
Abstract
Most seabirds die outside the breeding season, but understanding the key factors involved is hampered by limited knowledge of nonbreeding distributions. We used miniature geolocating loggers to examine the movements between breeding seasons of Atlantic puffins Fratercula arctica from a major North Sea colony where numbers have declined in recent years, apparently due to increased overwinter mortality. The most intensively used region was the northwestern North Sea but most puffins also made excursions into the east Atlantic in the early winter. Ringing recoveries previously indicated that adults from British east coast colonies remained within the North Sea and hence were spatially segregated from those breeding on the west throughout the year. Updated analyses of ringing recoveries support results from geolocators suggesting that usage of Atlantic waters is a recent phenomenon. We propose that the increased adult mortality is related to changes in distribution during the nonbreeding period and reflects worsening conditions in the North Sea.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Our understanding of how seabirds use the marine environment during the breeding season has increased dramatically in recent years (Catry et al. 2004; Hamer et al. 2007; Phillips et al. 2008). In contrast, there is considerably less information about where birds go for the remainder of the year (but see Shaffer et al. 2006; Gonzales-Solis et al. 2007; Phillips et al. 2008; Guilford et al. 2009). This lack of knowledge is particularly critical because most mortality of seabirds occurs in the winter when food abundance is depressed, weather conditions are generally poor and sometimes extreme, and shortened day length can restrict foraging opportunities (Daunt et al. 2006). In addition, from a management perspective, there is an increasing need to identify where birds spend the nonbreeding season as a prerequisite for the designation of important areas for marine species outside the breeding season (Skov et al. 1995; Petersen et al. 2008; Phillips et al. 2008).
Until recently, most of what was known about seabirds during the winter originated from at-sea surveys and the locations of ringed birds found dead (Stone et al. 1995; Wernham et al. 2002). Both these methods have major drawbacks: interpretation of results from the former is hampered because the birds that are seen are of unknown provenance, sex, age and breeding status and cannot be followed for more than a few moments, while records from the latter are biased towards birds dying close to land or as a result of incidental mortality in fisheries or through oiling, and in areas where they are likely to be reported. These problems can potentially be overcome through the use of bird-borne loggers that record where individuals go during the period between successive breeding seasons. Satellite tags have been used to track seabirds such as penguins and albatrosses during the nonbreeding period (Nicholls et al. 2002; Putz et al. 2007; Petersen et al. 2008) but such devices have, until recently, been too heavy to be used on the smaller species that dominate the marine avifauna in northern temperate and arctic regions without potential deleterious effects (Hatch et al. 2000; Phillips et al. 2003). However, the recent development of miniature logging devices using geolocation technology (geolocators) that weigh < 2 g and can be attached to leg rings has provided the opportunity to collect location data for known individuals from smaller species over long time periods. To date, geolocators have mainly been used to track albatrosses and medium to large petrels (Shaffer et al. 2006; Gonzales-Solis et al. 2007; Guilford et al. 2009). We used the technique to determine usage of areas during the nonbreeding season by Atlantic puffins Fratercula arctica, a small (<400 g) auk, that is thought to disperse widely but which is rarely observed during at-sea surveys and for which sequential locational data on individuals breeding at British colonies are currently lacking.
The Atlantic puffin (hereafter puffin) is a pursuit-diver and one of the commonest seabirds in the northern North Atlantic, with a breeding range extending from Brittany and the Bay of Fundy northwards to the Arctic ice on both sides of the Atlantic (Lowther et al. 2002). Puffins only visit land to breed and usually return to the colonies in the North Sea in March or early April, lay in April and May, and most birds leave by the end of July. During the winter, puffins are rarely seen from land, being entirely pelagic with a range extending south to waters off Virginia, the Azores and the Canary Islands and into the western Mediterranean (Lowther et al. 2002). Analyses of ringing recoveries have suggested that different populations have different wintering areas (Petersen 1998; Harris 2002; Bakken et al. 2003), and Harris (1984) concluded that puffins from colonies on the east coast of Britain spent the winter largely within the North Sea. Although breeding populations in this area increased substantially during the latter half of the twentieth century, counts from the two largest colonies (the Isle of May and the Farne Islands) indicate that this trend has recently been reversed, with numbers declining by c.30% between 2003 and 2008 (Harris et al. 2009; Steel 2009). Survival rates of colour-ringed adults on the Isle of May (estimated using the programme MARK) over the 2006/2007 and 2007/2008 winters were markedly depressed (69 and 67%, respectively) and were well below the 95% confidence limits of the previous long-term mean at this colony (92%, CI 90–94%, n = 22 years; Harris unpublished data), strongly suggesting that reduced adult survival contributed to the population decline. Given that ringing recoveries and at-colony studies suggest most mortality occurs outside the breeding season (Wernham et al. 2002), it seems likely that conditions experienced by puffins outside the breeding season have changed in some way. To explore this possibility, we deployed geolocators on puffins at the Isle of May over the 2007/2008 winter. Here, we report on the locations used by these birds, compare the results with those from ringing recoveries and discuss our findings in the context of the recent population decline.
