
Abstract
To estimate species turnover rates on scales of several tens of km in deep-sea benthic animals, we analyzed spatial and inter-annual changes in species diversity and composition of cerviniids, a typical group of deep-sea harpacticoids, at stations in and around Sagami Bay, central Japan. Associations with environmental factors were also investigated. Generally, bathymetrical patterns in diversity of benthos are unimodal and peak at depths of 2,000–3,000 m. In Sagami Bay, cerviniid diversity did not follow this trend; both species richness and evenness were negatively correlated with water depth. Multivariate analyses [detrended correspondence analysis (DCA) and non-metric multi-dimensional scaling] suggested that temporal changes in species composition of cerviniids are smaller than spatial changes that occur on horizontal scales of several tens of km. Community structure does not change completely on these scales in the bathyal zone around Sagami Bay. DCA also showed that bathymetrical changes in species composition can be regulated by certain factors associated with water depth.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Since the 1960s, when the astonishing species richness of organisms dwelling in deep-sea sediments was uncovered, explaining how the oligotrophic and apparently uniform environment of the deep sea supports such a rich fauna has been of primary interest in marine biology (cf. Gage and Tyler 1991; Gage 1996). Extrapolated estimates of total species numbers have suggested that the deep sea is a highly diverse environment (e.g., Grassle and Maciolek 1992; Seifried 2004). Many studies on spatiotemporal patterns of biodiversity in the deep sea have been based on comparisons of local or α diversity, which is often represented by the species diversity of individual samples or stations (e.g., Rex et al. 1993; Boucher and Lambshead 1995; Lambshead et al. 2000; Rose et al. 2005). β diversity (or turnover) is a measure of the extent to which the diversity of two or more spatial units differs in terms of their species composition (Magurran 2004), and spatial patterns of species turnover in the deep sea have been very poorly investigated (cf. Glover et al. 2002). On the other hand, γ diversity is the diversity of a landscape or other large area (Magurran 2004). Following Lande (1996), γ diversity can be treated as mean α diversity plus β diversity, showing that even though diversity of each local assemblage within a region (α diversity) is higher than that of other regions, total diversity of the whole region can be smaller when similarities in species composition among assemblages in the region are higher (namely β diversity is lower).
Recently, studies have analyzed regional biodiversity of the deep-sea floor based on data covering 1,000-km scales, which is comparable to data from neritic regions. In the abyssal plain in the central Pacific Ocean (depth 4,300–5,100 m, maximum distance between samples >3,000 km), 70–90% of polychaetes belonged to widespread or ubiquitous species (Glover et al. 2002). Furthermore, the regional diversity of nematodes in the same region was lower than the diversity of some coastal regions, despite lower local diversity in coastal water than in the deep sea (e.g., English Channel, range 495 km; Lambshead and Boucher 2003). The authors suggested that this is because similar patches of a similar habitat are repeated for a considerable distance in the deep sea, and the region may not be a “hyperdiverse” (>1 million species) environment. In contrast, in the continental slope of the northern Gulf of Mexico (depth 212–3,150 m, range >1,000 km), Baguley et al. (2006) reported little overlap in the community structure of harpacticoids even between stations on spatial scales of less than 50 km, suggesting that β and γ diversity of the organisms are large.
Several factors could produce the observed differences in regional diversity between these studies (polychaetes, nematodes vs. harpacticoids; abyssal vs. bathyal; Pacific vs. Atlantic). This suggests that to estimate global diversity, more information is needed not only on local diversity at more sites, but also on rates of species turnover with distance (β diversity) on the deep-sea floor, which covers over 50% of the Earth’s surface.
Little is known about the processes regulating spatial differences in the species composition of deep-sea benthic assemblages. Ecological studies at the species level dealing with deep-sea harpacticoids, a major meiofaunal taxon, are rare, although they have been studied in the last few decades (e.g., Coull 1972; Hessler and Jumars 1974; Thistle 1978; Montagna and Carey 1978; Eckman and Thistle 1991; Radziejewska and Drzycimski 2001; Ahnert and Schriever 2001; George and Schminke 2002; Rose et al. 2005; Baguley et al. 2006). The spatial heterogeneity of some environmental factors has been suggested to affect distribution patterns of deep-sea harpacticoid species (e.g., food supply, Baguley et al. 2006; grain size, Montagna 1982; biogenic structures, Thistle 1979). These surveyed areas are so restricted, however, considering the vast tracts of the environment. Therefore, the commonness and endemism of the effects of these factors on harpacticoid communities in various deep-sea regions, and their effects on different scales are still obscure.
In the San Diego Trough, Thistle (1978) found a nonrandom dispersion of harpacticoids at 100, 1-m, and 1-cm scales, suggesting that studies on harpacticoid distribution patterns should consider a hierarchy of spatial changes or concentrate on those of certain scales.
There is great variation in body size among harpacticoids. While adults of some species are retained with a 500-μm mesh, adults of other species pass through a 125-μm mesh. Considering their diversity in morphological characters and ecological habits derived from phylogeny (cf. Boxshall and Halsey 2004), it is informative to analyze differences in distribution patterns among subgroups (families or genera) as well as analyzing the whole community of harpacticoids. In the communities of deep-sea harpacticoids, several genera such as Bradya (Ectinosomatidae) and Stenhelia (Miraciidae) are observed commonly (Hicks and Coull 1983). Genera of Cerviniidae (or benthic Aegisthidae) are also typical members of deep-sea harpacticoid fauna, and can be abundant on clay or mud sediment (Boxshall and Halsey 2004). Usually cerviniids are not numerically abundant among harpacticoids, but they are dominant in biomass because they have the largest body sizes among deep-sea species (>1 mm). Thus, they are expected to occupy an important position in harpacticoid communities on the deep-sea floor, and because of their large body sizes, they can have different patterns in α diversity and specie turn over rates from the other smaller families.
