
Abstract
Mangrove forests, with their ecological significance and economic benefits, are vital inter-tidal wetland ecosystems. Lumnitzera littorea (Combreataceae) is a non-viviparous mangrove distributed in tropical Asia and North Australia. Due to natural and human impacts, populations of this species have been isolated, fragmented, and highly disturbed. In China, L. littorea is an endangered species, restricted to small regions of Hainan Island. The genetic composition of five populations of this species from the Indo-West Pacific (South China, Malay Peninsula, Sri Lanka, North Australia) was assessed using inter simple sequence repeat (ISSR) makers. At the species level, expected mean heterozygosity (He) was 0.240 with 75.6% of loci polymorphic (P). However, genetic variation was much lower at the population level (P = 37.1%, He = 0.118). A high coefficient of gene differentiation (Gst = 0.515) and low level of gene flow (Nm = 0.470) indicated significant genetic differentiation among populations. AMOVA also indicated that more than half the total variation (58.4%) was partitioned among populations. The high degree of differentiation observed among populations emphasizes the need for appropriate conservation measures that incorporate additional populations into protected areas, and achieve the restoration of separate, degraded populations.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Mangroves are salt-tolerant forest ecosystems found mainly in tropical and subtropical inter-tidal regions throughout the world. Mangrove forests are among the most productive and diverse wetlands on earth. They are vital inter-tidal wetland ecosystems and directly impact the welfare of coastal people in the tropics and subtropics (Ge and Sun 1999). However, these unique coastal forests also are among the most threatened habitats worldwide. These ecosystems are highly fragile and have been subjected to a long duration of human perturbation as well as incessant physiological and morphological stresses due to salinity aeration and the onslaught of wave action (Mongia et al. 1991).
Lumnitzera littorea (Jack.) Voigt (Combreataceae) is a non-viviparous mangrove species distributed in tropical Asia and Australia. Lumnitzera littorea is characteristic of landward, high salinity areas, often in association with other mangrove species such as Avicennia marina (Forsk.) Vierh., Clerodendron inerme (L.) Gaertn., Excoecaria agallocha L., Scyphiphora hyolrophyllacea Gaertn. F. and Lumnitzera racemosa Wild (Tomlinson 1986). The species occurs within the calm coastal waters, typically on silt with 0.5–2.7% edaphic salinity and 21–25°C average temperature. As non-viviparous, L. littorea does not produce large diaspores and its fruits can disperse far with suitable ocean currents (Tomlinson 1986). Due to natural impacts and human destruction, populations have been isolated, fragmented, and highly disturbed. In China, L. littorea is an endangered species, distributed in very restricted regions of Hainan province.
More than 8,000 tree species currently are threatened with extinction worldwide (Oldfield et al. 1998). However, relatively few genetic investigations have been undertaken on these species and for the vast majority of them, information on patterns of intraspecific genetic variation is entirely lacking (Allnutt et al. 2003). In recent years, genetic data have being used to successfully define units for conservation management and for inferring changes in population structure and dynamics in plants (Moritz 1995). It is widely appreciated that genetic analyses can provide valuable insights into processes influencing extinction (Clarke and Young 2000). A thorough understanding of genetic diversity is an important aspect of conservation strategies for threatened taxa because of their unusual genetic and life history features (Holsinger and Gottlieb 1991). Therefore, a genetic study of L. littorea should provide valuable data to evaluate the outcome of historical events such as range expansion or population fragmentation and bottlenecks, which may have significant implications for the development of conservation strategies (Allnutt et al. 2003; Newton et al. 1999).
The use of molecular markers has become routine for studying population genetic diversity in plants (Zietkiewicz et al. 1994; Wolfe and Liston 1998; Ge and Sun 1999; Nan et al. 2003). Inter-simple sequence repeat (ISSR) markers are particularly effective in the study of genetic variation, because they overcome some of the limitations of other common markers such as allozymes, random amplified polymorphic DNA (RAPD), and restriction fragment length polymorphism (RFLP). Long primers are used in the ISSR technique that allow for higher annealing temperatures, a factor resulting in greater reproducibility of the bands (Wolfe and Liston 1998). Because high levels of polymorphism typically are detected, ISSRs have been employed successfully in genetic studies not only of economically important cultivated and wild plants, but also some mangrove species (Ge and Sun 1999, 2001; Jian et al. 2004; Tan et al. 2005).