Materials and methods
Logger data
Geolocators (British Antarctic Survey, Mark 14) were attached to 50 adult puffins removed from breeding burrows during the chick-rearing stage at the colony on the Isle of May (56°11′N, 2°34′W), southeast Scotland between 28 June and 4 July 2007. Each device measured 20 mm× 9 mm× 5.5 mm and had a mass of 1.5 g (< 0.5% of the mean mass of a chick-rearing puffin, 390 g). The logger was attached to a coloured plastic leg ring, and a numbered metal and another coloured ring were placed on the other leg to allow individual recognition in the field. The attachment procedure took 3–5 min after which the bird was returned to its burrow. To minimise disturbance, burrows were not checked subsequently, and breeding success of the instrumented birds was therefore unknown.
Fourteen (28%) geolocators were retrieved (11 birds caught in burrows, 3 in mist-nets set in front of breeding burrows) between 4 April and 9 July 2008. Recaptured birds were sexed molecularly using DNA extracted from breast feathers (Griffiths et al. 1996). At least 11 additional birds with geolocators were seen, giving an overall minimum resighting rate of 50.0%. This value did not differ significantly from that of a control sample of colour-ringed adults checked more intensively as part of a long-term population study in another part of the colony (58.8%, n = 148, χ2 = 1.17, df = 1, P = 0.28).
Data were successfully downloaded from devices from four females and nine males. Three loggers failed part way through deployment (Bird 6514 on 20 November, Bird 6532 on 21 November and Bird 6531 on 31 December). Light data were processed following Phillips et al. (2004) in Multitrace-geolocation (Jensen Software Systems). Briefly, timing of dawn and dusk was estimated from light curves using a threshold of one, with the most appropriate angle of sun elevation (−6.0°) determined from initial analysis of a sub-sample of files, and used in the processing of all files. Latitude was derived from day length and longitude from the timing of local midday in relation to GMT and Julian day. This procedure produces two locations per day. Although use of low light thresholds partly overcomes problems of interference in the light curves at dawn or dusk, 504 locations (11.6% of those from outside the equinox periods) were later excluded for this reason and/or because they represented unrealistic movements. This resulted in a total of 3,842 validated locations (146–367 per individual). Locations were unavailable from 15 September to 26 October and from 12 February to 31 March because of the difficulties of estimating latitude around equinoxes (Wilson et al. 1992). However, longitudes were available during this period and could be used to determine whether a bird was in the Atlantic or North Sea and timing of movement. All validated locations were smoothed twice, following Phillips et al. (2004). Kernel density maps in a Lambert equal-area azimuthal (North Pole) projection were generated in ArcGIS (Hawth Tools), with a cell size of 50 km and a search radius of 200 km (Croxall et al. 2005; Phillips et al. 2005).
Although sample sizes were small, there was no suggestion of sex differences either in the likelihood of using Atlantic waters or in the timing or duration of the visit (Mann–Whitney tests, all NS) so data were pooled.