We analyzed inter-annual and spatial differences in species diversity and composition of cerviniids on scales of several tens of km, and their relationship with several environmental factors (depth, food supply, grain size, and biogenic structures). Our study was conducted around Sagami Bay, Japan, located on the border of the northwestern Pacific Ocean, where studies of community ecology of deep sea organisms are rare relative to the Atlantic and eastern Pacific Oceans. We focused especially on investigating those hypotheses mentioned below.
-
1.
There is a general trend among macrofauna and meiofauna that α diversity shows a parabolic response to depth, peaking in the bathyal zone (Rex 1981; Boucher and Lambshead 1995; Baguley et al. 2006). Cerviniids in Sagami Bay would also show a similar pattern.
-
2.
Among investigated environmental factors, biogenic structures located at the sediment–water interface are expected to influence the distributions of certain species on smaller scales (e.g., cm scales, cf. Eckman and Thistle 1991). Thus, differences in the amount of biogenic structures would not have as strong an effect on cerviniid communities as the others on much larger scales (km scales).
-
3.
Organisms with better locomotive abilities are expected to have better dispersal abilities, which would lead to higher similarities in their species structure between habitats. Considering the large body sizes of cerviniids, which may allow them to have higher locomotive abilities, species composition of their communities would not turn over completely on scales of several tens of km.
Materials and methods
Sampling and sample processing
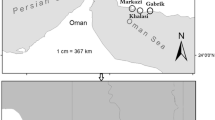
Sagami Bay was chosen as our study area because a relatively large number of deep-sea meiofaunal studies have been conducted around Japan (e.g., Kitazato and Ohga 1995; Shimanaga et al. 2004; Nomaki et al. 2006). Eleven sampling stations were located in Sagami Bay, and one sampling station [Station (St.) OB] was outside the bathyal zone of Sagami Bay (Fig. 1). Water depth ranged from 450 to 1,960 m. The largest distance between stations (Sts. A and OB) was about 60 km.

Sampling stations in Sagami Bay, Japan. The contours on the map were based on data from the Marine Information Research Center, Japan Hydrographic Association
Sampling was conducted using a multiple corer (Barnett et al. 1984), which can collect up to eight 52.8-cm2 sediment cores simultaneously. Usually, three cores were collected from each cast because of the limited sampling time and demand for cores. Sampling locations, depths, and number of cores taken from each location are listed in Table 1. Sampling dates ranged from 2000 to 2005. At most stations, sampling was done with only one cast. Three sampling casts in different years were done at St. C (depth 750 m; upper bathyal zone) and St. SB (depth 1,430 m; lower bathyal zone) to investigate inter-annual changes in species composition.
The top 1 cm of the sediment cores was examined, where most harpacticoid including cerviniids are distributed (Shimanaga et al. 2004). These sediment samples were fixed immediately with 5% buffered seawater formalin and preserved in sealed plastic bags on board the ship. Rose Bengal (final concentration 0.05 g/l) was added to this solution to stain the samples. Before these processes, the water column above the sediment in each core (about 30 cm) was siphoned off. The water layer just above the sediment (<1 cm) was removed with a pipette and added to the sediment samples because of the difficulty in extracting the layer without contamination of sediments. Subsamples (0.36 or 0.71 ml) were also taken with syringes to measure the amount of chloroplastic pigment equivalents (CPE) in the top sediments as an indicator of the amount of organic matter derived from primary production.
In the laboratory, the fixed sediment samples were wet-sieved using mesh sizes of 1, 0.5, 0.25, and 0.125 mm. The samples retained were transferred into flat-bottomed Petri dishes, and the organisms were sorted and counted under a binocular stereoscopic microscope. Most adult cerviniids were retained on a 0.25-mm mesh and very few juvenile cerviniids passed through 0.125-mm mesh. All sediment samples were checked to sort adult specimens without subdivision. For one core of casts J and SB04-c (cf. Table 1), however, 1/2 and 1/4 of the sediments were accidentally lost, respectively. Before each extraction, a small amount of the sediment (<1 ml) was taken with a spoon or pipette for analyses of median sediment grain sizes (MD).
Unfortunately, some samples for environmental factors were not collected (denoted “ND” in Table 1). However, since most sampling was done successfully, the influence of the missing samples on the analyses was expected to be small.
Remarks on the taxonomy and species identification of cerviniids
Under a compound microscope, only adult specimens were identified to species because of the difficulty in identifying juvenile copepodites. Most species-level studies have been based on adult harpacticoids. Taxon determination was done using identification keys (Boxshall and Halsey 2004; Burgess 1998; Huys et al. 1996; Wells 1976–1985), original articles in a catalog by Bodin (1997), and a database on the world wide web managed by Walter (http://www.nmnh.si.edu/iz/copepod/). Most identified species seemed to be new to science, and their seta formulae are shown in Table 2. Lengths of antennae and antennules excluding terminal setae or spines of specimens were measured based on camera-lucida drawings. As a description of these species was beyond the scope of this study, they were regarded as “working species”. Taxonomical work will be done by specialists in the near future.
Seifried and Schminke (2003) found that the family “Cerviniidae” is paraphyletic and is a junior synonym of Aegisthidae. Many species of “Aegisthidae” (or Aegisthinae), however, are regarded as holoplanktonic (Boxshall and Halsey 2004), and no specimens of the group were found in our samples. Thus, we use the terms “cerviniids” and “Cerviniidae” here for their familiarity, as the terms “reptiles” or “fish” would be used.