In the present study, ISSR markers were applied to elucidate genetic diversity and genetic structure within and among five natural populations of L. littorea from South China, Malay Peninsula, Sri Lanka, and North Australia. The major objectives were: (a) to estimate the level of genetic variation in L. littorea and assess the efficiency of ISSR markers for studying this species; (b) to detect whether genetic differentiation has occurred among populations and identify factors that influence population genetic structure in this species; (c) to elucidate factors contributing to the imperilment of the species and provide information that may be useful for developing future conservation and breeding programs for this endangered species.
Materials and methods
Plant sampling
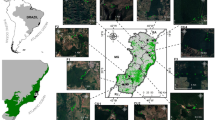
Leaf samples of L. littorea were collected with a total of 82 individuals from five natural populations in the Indo West Pacific range (Table 1; Fig. 1). Because of the fragmentation and rarity of L. littorea, these were the only populations we could locate, and all the individual trees that we could reach were sampled in each population, except for the biggest population in Lu-un, west of Thailand (see Table 1 and Fig. 1) where relatively dense stands of individual species were found. Leaf materials were stored with silica gel in zip-lock plastic bags until DNA isolation.

Map showing location of Lumnitzera littorea populations sampled. Population names are SC Dongzhaigang, Hainan, China ; EM Khanom, Thailand; WM Lu-un, Thailand; SL Balapitiya, Sri Lanka; NA Daintree River, Australia
DNA isolation and PCR amplification
Genomic DNA was isolated using the modified CTAB method (Doyle and Doyle 1987). After quantification with a Hoeffer fluorometer, a working sample solution, with concentration of 20 ng/μl of genomic DNA, was prepared.
One hundred ISSR primers from the Biotechnology Laboratory, University of British Columbia (UBC set no. 9) were initially screened and twelve of the primers that produced clear and reproducible fragments were used: 808 (AG)8C, 823 (TC)8C, 836 (AG)8YA, 840 (GA)8YT, 841 (GA)8YC, 842 (GA)8YG, 844 (CT)8RC, 845 (CT)8RG, 868 (GAA)6, 873 (GACA)4, 874 (CCCT)4, 876 (GATA)2(GACA)2.
Polymerase chain reaction (PCR) amplifications were performed in each 10 μl reaction, consisting of 10 mM Tris-HCl, 50 mM KCl, 1.5 mM MgCl2, 0.1 mM dNTPs, 0.1% Triton X-100, 2% formamide, 200 nM primer, 1.5 unit of Taq polymerase, and 20 ng of genomic DNA. Amplifications were performed using a PTC-100 thermocycler and the following profile: initial denaturation at 94°C for 5 min followed by 40 cycles of 1 min at 94°C, 1 min 5 s annealing at 52°C, and 2 min extension at 72°C; lastly a final extension of 7 min at 72°C. Amplification products were electrophoresed on 1.5% agarose gels buffered with 0.5× TBE, and detected by staining with ethidium bromide. A Gene Ruler 100 bp DNA ladder plus (Shengong Inc., Shanghai, China) was used to estimate the sizes of ISSR fragments. Furthermore, we had repeated amplifications to verify the integrity of the reproducible bands scored.
Data analysis
ISSR bands were scored as present (1) or absent (0) for each DNA sample excluding the smeared and weak ones. The binary data matrix was input into POPGENE (Yeh et al. 1997), assuming Hardy-Weinberg equilibrium. The following indices were used to quantify the amount of genetic variation within each population examined: the percentage of polymorphic loci (P), the mean expected heterozygosity (He) (Nei 1973), effective number of alleles (Ne), and Shannon’s information index of diversity (I). Genetic variation parameters (P, He, Ne and I) also were calculated at the species level.