Ringing data
Information on puffins ringed on or within 100 km of the Isle of May, known to be more than 3 years old [immatures tend to disperse more widely than adults (Harris 2002)] and recovered outside the breeding season (here taken to be between August and April), was obtained from the British Trust for Ornithology. Coordinates of recovery locations were plotted to provide an overall description of puffin distribution outside the breeding season. Preliminary inspection of the data indicated a marked change in finding locations around 2000. Accordingly, recoveries were grouped into those from August 1992 to April 2000 and from August 2000 to April 2008 to give approximately similar time periods and numbers of recoveries.
For the purposes of this paper, we define the area to the west of the Northern Isles (Orkney and Shetland Islands) as the Atlantic and the area to the east as the North Sea.
Results
Wintering areas as indicated by geolocators
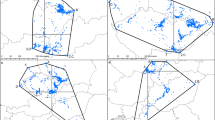
Density plots summarising overall area usage by puffins during three phases of the nonbreeding period, as defined in the following, are shown in Fig. 1. The first phase from 1 August to 31 December (excluding the period around the vernal equinox, from mid-September to late October, see “Methods”) was characterised by a widely dispersed distribution with birds recorded not only in the northern North Sea but also in the Atlantic, predominantly between 50°N and 62°N and 10°E and 15°W. In contrast, during the second phase between 1 January and 29 February, birds were much less widely dispersed, with fixes concentrated in the northwestern North Sea. Proximity to the spring equinox meant that estimated latitudes during March were unavailable, but longitudinal data were consistent with birds remaining in the same region. The third phase in April corresponded to the period when puffins had returned to the Isle of May and were spending progressively more time at the colony; birds were predominantly off northeast England, and northern and eastern Scotland, within commuting distance of the Isle of May given the puffin’s average flight speed of 70–80 km h−1 (Harris 1984). This distribution was the same as when birds were still feeding chicks and visiting the colony in July (data not shown). Apparent inclusion of the Irish Sea during all three periods and of the waters off northern and western Scotland during April in the overall range is an artefact of the kernel smoothing procedure. It is highly unlikely that birds were alternating between western Scotland and England and the northwest North Sea since puffins rarely cross land [only three inland records in Scotland between August and April 1968–2001 (Forrester et al. 2007)].

Kernel density distribution of Atlantic puffins in August–December (n = 13 birds), January–February (n = 10) and April (n = 10). Bathymetric contours are 100 m and 2,000 m. Place names referred to in the text are shown in the upper left panel
Inspection of locations of individual puffins indicated that there were two distinct between-season strategies (Fig. 2). Although 3 birds never left the North Sea, the other 10 (77%) all spent considerable periods around the north coast of Scotland or the Northern Isles, into the Atlantic. The timing and duration of the period spent in the Atlantic were highly variable (range of departures 1 August to 19 October, range of durations 27–120 days; Fig. 3). However, all eight birds whose devices were still functioning returned to the North Sea by mid-December.


Distributions of individual Atlantic puffins in August–December (red circles), January–February (blue circles) and April (green circles). Note that some devices failed part way through the period

Time periods spent in the Atlantic by 10 Atlantic puffins. Thin lines indicate periods over the equinox where the longitudinal data were inconclusive. Vertical bars indicate when devices failed. The other three birds remained within the North Sea throughout the whole nonbreeding period
The inherent error in geolocation (mean c. 185 km; Phillips et al. 2004) makes interpretation of individual short-scale movements problematic. However, it is clear that some birds, especially those entering the Atlantic, did make sustained long-distance flights. For instance Bird 6539 moved out of the North Sea on 22 August and was off the northwest coast of Scotland 24 h later (Fig. 2), between 31 October and 3 November it travelled rapidly from the Faeroe Islands to west of Ireland and later returned from near St Kilda to the northwest North Sea in about 48 h. We estimate that it travelled over 3,000 km during its excursion into the Atlantic. Other birds also visited areas around the Faeroe Islands, off western Scotland and Ireland and occasionally made similar major flights. Movements of the 3 birds that never left the North Sea were more limited.
Wintering areas as indicated by ringing recoveries
A total of 221 ringing recoveries of puffins were reported from 1992 to 2008, of which 117 were from nonbreeding periods up to April 2000 and 104 from subsequent periods. The frequency distribution of recovery months differed significantly between the two time periods, with 80% of recoveries until April 2000 occurring in January to April whereas subsequently recoveries were more dispersed with significantly fewer (58%) during this period (Fig. 4; χ2 = 34, P < 0.001, df = 1).