Specimens identified as “Cervinia sp. 1” by Shimanaga et al. (2004) are an assemblage of Neocervinia itoi and Cervinia sp. A. An individual identified as “Cervinia sp. 2” in that article was identified as a variation of N. itoi. Thus, Table 2 in Shimanaga et al. (2004) can be corrected partly as shown in Table 3 in this article. The data were ignored in this study because they were based on specimens extracted from subdivided sediments from only one core per cast, and are too small in sample size to compare.
Statistical analyses
Although a few cores were taken from the same casts as mentioned above, they were not regarded as replicates in this study because of a possibility that they are not independent (cf. Zar 1999). Therefore, mean densities of species (individual numbers per 52.8 cm2) and mean values of environmental factors for each sampling cast were calculated for further statistical analyses. The damaged cores mentioned above were also included in the analyses, and densities of species for the casts that contained these cores (J and SB04-c) were calculated based on adjusted sampled area (see Table 1).
For α species diversity indices, the total number of species per cast (an indicator of species richness) and the reciprocal of the relative abundance of the most abundant species (an indicator of evenness, cf. Magurran 1988) were calculated. For Sts. C and SB, where three casts (replicates) were conducted for each sampling period, differences in the means of these variables among year were investigated by the Kruskal–Wallis test (at α = 0.05), which is not dependent on a given distribution (Sokal and Rohlf 1995).
To find “latent” variables (ordination axes) that represent the best predictor for the values of all species, ordination was used (Lepš and Šmilauer 2003). While linear methods [e.g., principal component analysis (PCA)] are applicable over short environmental ranges, where species abundance appears to vary monotonically with variation in the environment, unimodal methods [e.g., detrended correspondence analysis (DCA)] are applicable over a wider range of environments (Ter Braak and Prentice 1988). In this study, ordination methods were chosen based on the gradient length (the range of sample scores) for the first axis in DCA (Ter Braak and Prentice 1988). When the length is over 3 SD, linear methods are ineffective. On the other hand, if the value is less than about 1.5 SD, unimodal methods yield poor results. The range of 1.5–3 SD can be regarded as a window over which both PCA and DCA can be used. This gradient length also can be used as an indicator of species turnover: a complete turnover in species composition occurs over samples in about 4 SD units (Legendre and Legendre 1998). DCA were done based on mean densities of species for each sampling cast with MVSP version 3.1 (Kovach Computing Services, UK).
For comparison with results of Baguley et al. (2006), which showed important information about spatial changes in species composition of deep-sea harpacticoids, the non-metric multi-dimensional scaling (MDS) and group average clustering (Clarke and Warwick 2001) were also done with PRIMER 6 (PRIMMER-E Ltd, UK), based on the same data for DCA. For the similarity index, the Bray–Curtis coefficient (Clarke and Warwick 2001) was used. Prior to scaling and clustering, data were 4th-root transformed to prevent the contribution of dominant species from being over-emphasized, as in Baguley et al. (2006).
Environmental factors and their effects on cerviniid community structure
Treatment of the CPE samples was based on Greiser and Faubel (1988). The details of grain size analysis were described by Sakamoto et al. (2005). To include all grain sizes for contribution to environmental factors, we omitted procedures for dissolution of biogenic shells such as acidification and sodium carbonate leaching. The dry weights of biogenic structures (DWBS, mainly tubes of polychaetes) retained on a 1-mm mesh were also investigated.
As mentioned in the “Introduction”, the spatial heterogeneity of these environmental factors is expected to influence the distribution patterns of deep-sea harpacticoid species. As an indicator of cerviniid community structure, scores for each cast for the first ordination axis were used. The Spearman’s rank correlation coefficients (cf. Zar 1999) were calculated between the axis 1 scores and environmental factors (CPE, MD, DWBS, and water depth). Although significant correlations do not always mean that one variable is the cause of the other, they indicate strong evidence that species turnover is associated with the factors, and the absolute values of coefficients are expected to show their ranks in the degree of their associations with species composition. Among pairs of environmental factors investigated in this study, there was a significant correlation only between MD and DWBS. The other parameters for sediment characters (e.g., sorting coefficients) were not used for analyses because they were strongly correlated with each other. All statistical analyses (at α = 0.05) were performed with SPSS version 11 (SPSS Inc., USA).
Results
Bathymetrical patterns in the standing stock
In total, 3,053 cerviniids were extracted from core samples, including 353 adults (12% of all individuals; Table 4). For the C05, SB04, and SB05 series cores, the number of cerviniids in the water above the sediment was also checked. While 1,634 cerviniids (120 adults) were extracted from sediments in these cores, only 18 adults and 52 juvenile copepodites were observed in the water above the sediments. These suggest that most cerviniids (87% of adults, 96% of total copepodites) exist on or in the top-most sediments, although we cannot deny they may have been inactive when core samples were processed on deck after drastic changes in pressure and temperature that they had experienced during sample recovery.
Bathymetrical patterns in density of total copepodites (most were juveniles) and adults are shown in Fig. 2. Values from St. SB suggest that temporal changes in the density of juveniles were large at some stations, thus strict comparisons could not be made. There was an overall tendency, however, for density to be lower at stations shallower than 1,000 m than at the deeper stations in Sagami Bay, especially Sts. E, F, G, and J, which are located on or near sea bights (see Fig. 1). A significant positive correlation was detected between total density and water depth (r = 0.57). Adult density also tended to be higher at Sts. E, F, G, and J, although the values were not significantly correlated with water depth. At St. OB, which was located outside of Sagami Bay, only a few specimens were obtained. Among pairs between density of total copepodites or adults and any other environmental factor (CPE, MD, or DWBS), a significant correlation was detected only between adult density and CPE (r = 0.45).