Genetic differentiation among populations was estimated by Nei’s gene diversity statistics (Nei 1973). The amount of gene flow among these populations was estimated as Nm = (1/Gst-1)/4 (Slatkin and Barton 1989). Pair wise genetic distances between populations (Nei 1972) were calculated and used to construct a UPGMA dendrogram.
Analysis of molecular variance (AMOVA) was conducted to calculate variance components and their significance levels for variation among populations, and within populations using AMOVA version 1.55 (Excoffier 1993). The input files for AMOVA were prepared using the program AMOVA-PREP version 1.01 (Miller 1998).
Results
Genetic variability
Across all 82 L. littorea individuals from the five populations surveyed, a total of 221 reproducible bands were generated from the 12 ISSR primers evaluated. Of the 221 loci surveyed, 178 were polymorphic in at least one of the sampled populations. Sizes of the ISSR fragments varied from 200 to 2,000 bp.
At the species level, the percentage of polymorphic loci (P), mean expected heterozygosity (He), effective number of alleles (Ne) and Shannon’s index (I) were 75.57, 0.240, 1.402, and 0.365%, respectively (Table 1) (data coming from two individuals of SL population was inadequate to compare with all the other populations about population genetic variability but could be used for assessing relationships among populations, i.e., the ones synthesized in the dendrogram). At the population level, the value of P per population varied from 28.96% (North Australia) to 45.07% (Hainan, China), with an average of 37.06% (Table 1). The mean values of He, Ne and I were 0.118, 1.196, and 0.179. The highest level of expected heterozygosity existed in the Hainan, China population (He = 0.158); whereas, the lowest was that from North Australia (He = 0.098) (Table 1).
Population structure
The data showed that total gene diversity (Ht = 0.243 ± 0.035) was primarily distributed between populations (Dst = 0.125 ± 0.021). The value of coefficient of gene differentiation (Gst) was 0.515, indicating that a relatively high level of genetic differentiation existed among the five populations. Based on the Gst value, the estimated number of migrants per generation (Nm) was 0.470 (Table 2).
South China Sea is a semi-enclosed tropic sea, isolated by the Malay Peninsula and Indonesia archipelago. Favorable currents enable fruits of L. littorea to disperse over considerable distances, resulting in effective gene flow in this region (Zhang et al. 2005). Therefore, population SC and EM from South China Sea were analyzed together, separated from other populations. Values of P, Ht, Gst, and Nm were 54.75%, 0.171 ± 0.035, 0.227, and 1.708, respectively (Table 2).
Analysis of pair wise ISSR distances by AMOVA indicated that more than half the total variation (58.36%) was accounted for by differentiation among populations, with the remainder (41.64%) partitioned among individuals within populations (Table 3), suggesting that significant genetic structure exists among populations of L. littorea. When individual pairs of populations were compared, pair-wise phiST values derived from AMOVA also highlighted a large number of differences between populations, except between population SC and EM (Table 4). All components of molecular variance were highly significant (P < 0.001).
Relationships among geographical regions
The mean value of Nei’s genetic distance as calculated by POPGENE was 0.2804 among populations. A UPGMA dendrogram constructed from genetic distances indicated that populations from South China Sea (SC and EM) clustered as an inner group, which clustered next with population from North Australia (NA), with the population from West Malaya (WM) as the outlier. The dendrogram showed that individuals of each population formed a cluster, respectively (Fig. 2).

UPGMA dendrogram of Lumnitzera littorea populations based on ISSR markers (see Fig. 1 for population locations). SC South China; EM East Malaya; WM West Malaya; SL Sri Lanka; NA North Australia
Discussion
The present study is the first report of genetic diversity within and among natural populations of L. littorea using ISSR markers. The detection of high levels of polymorphism detected in L. littorea makes ISSR analysis a powerful tool for assessing genetic diversity in this species.