Monthly distributions of nonbreeding season recoveries of Atlantic puffins from August 1992 to April 2000 (n = 117) and from August 2000 to April 2008 (n = 104)
This temporal change in recovery pattern was associated with a marked distributional shift, with the proportion of recoveries coming from the Atlantic increasing significantly from 3.4% prior to 2000 to 18.3% subsequently (Fig. 5; χ2 = 13.0, P < 0.001, df = 1). This change was predominantly due to an increase in the numbers of birds reported from the Faeroe Islands (1.7% in the former period, 14.4% in the latter) and a decline in recoveries from the Bay of Biscay and the English Channel (4.4% to zero). As is normal with recoveries of ringed seabirds, the cause of death was not given (87% of 117 in the first period, 79% of 104 in the second). However, in the first period 10% were reported oiled and 2% shot whereas in the latter the equivalent values were 3 and 14%, respectively.

Ringing recoveries of adult Atlantic puffins in August–December (red circles) and January–April (blue circles) in two periods, August 1992–April 2000 (n = 117) and August 2000–April 2008 (n = 104). Place names referred to in the text are shown in the left panel
Discussion
Although we could not directly compare nonbreeding distributions of birds with and without loggers, we had no reason to suspect that the areas used by equipped birds differed significantly from those of other puffins from the Isle of May. In addition, past studies of seabirds <1.3 kg have shown no obvious deleterious effects of geolocator attachment (Rayner et al. 2008; Catry et al. 2009).
Our results using geolocators to reconstruct movements of individual puffins between successive breeding seasons indicated that the most intensively used region was the northwestern North Sea, a result which is in accordance with previous studies at this colony based on ringing recoveries (Harris 1984; Harris 2002) and general observations from at-sea surveys (Skov et al. 1995; Stone et al. 1995, S. Garthe, unpublished data, K. Kober, unpublished data, A. Webb, unpublished data)). However, the finding that more than three-quarters of the birds made a major excursion into the Atlantic was entirely unexpected and challenges the previous view that puffin populations from the east and west of Britain are spatially segregated during both the breeding season and at other times of year (Harris 2002). A study using satellite telemetry to follow post-breeding adult Atlantic puffins from a colony in northern Norway also highlighted unexpected areas with the birds moving north into the Barents Sea where there was little chance of ringed birds being reported (Anker-Nilssen and Aarvak 2009).
Comparisons of various aspects of puffin biology at east and west coast colonies during the 1970s and 1980s found marked differences, for example in the scheduling of colony attendance and pollutant levels, and in both cases differences in overwintering areas were suggested as potential explanations (Harris 1982, 1985). Thus, Harris (1985) speculated that the earlier return to east coast colonies was because ‘adults were freed from the necessity of flying several thousand miles during the winter’, and remained in the northwestern North Sea, relatively close to their breeding colonies between breeding seasons. Similarly, higher levels of polychlorinated biphenols (PCBs) but lower levels of cadmium in east coast birds compared to those from the west were thought to reflect differences in concentrations of anthropogenic and naturally occurring chemicals and heavy metals in the North Sea and east Atlantic (Bull et al. 1977; Harris 1984). In the light of the new evidence from the geolocators indicating less pronounced spatial segregation over the winter, a re-evaluation of east—west differences in phenology and pollutant levels is needed. Comparisons of adult survival rates would also be instructive. The only colony in west Britain where the survival of puffins is monitored is Skomer, southwest Wales. Previously, relationships between survival and oceanographic conditions were similar on the Isle of May and Skomer (Harris et al. 2005) but the very low survival rates on the Isle of May in recent years have not been apparent on Skomer (C.M. Perrins, unpublished data), despite the likely overlap in distributions of the two populations during the nonbreeding season. Clearly, integrated comparisons across study sites are needed to investigate this further.
