Cell Biol 20:608–624
Chen Y, Wang J, Xu D, Xiang Z, Ding J, Yang X, Li D, Han X (2021) m(6)A mRNA methylation regulates testosterone synthesis through modulating autophagy in Leydig cells. Autophagy 17:457–475
Niu Y, Lin Z, Wan A, Chen H, Liang H, Sun L, Wang Y, Li X, Xiong XF, Wei B, Wu X, Wan G (2019) RNA N6-methyladenosine demethylase FTO promotes breast tumor progression through inhibiting BNIP3. Mol Cancer 18: 46
Yan Q, Chen Y, Liu H, Li G, Liang C, Hao Z (2022) Effects of alternative splicing events and transcriptome changes on kidney stone formation. Urolithiasis 50(2):131–140
Sun Y, Kang J, Guan X, Xu H, Wang X, Deng Y (2021) Regulation of endoplasmic reticulum stress on the damage and apoptosis of renal tubular epithelial cells induced by calcium oxalate crystals. Urolithiasis 49:291–299
Patel M, Yarlagadda V, Adedoyin O, Saini V, Assimos DG, Holmes RP, Mitchell T (2018) Oxalate induces mitochondrial dysfunction and disrupts redox homeostasis in a human monocyte derived cell line. Redox Biol 15:207–215
Yu F, Wei J, Cui X, Yu C, Ni W, Bungert J, Wu L, He C, Qian Z (2021) Post-translational modification of RNA m6A demethylase ALKBH5 regulates ROS-induced DNA damage response. Nucleic Acids Res 49:5779–5797
Peerapen P, Thongboonkerd V (2013) p38 MAPK mediates calcium oxalate crystal-induced tight junction disruption in distal renal tubular epithelial cells. Sci Rep 3: 1041
Qi S, Wang Q, Xie B, Chen Y, Zhang Z, Xu Y (2020) P38 MAPK signaling pathway mediates COM crystal-induced crystal adhesion change in rat renal tubular epithelial cells. Urolithiasis 48:9–18
Funding
This study was funded by Beijing Municipal Administration of Hospitals Clinical medicine Development of special funding support, code: 2022–2-1102; National Natural Scientific Foundation of China (Grant No. 82002711); Beijing Municipal Administration of Hospitals’ Youth Programme (Code: QML20200105).
Author information
Authors and Affiliations
Contributions
B-YY and F-ZZ conceptualized the study. F-ZZ, B-YY, JL designed the study, carried out the statistical analyses, reviewed and revised the manuscript. B-YY and F-ZZ wrote the paper. X-CW, H-MZ and YT coordinated and supervised data collection, critically reviewed and revised the manuscript. All the authors approved the final manuscript as submitted.
Corresponding author
Ethics declarations
Ethical approval
Ethical approval was not required in this study.
Conflict of interest
The authors declare that they have no conflict of interest.
Informed consent
Informed consent was not required in this study.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

{kind=link}
{kind=link}
{kind=link}
Cite this article
Yang, BY., Zhao, FZ., Li, J. et al. Epitranscriptomic profiling of m6A RNA methylation in renal epithelial tubular cells stimulated with calcium oxalate crystals through microarray analysis. Urolithiasis 51, 57 (2023). https://doi.org/10.1007/s00240-023-01425-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00240-023-01425-7