Genetic variation
At the species level, the present indices showed a relatively high level of genetic variation (P = 75.57%, He = 0.240, Ne = 1.402) in L. littorea. Compared with other studied mangroves, our results were similar to those reported for Heritiera littoralis (P = 91.69%, He = 0.236), another non-viviparous mangrove (Jian et al. 2004), and for Ceriops decandra (P = 72.46%, He = 0.256), a viviparous species (Tan et al. 2005), also studied using ISSR markers.
As Hamrick et al. (1991) suggested several factors are important in determining levels of genetic diversity. For instance, geographic range is strongly associated with the level of variation maintained at the species level. Widely distributed plant species tend to maintain more variation than more narrowly distributed species. Although L. littorea in China is restricted to Hainan, the species covers a wide geographic range throughout the Indo-Pacific region. Other factors such as breeding systems, vegetative reproduction, etc., also significantly influence the genetic diversity of a species (Hamrick et al. 1991).
At the population level, genetic variation was low in L. littorea in contrast to the high variation detected at the species level. This result was expected as these populations may have been subjected to repeated bottleneck or founder events in earlier times, due to episodes of glaciations and transgressions. Successive glaciations during the Pleistocene caused the lowering and rising of sea levels, which resulted in contractions and expansion of mangroves during glacial and interglacial periods, respectively (Saenger 1998). On the other hand, Huang et al. (1994) suggested that low genetic variation within populations is an ecological consequence of high habitat homogenization. Because mangrove species constantly are subjected to physiological stress caused by unstable growing conditions, their successful colonization to suitable areas might be associated with morphological, physiological, and reproductive adaptations (Lakshmi et al. 1997). Therefore, there is lesser population genetic variation caused by edaphic preferences and adaptations in mangroves.
Genetic structure
The present distribution of genetic structure within a species is influenced both by its natural and evolutionary history (Hamrick and Godt 1996). The coefficient of gene differentiation (Gst) was 0.515, under the assumption of Hardy-Weinberg equilibrium. Moreover, AMOVA also showed that more than half of the total variation (58.36%) was accounted for by differentiation among populations. Therefore, both genetic and phenetic analytical approaches indicated significant genetic structure among L. littorea populations.
This result may reflect the effects of biogeographic history. Palynological and glaciological evidence indicate that during the last glaciation, Sumatra, Borneo, and the Malay Peninsula were connected to mainland Asia; whereas, the Indian Ocean populations were isolated from those in South China Sea (Brown and Lomolino 1998). Much of the coastal and estuarine habitat was covered by an ice-sheet, and mangrove forests were greatly reduced in extent compared to current distribution. Mangroves may have survived only in some refugia. Thus, differentiation would have been likely to occur between the relatively small isolated populations as a consequence of increased inbreeding and genetic drift.
The low value of gene flow (Nm = 0.470) estimated among populations and the well-defined clusters that appear in the genetic distance dendrogram (Fig. 2) revealed that geographical isolation might be another factor influencing genetic structure and gene flow in L. littorea. Lumnitzera littorea is an out crossing non-viviparous species, and successful fruit dispersal relies on suitable ocean currents (Tomlinson 1986). Although mature diasporas of non-viviparous mangroves generally cannot anchor in the sediment immediately after falling from parent trees and are subject to tidal flushing (Ye et al. 2004), fruits of L. littorea cannot disperse far without suitable ocean currents. The Andaman Sea is connected with South China Sea only through the Strait of Malacca, and the Indonesia archipelago is a porous barrier to gene flow. Therefore, genetic exchange and gene flow among South China Sea, Andaman Sea, and North Australia may be very restricted by the isolation of Malay Peninsula and Indonesia archipelago. Strong and significant differentiation was also found in many mangroves and marine species between the West Pacific and Indian Ocean (Ge and Sun 2001; Yamazaki 1998; Benzie 1999).