Previous analyses of ringing recoveries indicated that the small minority of Isle of May birds that left the North Sea moved south into the English Channel and Bay of Biscay (Harris 1984). In contrast, our updated analysis highlights that since 2000, birds have increasingly been recovered in northern and western areas while usage of southern areas has declined. Additional support for this change comes from at-sea surveys with very few puffins being recorded in winter around the Faeroe Islands between 1979 and 2000 (Skov et al. 2002). However, hunters and local naturalists both report that many more birds now winter in the area, including many small-sized individuals typical of those that breed in Britain and the Faeroe Islands (J.-K. Jensen, unpublished data). All the recent recoveries in the Faeroe Islands are of birds that have been shot during the hunting season from 1 October to 20 January, which is a serious potential bias. However, although there was an increase in winter hunting of auks following the restriction of summer shooting in 1980, there has been no obvious further change in the overall hunting pressure in recent decades (B. Olsen, unpublished data, J.-K. Jensen, unpublished data). Taken together with our tracking data, these findings provide strong evidence for a recent major northwards shift in the distribution of puffins from eastern Britain outside the breeding season.
Ringing recovery data also suggest that the change in distribution has been accompanied by a change in the temporal pattern of overwinter mortality. Thus, prior to 2000 most recoveries occurred in the second half of the winter but recently there has been a second peak in autumn/early winter (Fig. 4). Beached Bird Survey data from the Northern Isles also suggest increased early winter mortality with the numbers recorded on beaches in autumn being particularly high in 2006 and 2007 compared with the previous 25–30 years (Heubeck et al. in press).
This accumulating evidence for a recent change in locations and mortality patterns of Isle of May puffins is of particular interest given the dramatic decrease in the breeding population recorded at this colony in 2008 and the apparent increase in adult mortality rates (Harris et al. 2009). Our results suggest that conditions in the North Sea may have become less favourable for puffins in recent years, particularly in the autumn and early winter, forcing many birds to move into the Atlantic, in some cases into areas where they are at risk of being shot. Individuals are thus likely to travel greater distances and use a wider variety of areas than previously. The inherent error in geolocation fixes means that investigating fine scale habitat usage is problematic. However, some broad differences are apparent, with areas used by puffins in the North Sea characterised by relatively shallow water typically <100 m deep while locations in the Atlantic are associated with deeper waters out to the 2,000 m isobath along the shelf break (Fig. 1). While it seems clear that changes in distribution and mortality patterns have occurred, the factor(s) causing these changes are currently unknown. Hydrobiological conditions in the North Sea during the spring and summer are known to have changed dramatically since the late 1990s, which has been implicated in recent breeding failures of seabirds (Frederiksen et al. 2004; Wanless et al. 2005). In particular, the incursion of warmer water and associated changes in the Calanus copepod community are thought to have had a negative impact on recruitment rates of lesser sandeel Ammodytes marinus, a major prey of puffins and many breeding seabirds (Arnott and Ruxton 2002; van Deurs et al. 2009). Sea temperatures in the North Sea in autumn and winter have also increased markedly, but little is known about how this affects important prey for marine birds, since in many cases (including for the puffin) almost nothing is known about the diet outside the breeding season. In contrast to the situation in the North Sea, temperatures in the Atlantic, particularly off northwest Scotland, have shown much less change (Holliday et al. 2008). Whether this difference has any relevance to changes in puffin area usage is uncertain. However, as more data from geolocators become available, combining them with information on hydrobiological conditions could help to start elucidate the mechanisms underpinning observed changes in population processes.