Bathymetrical patterns of cast-based mean densities of a total copepodites and b adult cerviniids, c species number per cast, and d evenness. The replicate samples from Sts. C and SB are enclosed by the dotted lines. For species richness and evenness, St. OB is not plotted because of no datum
Species diversity
Cerviniid adults were identified to nine species (Fig. 3, Table 4). Both the number of species per cast and the reciprocal of the relative abundance of the most abundant species tended to be higher at shallower stations (Table 4; Fig. 2c, d), and they were significantly negatively correlated with water depth (number of species, r = −0.47; evenness, r = −0.41), although the total number of specimens was very small (<10 individuals) at some stations (Sts. B, D, and H). On the other hand, regression analyses did not support their unimodal relationships with depth: quadratic regression curves for the species number and the reciprocal of the relative abundance of the most abundant species against depth were Y = 8.8 − 1.2 × 10−2 X + 4.7 × 10−6 X2 and Y = 5.0 − 6.5 × 10−3 X + 2.7 × 10−6 X2, respectively, and both are concave. Neither species richness nor evenness was correlated significantly with any environmental factor (MD, BWBS, or CPE) investigated in this study. Inter-annual changes in these parameters were investigated for Sts. C and SB, where multiple samples were taken. The Kruskal–Wallis test did not reveal any significant temporal differences.

Cerviniid species in Sagami Bay. a Stratiopontotes sp. A, b Cerviniopsis sp. A, c Cerviniopsis sp. B, d Cervinia? sp. A, e Cervinia? sp. B, f Neocervinia itoi, g Cervinia spp. h Eucanuella? sp. A and i Hemicervinia? sp. A
Distribution patterns of cerviniid species
Among the nine identified species, N. itoi Lee and Yoo was the most abundant. About 80% of all adults were N. itoi. They were found at all stations in Sagami Bay and were dominant at most stations, except St. A, where Cerviniopsis sp. A was the most abundant (Table 4, Fig. 4). Six species, including Cerviniopsis sp. A, were distributed at stations shallower than 1,000 m, and three of the six were found only at one station (Fig. 4). No adults and only three juveniles were obtained from cores at St. OB. Although species structures in juveniles were not investigated in this study, two juveniles from St. OB were copepodite V, which could be identified as N. itoi and Cervinia spp. based on their seta formula and characteristics of exopods of P5 (fused or separated from basoendopods, cf. Table 2).

Distribution patterns of species in Sagami Bay. Filled squares represent stations where the species were the most abundant. Filled circles represent stations where the species existed. Open circles represent stations where the species did not exist
DCA based on mean adult densities of species for each cast revealed a gradient length for the first axis of 2.39 SD (<4 SD), suggesting that a complete turnover in species composition did not occur over the study sites, although the cast from St. OB was omitted in the analysis because of the lack of adult specimens. The gradient length for the first axis (>1.5 SD) also suggests the DCA results are useable for further analyses (see “Materials and methods”).
The DCA plot is shown in Fig. 5. Axes 1 and 2 explained 37.7 and 16.5% of the variation among sites, respectively. The majority of casts were plotted very closely to the point of N. itoi, because of their highest relative abundance in these samples. Most casts taken at St. SB were plotted within the cluster, except SB00-b where Cervinia spp. were the most abundant, although the sample size on which that datum was based was quite small (only six individuals, see Table 4). On the other hand, the plot patterns of the St. C series suggest that species compositions at the site differed between 2002 and 2005. The temporal difference, however, was smaller than the species difference that occurred over bathyal Sagami Bay at 10-km scales. As a whole, there was no distinct group among the sites in species structure. However, there appeared to be a gradient along axis 1: the casts from shallower sites (Sts. A, B, and C) were scattered more widely at the right side of the plot.

DCA joint plot with respect to axes 1 and 2, based on mean density of each species in the sediment from each cast. Black circles and grey diamonds denote cast scores and species scores, respectively. Minor species are not shown
The results of the MDS and clustering were similar to those of DCA (Fig. 6). Casts from the same stations or stations at similar water depths tended be plotted closely. Four groups are formed at around 60% similarity level, and three of the four were composed by casts form shallower sites. All casts were clustered at about 40% similarity.

MDS plot with superimposed clusters based on Bray Curtis similarity. Black circles denote casts. Continuous and dashed lines denote similarity levels of 40 and 60%, respectively, showing that all casts are clustered at ≥40% similarity, and 4 groups are formed at around 60%
While a strong and significant correlation was found between the DCA axis l scores of casts (an indicator of community structure) and water depth, the other correlation analyses failed to detect significant differences between the axis l values and any investigated environmental factor (Fig. 7). For MD, the value of St. D was much larger than those of the other sites (Table 1) and seemed to be an outlier. No significant correlation could be detected, however, between MD and axis 1, even though the outlier was omitted.