However, gene exchange can occur with suitable ocean currents. For example, in the region of South China Sea, ocean currents are determined by the Northeast (November–March) and the Southwest (May–September) Monsoons, which change direction twice per year. The high value of gene flow (1.708) and low Gst (0.227) between population SC and EM in this region suggested that gene flow might have occurred between these two populations. A similar conclusion was reached for Excoecaria agallocha, another non-viviparous mangrove (Zhang et al. 2005).
Decline and conservation of L. littorea
During recent years, great loss of L. littorea has been caused not only by natural impacts, but also by human destruction. For example, L. littorea was recorded 45 years ago for the first time by Abeywickrema (1960) and has now become a very rare species in Sri Lanka (Jayatissa et al. 2002). The unique geographical environment makes mangrove ecosystems highly fragile. The habitats have been subjected to long duration of intrusion as well as incessant physiological and morphological stresses due to salinity aeration and onslaught of wave action (Mongia et al. 1991). In addition, natural calamities, such as cyclone, storm and flood, often cause extensive damage to mangrove areas (Koteswaran 1984). Besides natural impacts, mangroves are prone to man-made threats, such as gazing by cattle/goat/camel, overexploitation of juvenile fishes, felling for timber and firewood, human inhabitation and pollution etc., (Chaudhuri and Choudhurg 1994; Banerjee et al. 1998). Therefore, L. littorea tends to occur in small, isolated population, and becomes more and more endangered in many regions. For instance, L. littorea can only survive in the very limited areas in the nature reserve in Hainan Island, South China.
It is widely appreciated that fragmentation and isolation of populations can result in reduced reproduction or survival and thereby reduce population viability (Allnutt et al. 2003; Shrewin and Moritz 2000). Accurate estimates of genetic structure are very useful for conserving and managing tree genetic resources (Hamrick et al. 1991). Genetic information obtained from this study should help provide a clear framework for the development of conservation program for L. littorea. The extensive population genetic divergence among populations detected in L. littorea indicated that each population should be conserved separately. Populations that are characterized by a particularly high differentiation could be considered as conservation priorities. For example, the population SL with genetically highest distinction of those considered here is particularly vulnerable, because of its very small size and extreme isolation.
The choice of populations as sources of seeds for the restoration of threatened populations should depend, in part, on genetic similarity of donor populations to those in need of restoration (Hamrick and Godt 1996). As populations of L. littorea maintain a high degree of genetic differentiation except between population SC and EM, it is not recommended to try any transplantation because this could lead to out breeding depression and reduced fitness (Ellstrand and Elam 1993). On the contrary, due to genetic similarity and higher gene flow between population SC and EM (Gst = 0.227, Nm = 1.708) (Table 2), transfer of germplasms between these two places could be tried to increase their probability of survival. On the other hand, if the purpose is to replace the lost populations at certain locations, then sampling strategies should be focused on transplanting a large number of seeds from long-established, large populations to obtain the maximum representation of genetic adaptability.
Lumnitzera littorea tends to occur in small, isolated populations in many part of its range. Small populations are at risk of losing genetic flexibility by drift and short-term lowering of fitness resulting from inbreeding depression (Allnutt et al. 2003; Shrewin and Moritz 2000). Furthermore, the Allee effect states that if numbers in a given population are too low, the population will continuously decline in numbers and fitness, and is increasingly likely to go to extinction (Aaron and John 2003; Stephens and Sutherland 1999). This suggests that small L. littorea populations will go to extinction once the number of individuals drops below a critical threshold. Therefore, it is most important to monitor those small, isolated populations, maintaining their population sizes and fitness.