The overwinter period is particularly crucial for puffins because like other members of the Alcidae, birds undergo their main moult outside the breeding season. At this time, the primary wing feathers are moulted more-or-less synchronously rendering the bird flightless and thus highly susceptible to adverse conditions. We know very little about moult in wild Atlantic puffins but captive tufted puffins Lunda cirrhata are flightless for 4–5 weeks (Bridge 2004), and it seems likely that the period will be similar in Atlantic puffins. Flightless birds have been recorded in all months between October and April and assuming that birds found dead on beaches provides a robust index, moult in the 1970s appeared to peak in late winter (Harris and Yule 1977). However, recently increased numbers of birds have been found flightless in autumn. For example, 89% of a sample of 36 adults (including two ringed on the Isle of May), found dead during a wreck of in the Northern Isles between 22 October and 5 November 2007, were in wing moult (Heubeck et al. in press; Harris unpublished data). This raises the intriguing possibility that puffins are moving out of the North Sea to find suitable areas to moult. However, testing this hypothesis is problematic with the current generation of geolocators because their spatial resolution is not good enough to pinpoint periods when birds do not move. The loggers also recorded saltwater immersion, which we assumed would allow us to discriminate when a bird was on the water compared to in flight, and hence to identify long periods when the bird did not fly. To our surprise, no such periods were obvious in any of the records. However, discussions with colleagues familiar with puffins in captivity revealed that birds on the water often have either one or both legs and feet retracted into the feathers where presumably they dry out and so reduce heat loss. Hence, in comparison with recent studies of other seabirds (Phalan et al. 2007; Catry et al. 2009; Guilford et al. 2009), which presumably do not show this behaviour, the usefulness of immersion data for inferring activity patterns is limited in the puffin and in particular the method will not identify the flightless period.
Uncertainty about distributions of puffin populations when not breeding has previously hampered interpretation of studies comparing demographic parameters, particularly adult survival. A recent analysis by Grosbois et al. (2009) found evidence for synchronous changes in survival at colonies distributed across a wide latitudinal range from southern Britain to northern Norway, which was partly related to local sea surface temperature, but in which another unknown environmental factor had to be involved. Combining cross-colony survival analyses with traditional ringing methods and large scale deployment of geolocators could potentially provide a rapid and powerful way not only to identify important population level area and habitat preferences but also to evaluate the degree of migratory connectivity amongst these populations.
References
Anker-Nilssen T, Aarvak T (2009) Satellite telemetry reveals post-breeding movements of Atlantic puffins Fratercula arctica from Røst, North Norway. Polar Biol. doi 10.1007/s00300-009-0665-7
Arnott SA, Ruxton GD (2002) Sandeel recruitment in the North Sea: demographic, climatic and trophic effects. Mar Ecol Prog Ser 238:199–210
Bakken V, Runde O, Tjørve E (2003) Norsk Ringmerkingsatlas (Norwegian Ringing Atlas), vol 1. Stavanger Museum, Stavanger
Bridge ES (2004) The effects of intense wing molt on diving in alcids and potential influences on the evolution of molt patterns. J Exp Biol 207:3003–3014
Bull KR, Murton RK, Osborn D, Ward P, Cheng L (1977) High levels of cadmium in Atlantic seabirds and sea-skaters. Nature (London) 269:507–509
Catry P, Phillips RA, Phalan B, Silk JRD, Croxall JP (2004) Foraging strategies of grey-headed albatrosses Thalassarche chrysostoma: integration of movements, activity and feeding events. Mar Ecol Prog Ser 280:261–273
Catry T, Ramos JA, Le Corre M, Phillips RA (2009) Movements, at-sea distribution and behaviour of a tropical pelagic seabird: the wedge-tailed shearwater in the western Indian Ocean. Mar Ecol Prog Ser 391:231–242