DCA axis 1 values for casts plotted against a water depth, b median sediment grain sizes (MD), c dry weights of biogenic structures (DWBS) and d chloroplastic pigment equivalents (CPE) concentration in the sediments
Discussion
Standing stock and diversity
There is a general trend along continental margins for the abundance of metazoan meiofauna to decrease with water depth. The supply of organic matter from surface productivity is one of the important factors regulating this pattern (Soltwedel 2000). Harpacticoids in the bathyal zone in the Gulf of Mexico also show a significant negative correlation with water depth (Baguley et al. 2006). In the bathyal Sagami Bay, however, total cerviniids (mostly juveniles) showed the inverse trend: they tended to increase in abundance at sites located deeper than 1,000 m. The trend was more obscure in adults, but both total and adult density seemed to be higher at stations on or below sea bights with steep slopes (Sts. E, F, G, and J).
Based on sediment trap samples, Nakatsuka et al. (2003) suggested that re-suspension of phytodetritus occurs on the deep-sea bottom of Sagami Bay. The lack of a significant correlation between water depth and CPE concentration in the sediments in our study is partly because organic matter sinking to the bottom is rebounded and transported from the steep upper slopes to the deeper areas in the bay. Through lateral flux down steep slopes, organic matter is concentrated in the moderate slope zones, where an increase in organisms can be expected. However, CPE amount was not significantly correlated with the density of total cerviniids, although significance was detected between the amount and the adult density. Data from St. SB showed that cerviniid density and CPE level may fluctuate temporally in some areas on the bathyal floor. Thus, strict analyses of their relationship were not possible based on our samples, which were taken in different years, because temporal heterogeneity may mask the relationship between spatial differences. Regardless, the supply of organic matter from surface waters is not the sole factor regulating standing stocks of cerviniids.
The results of our analyses of species diversity seem to be more conservative, but deny a unimodal pattern along water depth. Between depths of 500 and 1,500 m, species richness and evenness of adult cerviniids in bathyal Sagami Bay tended to be higher in the upper 1,000 m. This tendency was not masked by inter-annual changes in these parameters observed at Sts. C and SB. Although no adults or juvenile Eucanuella, Cerviniopsis, or Stratiopontotes were obtained at St. OB, many juveniles of these genera were found at stations where congeneric adults were obtained. Therefore, low diversity cerviniid communities would also spread outside of Sagami Bay, at least up to 2,000 m depth.
Generally, the local-scale (1–10 m2, cf. Levin et al. 2001) diversity of primary macrofauna shows a parabolic response to depth, peaking at 2,000–3,000 m (cf. Rex 1981). A similar pattern was seen in the diversity of harpacticoids in the Gulf of Mexico, although it peaked at about 1,200 m (Baguley et al. 2006). Levin et al. (2001) attributed this pattern to food availability: at low food levels, diversity is low because there is insufficient food to support populations of many species; as food supply increases, diversity should increase because more species can maintain their populations, which may explain the increase in diversity from the abyssal to the bathyal zones. On the other hand, at high food levels, diversity may decline with increases in food availability, reflecting the strength of species interactions (primarily interspecific competition), leading to increased dominance by a few species. The increase in diversity with increasing water depth in the bathyal zone could be explained partly by this process. Baguley et al. (2006) showed a significant inverse relationship between harpacticoid diversity and particulate organic matter flux in the Gulf of Mexico.
Here, we discuss the possibility that the bathymetric pattern in the diversity of cerviniids seen here was caused by food availability. While most studies on the bathymetric patterns of diversity along continental slopes show the right sides of unimodal patterns of diversity with food availability (from intermediate to high food levels), did our results reveal the left side of the pattern (from low and intermediate food levels)? This possibility is very unlikely because the CPE concentrations in sediments were several-fold greater than those reported from other regions when adjusted for water depth (cf. Soltwedel 2000), suggesting that Sagami Bay is eutrophic. High dominance of N. itoi observed at stations located between 1,000 and 1,300 m might reflect extraordinarily high food input, as discussed above. This speculation is not supported, however, by the fact that CPE concentrations at different stations in Sagami Bay at the same time periods were similar, regardless of differences in water depth. Additionally, there was no significant correlation between the diversity index and CPE. Furthermore, statistical analyses did not suggest a correlation between diversity and any other environmental factor. The reason why cerviniid species diversity did not show any association with these factors is discussed below.
Community structure
Since our study was based on only about 350 specimens, the resolution is not high enough to describe exact similarities in species composition between stations. We want to emphasize, however, that our collection is larger in the number of cerviniid specimens than those of recent studies of spatial changes in species structure of harpacticoid assemblages, for example, George (2005) collected 75 cerviniids in the Magellan region; 25 specimens were collected from the Angola Basin by Rose et al. (2005); the study of Baguley et al. (2006) was based on 3,654 adult harpacticoids, and cerviniids contributed about 1% of them (≈37 individuals). Therefore, our results should reflect general trends in cerviniid communities.
Both DCA and MDS revealed the bathymetrical changes in species composition of cerviniids. There was a significant correlation between DCA axis 1, an indicator of species composition, and water depth, suggesting that cerviniid community structure can be regulated by certain environmental gradients along water depth. Correlations of DCA axis 1 were also investigated with environmental factors thought to regulate the distributions of harpacticoids. Among these factors, differences in the amount of biogenic structures, as indicated by DWBS, were hypothesized not to have as strong an effect as the others. These analyses, however, failed to detect a significant correlation not only between DCA axis l and DWBS, but also between DCA axis 1 and any other factor. The fact that there was no bathymetrical pattern in any of these factors also indicates they are not the environmental gradients regulating the community structure.