References
Aaron R, John SP (2003) Conservation of the critically endangered Rumex rothschildianus as implied from AFLP diversity. Biol Conserv 114(2003):299–303
Abeywickrema BA (1960) Estuarine vegetation of Ceylon. Proceedings of the Abidjan symposium on the humid tropics. UNESCO, Paris, pp 207–210
Allnutt T, Newton AC, Premoli A, Lara A (2003) Genetic variation in the threatened South American conifer Pilgerodendron uviferum (Cupressaceae), detected using RAPD markers. Biol Conserv 114(2003):245–253
Banerjee LK, Ghosh D, Sastry ARK (1998) Mangroves, associates and salt marshes of the Godavari and Krishna Delta. Bot Surv India, Kolkata, p 128
Benzie JAH (1999) Major genetic differences between crown-of-thorns starfish (Acanthaster planci) populations in the Indian and Pacific Ocean. Evolution 53:1782–1795
Brown JH, Lomolino MV (1998) Biogeography. 2 edn. Sinauer Associates, Sunderland
Chaudhuri AB, Choudhury A (1994) Mangroves of the Sundarbans. Vol 1. India, p 247
Clarke GM, Young AG (2000) Introduction: genetics, demography and the conservation of fragmented populations. In: Young AG, Clarke GM (eds) Genetics, demography and viability of fragmented population. Cambridge University Press, Cambridge, pp 1–6
Doyle JJ, Doyle JL (1987) A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull 19:11–15
Ellstrand NC, Elam DR (1993) Population genetic consequences of small population size: implications for plant conservation. Annu rev Ecol Syst 24:217–242
Excoffier L (1993) Analysis of molecular variance. Version 1.55. Genetics and Biometry Laboratory, University of Geneva
Ge X.J, Sun M (1999) Reproductive biology and genetic diversity of a cryptoviviparous mangrove Aegiceras corniculatum (Myrsinaceae) using allozyme and inter-simple sequence repeat (ISSR) analysis. Mol Ecol 8:2061–2069
Ge XJ, Sun M (2001) Population genetic structure of Ceriops tagal (Rhizophoraceae) in Thailand and China. Wetlands Ecol Manag 9:203–209
Hamrick JL, Godt MJW, Murawski DA, Loveless MD (1991) Correlations between species and allozyme diversity: implications for conservation biology. In: Falk DA, Holsinger KE (eds) Genetics and Conservation of Rare Plants. Oxford University Press, New York, pp 75–86
Hamrick JL, Godt MJW (1996) Conservation genetics of endemic plant species. In: Avise JC, Hamrick JL (eds) Conservation Genetics. Chapman & Hall, New York, pp 281–304
Holsinger KE, Gottlieb LD (1991) Conservation of rare and endangered plants: principles and prospects. In: Falk DA., Holsinger KE (eds) Genetics and Conservation of Rare Plants. Oxford University Press, Oxford, pp 195–223
Huang H, Dane F, Norton JD (1994) Allozyme diversity in Chinese, Seguin and American chestnut (Castanea spp.). Theor Appl Genet 88:981–985
Jayatissa LP, Dahdouh G, Koedam N (2002) A review of the floral composition and distribution of mangroves in Sri Lanka. Bot J Linnean Soc 2002 138:29–43
Jian SG, Tang T, Zhong Y, Shi SH (2004) Variation in inter-simple sequence repeat (ISSR) in mangrove and non-mangrove populations of Heritiera littoralis (Sterculiaceae) from China and Australia. Aquat Bot
Koteswaran P (1984) Climate and mangrove forests. Report of the second introductory training course on mangrove ecosystems. Sponsored by UNPD and United Nations Educational, Scientific and Cultural Organisation, Goa, India, pp 29–46
Lakshmi M, Rajalakshmi S, Parani M, Anuratha CS, Parida A (1997) Molecular phylogeny of mangroves: I. Use of molecular markers in accessing the intraspecific genetic variability in the mangrove species Acanthus ilicifolius Linn. (Acanthaceae). Theor Appl Genet 94:1121–1127
Miller MP (1998) AMOVA-PREP 1.01: a program for the preparation of the AMOVA input files from dominant-marker raw data. Department of Biological Sciences, Northern Arizona University, Flagstaff, AZ