Croxall JP, Silk JRD, Phillips RA, Afanasyev V, Briggs DR (2005) Global circumnavigations: tracking year-round ranges of non-breeding albatrosses. Science 307:249–250
Daunt F, Afanasyev V, Silk JRD, Wanless S (2006) Extrinsic and intrinsic determinants of winter foraging and breeding phenology in a temperate seabird. Behav Ecol Sociobiol 59:381–388
Frederiksen M, Wanless S, Harris MP, Rothery P, Wilson LJ (2004) The role of the industrial fishery and climate change in the decline of North Sea black-legged kittiwakes. J Appl Ecol 41:1129–1139
Gonzales-Solis J, Croxall JP, Oro D, Ruiz X (2007) Transequatorial migration and mixing in the wintering areas of a pelagic seabird. Front Ecol Environ 5:297–301
Griffiths R, Daan S, Dijkstra C (1996) Sex identification in birds using two CHD genes. Proc Royal Soc Lond B 263:1251–1256
Grosbois V, Harris MP, Anker-Nilssen T, McCleery RH, Shaw DN, Morgan BJT, Gimenez O (2009) Modeling survival at multi-population scales using mark-recapture data. Ecology 90:2922–2932
Guilford T, Meade J, Willis J, Phillips RA, Boyle D, Roberst S, Collett M, Freeman R, Perrins CM (2009) Migration and stopover in a small pelagic seabird, the Manx shearwater Puffinus puffinus: insights from machine learning. Proc Royal Soc B 276:1215–1223
Hamer KC, Humphreys EM, Garthe S, Hennicke J, Peters G, Phillips RA, Harris MP, Wanless S (2007) Annual variation in diets, feeding locations and foraging behaviour of gannets in the North Sea: flexibility, consistency and constraint. Mar Ecol Prog Ser 338:295–305
Harris MP (1982) The breeding seasons of British puffins. Scott Birds 12:11–17
Harris MP (1984) The Puffin. T. and A.D. Poyser, Calton, London
Harris MP (1985) Morphology and breeding of puffins at Isle of May and St Kilda. Scotl Biol Conserv 32:81–97
Harris MP (2002) Atlantic Puffin (Puffin) Fratercula arctica. In: Wernham CV, Toms M, Marchant JH, Clark JA, Siriwardena GM, Baillie SR (eds) The Migration Atlas; movements of the birds of Britain and Ireland. T. and A.D. Poyser, London, pp 407–409
Harris MP, Yule RF (1977) The moult of the puffin Fratercula arctica. Ibis 119:535–540
Harris MP, Anker-Nilssen T, McCleery RH, Erikstad KE, Shaw DN, Grosbois V (2005) Effect of wintering area and climate on the survival of adult Atlantic puffins Fratercula arctica in the eastern Atlantic. Mar Ecol Prog Ser 297:283–296
Harris MP, Newell M, Leitch A, Bruce B, Hunt J (2009) Dramatic decline in numbers of Atlantic Puffins in the Firth of Forth. Scottish Birds 29:132–134
Hatch SA, Meyers PM, Mulcahy DM, Douglas DC (2000) Performance of implantable satellite transmitters in diving seabirds. Waterbirds 23:84–94
Heubeck M, Meek ER, Mellor RM, Wilson M (2009) Numbers of Atlantic Puffins Fratercula arctica found on beached bird surveys in Orkney and Shetland over a 30-year period. Seabird (in press)
Holliday NP, Kennedy J, Kent EC, Marsh R, Hughes SL, Sherwin T, Berry DI (2008) MCCIP annual report card 2007–2008 annual review—sea temperature marine climate change impacts partnership. www.mccip.org.uk/arc
Lowther PE, Diamond AW, Kress SW, Robertson GJ, Russell K (2002) Atlantic Puffin (Fratercula arctica). In: Poole A, Gill F (eds) The birds of North America. The Birds of North America, Inc, Philadelphia, PA, pp 1–23
Nicholls DG, Robertson CJR, Prince PA, Murray MD, Walker KJ, Elliott GP (2002) Foraging niches of three Diomedea albatrosses. Mar Ecol Prog Ser 231:269–277
Petersen A (1998) Islenskir Fuglar. Vaka-Helgafell, Reykjavik
Petersen SL, Phillips RA, Ryan PG, Underhill LG (2008) Albatross overlap with fisheries in the Benguela upwelling system: implications for conservation and management. Endanger Species Res 5:117–127
Phalan B, Phillips RA, Silk JRD, Afanasyev V, Fukuda A, Fox J, Catry P, Higuchi H, Croxall JP (2007) Foraging behaviour of four albatross species by night and day. Mar Ecol Prog Ser 340:271–286
Phillips RA, Xavier JC, Croxall JP (2003) Effects of satellite transmitters on albatrosses and petrels. Auk 120:1082–1090
Phillips RA, Silk JRD, Croxall JP, Afanasyev V, Briggs DR (2004) Accuracy of geolocation estimates for flying seabirds. Mar Ecol Prog Ser 266:265–272