Quantitative data on spatial differences in cerviniid community structure that are comparable to our study are rare, probably because of their low relative abundance in harpacticoid assemblages and the difficulty in obtaining enough specimens to evaluate species composition. George (2005) ignored cerviniids in his work because only juveniles were represented. Based on specimens collected from 5 to 3,576 m deep over a period of 7 years, Montagna (1982) observed a bathymetric cline in four cerviniid species from the Beaufort Sea, Alaska, and attributed the distribution pattern to adaptations correlated with the sedimentary environment, where shelf sediments are coarser than deeper sediments with little or no sand. The shelf species are adapted to burrowing into coarse sediments and have larger antennae than antennules. The deeper species are adapted to epibenthic habitats, with long caudal rami that prevent their bodies from sinking into the fine sediments.
We noted a different trend in the morphological characters of cerviniids in Sagami Bay. Five species that have shorter antennae than antennules and long caudal rami (see Table 2) tended to be found from casts at shallower stations (Table 4). There was no significant difference in median sediment size between these casts and the others (the Kruskal–Wallis test, P = 0.88), suggesting those species with shorter antennae and longer caudal rami have little association with the fine sediment. Since our data did not cover the shelf regions, the possibility remains that similar trends exist in cerviniid communities from the entire Sagami Bay. However, it can be safely said that adaptations to sediment characters have no or little relationship with the bathymetrical change in species composition observed in this study.
As shown above, we do not have evidence suggesting which factors directly regulate species diversity and structure in cerviniid communities. However, food availability is still a candidate factor, although an association between species composition and CPE was not detected. We observed rotaliid foraminifera in the guts of some cerviniid specimens. Analyses using stable carbon and nitrogen isotope ratios indicate that their trophic level is higher than that of primary detritus consumers such as foraminifera (Nomaki, unpublished data). These suggest cerviniids do not eat fresh organic matter directly but eat other smaller organisms, which may regulate their distributions.
Multivariate analyses also suggested little inter-annual change in species composition at St. SB. Data from 1996 to 1998, which were omitted in this study as discussed in the Materials and Methods, were consistent with these results (Table 3). At St. C, a considerable difference in species composition appeared between 2002 and 2005. It may be that in the upper bathyal slopes, where species evenness is high, the relative abundance of each cerviniid species changes more easily than in the deeper region of the bay. As shown in Table 1, the sampling positions for St. C in each year tended to be clumped rather than scattered. Therefore, the apparent temporal difference might reflect a spatial difference caused by topographic complexity on the steep continental slopes on 100-m scales. If anything, our results suggest that the temporal changes in cerviniid community structure over a period of several years are smaller than changes occurring on scales of several tens of km.
The gradient length calculated by DCA based on species composition of adult specimens was 2.39 SD (<4 SD), suggesting that cerviniid species do not change completely on a 30-km horizontal scale or a 1,000-m bathymetrical scale in Sagami Bay. The cluster analyses demonstrated that all casts were clustered at about 40% similarity, which is consistent with the DCA results. Although no adults were obtained at St. OB, juvenile specimens collected at this station implied that cerviniid communities dominated by N. itoi and Cervinia spp. are distributed outside the bay and the turnover of species is not completed at least on an approximately 60-km scale.
Among 43 deep-sea stations in the northern Gulf of Mexico, no pair of stations was more than 40% similar, suggesting very little overlap between stations on spatial scales of less than 50 km (Baguley et al. 2006). Considering the spatial scale on which our stations were set, our data seem to show similar results with regard to species turnover rates of deep-sea harpacticoids. However, we cannot reject our impression that both α diversity and species turnover rates (β diversity) of cerviniids are low in the lower bathyal zone around Sagami Bay, because all stations located deeper than 1,000 m were dominated by N. itoi and were clustered at 60% similarity. Wide spatial distributions of cerviniid species were also observed in the Beaufort Sea, where four of five species found were observed at two or more stations separated by more than 100 km, although many species of other harpacticoid families were also distributed on such large scales (Montagna and Carey 1978). On the other hand, six out of nine species (e.g. Cerviniopsis sp. A, Stratiopontotes sp. A) were distributed only at one or two stations located shallower than 1,000 m in Sagami Bay, suggesting these species have more distinct distributions than N. itoi. Thus, β diversity of cerviniids in the bay might be higher in the upper bathyal zone, although there is a possibility of incomplete sampling of the rare species at those stations (cf. Glower et al. 2002).
Because of a lack of planktonic development stages, dispersal of most harpacticoids depends on suspension and transport by current flow (Baguley et al. 2006). On the other hand, cerviniids are one of the largest groups among the deep-sea harpacticoids. Large size, as in scavenging amphipods, has been related to the need for high motility in locating sparsely dispersed food (Gage and Tyler 1991). In our study, about 15% of adult cerviniids were obtained from the water above the sediment in core samples. N. itoi, the dominant species in the study area, was described based on an individual collected in a net towed near the sea bottom by a submersible (Lee and Yoo 1998). These facts imply that some cerviniids have good swimming abilities, which may contribute partly to the distribution of cerviniid species on spatial scales of several tens of km. These abilities may also be related to the poor associations of cerviniids with spatial heterogeneity in the environmental factors studied here. Their distribution might be influenced by spatial heterogeneity that occurs on scales of a few tens to thousands of m (e.g., topographic complexity), and/or by the frequency of periodic events such as sediment re-suspension, which were out of the scope of this study, and are expected to show bathymetric differences.
We must investigate the distribution patterns of other harpacticoid families, especially groups that have smaller body sizes (and are expected to have poorer locomotor ability) than cerviniids, to determine if the rate of species turnover of cerviniids is actually lower than those of the other harpacticoids in the bay.