Mongia AD, Bandyopadhyay AK, Dagar JC (1991) Mangroves of Andaman and Nicobar Islands
Moritz C (1995) Uses of molecular phylogenies for conservation. Philos Trans R Soc Lond B Biol Sci 349:113–118
Nan P, Shi S, Peng S, Tian C, Zhong Y (2003) Genetic diversity in Primula obconica from Central and Southwest China as revealed by ISSR markers. Ann Bot 91:329–333
Nei M (1972) Genetic distance between populations. Am Nat 106:283–282
Nei M (1973) Analysis of gene diversity in subdivided populations. Proc Natl Acad Sci USA 70:3321–3323
Newton AC, Allnutt T, Gillies ACM, Lowe A, Ennos RA (1999) Molecular phylogeography, intraspecific variation and the conservation of tree species. Trends Ecol Evol 14(4):140–145
Oldfield S, Lusty C, MacKinven A (1998) The world list of threatened trees. World Conservation Press, WCMC, Cambridge
Saenger P (1998) Mangrove vegetation: an evolutionary perspective. Mar Freshw Res 4:277–286
Sherwin WB, Moritz C (2000) Managing and monitoring genetic erosion. In: Young AG, Clarke G.M (eds) Genetics, demography and viability of fragmented populations. Cambridge University Press, Cambridge, pp 9–34
Slatkin M, Barton NH (1989) A comparison of three indirect methods for estimating average levels of gene flow. Evolution 43:1349–1368
Stephens PA, Sutherland WJ (1999) Consequences of the Allee effect for behaviour, ecology and conservation. Trends Ecol Conserv 14:401–405
Tan FX, Huang YL, Ge XJ, Su GH, Ni XW, Shi SH (2005) Population genetic structure and conservation implications of Ceriops decandra in Malay Peninsula and North Australia. Aquat Bot 81(2):175–188
Tomlinson PB (1986) The Botany of mangroves. Cambridge University Press, Cambridge, pp 374–381
Wolfe AD, Liston A (1998) Contributions of PCR-based methods to plant systematics and evolutionary biology. In: Soltis DE, Soltis PS, Doyle JJ (eds) Molecular systematics of plants. II.DNA sequencing. Chapman & Hall, New York, pp 43–86
Yamazaki T (1998) Molecular polymorphism and population structure of several tropical tree species in Southeast Asia. Amer. J. Bot. 85, 62 (Abstract)
Ye Y, Lu CY, Wong YS, Tam NFY (2004) Diaspore traits and inter-tidal zonation of non-viviparous mangrove species. Acta Bot Sin 46(8):896–906
Yeh FC, Yang RC, Boyle TBJ, Ye ZH, Mao JX (1997) POPGENE, the user-friendly shareware for population genetic analysis. Mol. Biol. Biotech. Center, University of Alberta, Canada
Zhang ZH, Tang T, Zhou RC, Wang YG, Jian SG, Zhong CR, Shi SH (2005) Effects of divergent habitat on genetic structure of population of Excoecaria agallocha, a mangrove associate. Acta Genet Sin, In press
Zietkiewicz E, Rafalski A, Labuda D (1994) Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics 20:176–783
Acknowledgments
We thank Dr. Norman Duke, Dr. Sonjai Havanond, Dr. Xuejun Ge, Mr. Cairong Zhong, and Dr. L.P. Jayatissa for their assistance in the collection of plant material. This study was supported by grants from the National Natural Science Foundation of China (30070053, 30230030, 30300033, 30470119, 30500049), the Natural Science Foundation of Guangdong Province (001223, 5003316), International Foundation for Sciences (D/3836-1), the Start-up Research Funds for Young Teachers from Sun Yat-Sen University, the Chung Hungta Science Foundation of Sun Yat-sen University, the Qiu Shi Science and Technology Foundation and the National Key Project for Basic Research (973) grant 2003CB715904.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by O. Kinne, Oldendorf/Luhe
Rights and permissions
About this article
Cite this article
Su, G., Huang, Y., Tan, F. et al. Conservation genetics of Lumnitzera littorea (Combretaceae), an endangered mangrove, from the Indo-West Pacific. Mar Biol 150, 321–328 (2007). https://doi.org/10.1007/s00227-006-0357-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-006-0357-6