Phillips RA, Silk JRD, Croxall JP (2005) Foraging and provisioning strategies of the light-mantled sooty albatross at South Georgia: competition and co-existence with sympatric pelagic predators. Mar Ecol Prog Ser 285:259–270
Phillips RA, Croxall JP, Silk JRD, Briggs DR (2008) Foraging ecology of albatrosses and petrels from South Georgia: two decades of insights from tracking technologies. Aquat Conserv Mar Freshw Ecosyst 17:S6–S21
Putz K, Schiavini A, Rey AR, Luthi BH (2007) Winter migration of magellanic penguins (Spheniscus magellanicus) from the southernmost distributional range. Mar Biol 152:1227–1235
Rayner MJ, Hauber ME, Clout MN, Seldon DS, Van Dijken S, Bury S, Phillips RA (2008) Foraging ecology of the Cook’s petrel Pterodroma cookii during the austral breeding season: a comparison of its two populations. Mar Ecol Prog Ser 370:271–284
Shaffer SA, Tremblay Y, Weimerskirch H, Scott D, Thompson DR, Sagar PM, Moller H, Taylor GA, Foley DG, Block BA, Costa DP (2006) Migratory shearwaters integrate oceanic resources across the Pacific Ocean in an endless summer. Proc Natl Acad Sci USA 103:12799–12802
Skov H, Durinck J, Leopold MF, Tasker ML (1995) Important areas for seabirds in the North Sea. Bird Life International, Cambridge
Skov H, Upton A, Reid J, Webb A, Taylor S, Durinck J (2002) Dispersion and vulnerability of marine birds in Faroese waters. Joint Nature Conservation Committee, Aberdeen
Steel D (2009) Birds on the Farne Islands in 2008. Trans Nat Hist Soc Northumbr 69:53–118
Stone CJ, Webb A, Barton C, Ratcliffe N, Reed TC, Tasker ML, Camphuysen CJ, Pienkowski MW (1995) An Atlas of seabird distribution in north-west European waters. JNCC, Peterborough
van Deurs M, van Hal R, Tomczak MT, Jónasdóttir SH, Dolmer P (2009) Recruitment of lesser sandeel Ammodytes marinus in relation to density dependence and zooplankton composition. Mar Ecol Prog Ser 381:249–258
Wanless S, Harris MP, Redman P, Speakman J (2005) Low fish quality as a probable cause of a major seabird breeding failure in the North Sea. Mar Ecol Prog Ser 294:1–8
Wernham CV, Toms M, Marchant JH, Clark J, Siriwardena G, Baillie SR (2002) The migration atlas; movements of birds of Britain and Ireland. T. & A.D.Poyser, London
Wilson RP, Ducamp JJ, Rees G, Culik BM, Niekamp K (1992) Estimation of location: global coverage using light intensity. In: Priede IM, Swift SM (eds) Wildlife telemetry: remote monitoring and tracking of animals. Ellis Horward, Chichester, pp 131–134
Acknowledgments
We thank J. Fox and V. Afansyev for geolocator device development, M. Bogdanova for help with post-processing data, M. Frederiksen for calculation of survival rates, S. Garthe, K. Kober and A. Webb for information on at-sea distributions, C. M. Perrins for supplying survival data from Skomer, J.-K. Jensen and B. Olsen for information on puffins in the Faeroes and C. J. McCarty, T. DiMarzio, Z. Eppley, D. A. Oehler, and S. Cruciger for information on captive auks. Scottish Natural Heritage allowed us to work on the Isle of May National Natures Reserve and the British Trust for Ornithology supplied the recoveries from their Ringing Scheme that is funded by a partnership of the British Trust for Ornithology, the Joint Nature Conservation Committee (on behalf of Natural England, Scottish Natural Heritage and the Countryside Council for Wales, and also on behalf of the Council for Nature Conservation and the Countryside in Northern Ireland), The National Parks and Wildlife Service (Ireland) and the ringers themselves. We thank three anonymous reviewers for comments on an earlier version of the manuscript. This research represents a contribution to the BAS Ecosystems Programme.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. Garthe.
An erratum to this article can be found at http://dx.doi.org/10.1007/s00227-010-1390-z
Rights and permissions
About this article
Cite this article
Harris, M.P., Daunt, F., Newell, M. et al. Wintering areas of adult Atlantic puffins Fratercula arctica from a North Sea colony as revealed by geolocation technology. Mar Biol 157, 827–836 (2010). https://doi.org/10.1007/s00227-009-1365-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-009-1365-0