Finally, we must mention that since our study only examined the spatial distribution of cerviniids on scales of several tens of km, strict comparisons cannot be made to studies based on data from the whole harpacticoid assemblage on much larger spatial scales (>1,000 km). However, this study provides information on spatial differences in the community structure of a typical group of deep-sea harpacticoids based on quantitative analyses and is key to evaluating regional diversity of harpacticoids, differences in species turnover rate among families and/or genera, and the processes that regulate spatial heterogeneity in species composition in the northwestern Pacific Ocean.
Conclusions
-
1.
Both species richness and evenness of cerviniid communities tended to decrease with increasing water depth in the bathyal Sagami Bay.
-
2.
DCA showed that certain environmental factor(s) associated with water depth may regulate spatial differences in species composition of cerviniids, but failed to detect what the factors are.
-
3.
Cerviniid species did not change completely on horizontal scales of several tens of km or 1,000-m bathymetrical scales, suggesting that β diversity of cerviniids is low around Sagami Bay. It is still unclear, however, whether it is the lowest among different harpacticoid families in the study area.
References
Ahnert A, Schriever G (2001) Response of abyssal Copepoda Harpacticoida (Crustacea) and other meiobenthos to an artificial disturbance and its bearing on future mining for polymetallic nodules. Deep Sea Res II 48:3779–3794
Baguley JG, Montagna PA, Lee W, Hyde LJ, Rowe GT (2006) Spatial and bathymetric trends in Harpacticoida (Copepoda) community structure in the Northern Gulf of Mexico deep-sea. J Exp Mar Biol Ecol 330:327–341
Barnett PRO, Watson J, Connelly D (1984) A multiple corer for taking virtually undisturbed samples from shelf, bathyal and abyssal sediments. Oceanol Acta 7:399–408
Bodin P (1997) Catalogue of the new marine Harpacticoid Copepods (1997 Edn.). Documents de Travail de l’Institut Royal des Sciences Naturelles de Belgique 89:1–304
Boucher G, Lambshead PJD (1995) Ecological biodiversity of marine nematodes in samples from temperate, tropical, and deep-sea regions. Conserv Biol 9:1594–1604
Boxshall GA, Halsey SH (2004) An Introduction to Copepod diversity. The Ray Society, London
Burgess R (1998) Two new species of harpacticoid copepods from the Californian continental shelf. Crustaceana 71:258–279
Clarke KR, Warwick RM (2001) Change in marine communities: an approach to statistical analysis and interpretation, 2nd edn. PRIMER-E, Plymouth
Coull BC (1972) Species diversity and faunal affinities of meiobenthic Copepoda in the deep sea. Mar Biol 14:48–51
Eckman JE, Thistle D (1991) Effects of flow about a biologically produced structure on harpacticoid copepods in San Diego Trough. Deep Sea Res 38:1397–1416
Gage JD (1996) Why are there so many species in deep-sea sediments? J Exp Mar Biol Ecol 200:257–286
Gage JD, Tyler PA (1991) Deep-sea biology. Cambridge University Press, Cambridge
George KH (2005) Sublittoral and bathyal Harpacticoida (Crustacea: Copepoda) of the Magellan region. Composition, distribution and species diversity of selected major taxa. Sci Mar 69:147–158
George KH, Schminke HK (2002) Harpacticoida (Crustacea, Copepoda) of the Great Meteor Seamount, with first conclusions as the origin of the plateau fauna. Mar Biol 141:887–895
Glover AG, Smith CR, Paterson GLJ, Wilson GDF, Hawkins L, Sheader M (2002) Polychaete species diversity in the central Pacific abyss: local and regional patterns, and relationships with productivity. Mar Ecol Prog Ser 240:157–170
Grassle JF, Maciolek NJ (1992) Deep-sea species richness: regional and local diversity estimates from quantitative bottom samples. Am Nat 139:313–341
Greiser N, Faubel A (1988) Biotic factors. In: Higgins RP, Thiel H (eds) Introduction to the study of Meiofauna. Smithsonian Institution Press, Washington, pp 79–114
Hessler RR, Jumars PA (1974) Abyssal community analysis from replicate box cores in the central North Pacific. Deep Sea Res 21:185–209
Hicks GRF, Coull BC (1983) The ecology of marine meiobenthic harpacticoid copepods. Oceanogr Mar Biol Ann Rev 21:67–175
Huys R, Gee JM, Moore CG, Hamond R (1996) Marine and brackish water Harpacticoid Copepods, Part 1. Field Studies Council, Shrewsbury
Kitazato H, Ohga T (1995) Seasonal changes in deep-sea benthic foraminiferal populations: results of long-term observations at Sagami Bay, Japan. In: Sakai H, Nozaki Y (eds) Biogeochemical processes and Ocean flux in the Western Pacific. Terra Scientific Publishing Comp, Tokyo, pp 331–342
Lambshead PJD, Boucher G (2003) Marine nematode deep-sea biodiversity—hyperdiverse or hype? J Biogeogr 30:475–631
Lambshead PJD, Tietjen J, Ferrero T, Jensen P (2000) Latitudinal diversity gradients in the deep sea with special reference to North Atlantic nematodes. Mar Ecol Prog Ser 194:159–167
Lande R (1996) Statistics and partitioning of species diversity, and similarity among multiple communities. Oikos 89:5–13
Lang K (1934) Marine Harpacticiden von der Campbell-Insel und einigen anderen südlichen Inseln. Acta Univ lund (N.S.) (Avd. 2) 30:1–56
Lee W, Yoo KI (1998) A new species of Neocervinia (Copepoda: Harpacticoida: Cerviniidae) from the hyperbenthos of the Hatsushima cold-seep site in Sagami Bay, Japan. Hydrobiol 377:165–175
Legendre P, Legendre L (1998) Numerical ecology, 2nd English edn. Elsevier, Tokyo
Lepš J, Šmilauer P (2003) Multivariate analysis of ecological data using CANOCO. Cambridge University Press, Cambridge
Levin LA, Etter RJ, Rex MA, Gooday AJ, Smith CR, Pineda J, Stuart CT, Hessler RR, Pawson D (2001) Environmental influences on regional deep-sea species diversity. Annu Rev Ecol Syst 32:51–93
Magurran AE (1988) Ecological diversity and its measurement. Princeton University Press, Princeton
Magurran AE (2004) Measuring biological diversity. Blackwell, Malden
Montagna P (1982) Morphological adaptation in the deep-sea benthic harpacticoid copepod family Cerviniidae. Crustaceana 42:37–43
Montagna PA, Carey AG JR (1978) Distributional notes on Harpacticoida (Crustacea: Copepoda) collected from the Beaufort Sea (Arctic Ocean). Astarte Astarte 11:117–122
Nakatsuka T, Kanda J, Kitazato H (2003) Particle dynamics in the deep water column of Sagami Bay, Japan. II: Seasonal change in profiles of suspended phytodetritus. Prog Oceanogr 57:47–57
Nomaki H, Heinz P, Nakatsuka T, Shimanaga M, Ohkouchi N, Ogawa NO, Kogure K, Eiko Ikemoto E, Kitazato H (2006) Different ingestion patterns of 13C-labeled bacteria and algae by deep-sea benthic foraminifera. Mar Ecol Prog Ser 310:95–108
Radziejewska T, Drzycimski I (2001) Changes in genus-level diversity of meiobenthic free-living nematodes (Nematoda) and harpacticoids (Copepoda: Harpacticoida) at an abyssal site following experimental sediment disturbance In: Chung JS (eds) Proceedings of the 4th (2001) ISOPE Ocean Mining Symposium. International Society of Offshore and Polar Engineers. California, pp 38–43
Rex MA (1981) Community structure in the deep-sea benthos. Annu Rev Ecol Syst 12:331–353
Rex MA, Stuart CT, Hessler RR, Allen JA, Sanders HL Wilson DF (1993) Global-scale latitudinal patterns of species diversity in the deep-sea benthos. Nature 365:636–639
Rose A, Seifried S, Willen E, George KH, Veit-Köhler G, Bröhldick K, Drewes J, Moura G, Martínez Arbizu, Schminke HK (2005) A method for comparing within-core alpha diversity values from repeated multicorer samplings, shown for abyssal Harpacticoida (Crustacea: Copepoda) from the Angola Basin. Org Divers Evol 5:3–17
Sakamoto T, Ikehara M, Aoki K, Iijima K, Kimura N, Nakatsuka T, Wakatsuchi M (2005) Ice-rafted debris (IRD)-based sea-ice expansion events during the past 100 kyrs in the Okhotsk Sea. Deep Sea Res II 52:2275–2301
Seifried S (2004) The importance of a phylogenetic system for the study of deep-sea harpacticoid diversity. Zool Stud 43:435–445
Seifried S, Schminke HK (2003) Phylogenetic relationships at the base of Oligoarthra (Copepoda, Harpacticoida) with a new species as the cornerstone. Org Divers Evol 3:13–37
Shimanaga M, Kitazato H, Shirayama Y (2004) Temporal patterns in diversity and species composition of deep-sea benthic copepods in bathyal Sagami Bay, central Japan. Mar Biol 144:1097–1110
Sokal RR, Rohlf FJ (1995) Biometry, 3rd edn. W. H. Freeman and Company, New York
Soltwedel T (2000) Metazoan meiobenthos along continental margins: a review. Prog Oceanogr 46:59–84
Ter Braak CJF, Prentice IC (1988) A theory of gradient analysis. Adv Ecol Res 18:271–317
Thistle D (1978) Harpacticoid dispersion patterns: implications for deep-sea diversity maintenance. J Mar Res 36:377–397
Thistle D (1979) Harpacticoid copepods and biogenic structures: implications for deep-sea diversity maintenance. In: Livingston RJ (ed) Ecological processes in coastal and marine systems. Plenum, New York, pp 217–231
Wells JBJ (1976) Keys to aid in the identification of marine Harpacticoid Copepods. The Aberdeen University Press, Aberdeen
Wells JBJ (1978–1985) Keys to aid in the identification of marine Harpacticoid Copepods. Amendment Bulletins 1–5 Zool Publ Victoria Univ Wellington
Zar JH (1999) Biostatistical analysis, 4th edn. Prentice-Hall, Englewood Cliffs
Acknowledgments
We are grateful to the officers and crew members of the research vessel Tansei-maru of the independent administrative institution, the Japan Agency for Marine–Earth Science and Technology (JAMSTEC). We also thank Dr. W. Lee who helped to identify males of N. itoi. Special thanks to two anonymous reviewers who kindly checked our manuscript. This study was partly supported by a grant from the Ministry of Education, Sports, Culture, Science, and Technology of Japan (No. 16770012), and complied with the current laws of Japan where it was performed.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. Nishida.
Rights and permissions
About this article
Cite this article
Shimanaga, M., Nomaki, H. & Iijima, K. Spatial changes in the distributions of deep-sea “Cerviniidae” (Harpacticoida, Copepoda) and their associations with environmental factors in the bathyal zone around Sagami Bay, Japan. Mar Biol 153, 493–506 (2008). https://doi.org/10.1007/s00227-007-0817-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-007-0817-7