
Abstract
There is a concern that the combination of exercise with food intake reduction has a risk of reducing bone strength and bone mass in young female athletes. We examined the influence of the interaction of voluntary running exercise and food restriction on bone in young female rats. Seven-week-old female Sprague–Dawley rats were divided into four groups: the sedentary and ad libitum feeding group (SED), voluntary running exercise and ad libitum feeding group (EX), sedentary and 30 % food restriction group (SED-FR), and voluntary running exercise and 30 % food restriction group (EX-FR). The experiment lasted 12 weeks. Statistical analysis was carried out by two-way analysis of variance with exercise and restriction as the between-subjects factors. As a result, there were significant interactions of running and restriction on energy availability, breaking force, breaking energy, and bone mineral density (BMD). Breaking force and energy in the EX group were significantly higher than in the SED group; breaking force and energy were significantly lower in the EX-FR group than in the EX group, and breaking force in the EX-FR group was significantly lower than that in the SED-FR group. BMD in the EX-FR group was significantly lower than in the EX and SED-FR groups. These results suggest that food restriction induced low bone strength in young female rats engaging in voluntary running exercise. Also, through the interaction of exercise and food restriction, voluntary running exercise combined food restriction, unlike ad libitum feeding conditions, induced low bone strength, and low BMD in young female rats.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
There is a concern that the combination of exercise and food intake reduction has a risk of reducing bone strength and bone mass in young female athletes [1, 2]. It is generally recognized that exercise increases bone strength by increasing mechanical loading [3–5]. For exercise to exert an anabolic effect on bones, adequate nutritional status is essential [6]. It has been reported that females who exercise with an inadequate energy intake can suffer low bone mineral density (BMD) due to a reduction in energy availability, which is the amount of dietary energy remaining for other body functions after exercise training, as well as menstrual dysfunction [1]. Low bone strength, including that caused by the combination of exercise and food intake reduction, may cause stress fractures [7, 8], is an important problem to address.
Recently, low bone strength and bone mass resulting from the combination of running exercise and food intake reduction in females has been studied using mature female rats [9–11]. Previous studies have reported that food restriction in sedentary female rats induces low BMD [12–14]. Furthermore, it has been reported that food restriction in female rats undergoing an exercise regimen induced low bone mineral content (BMC) [9, 10] and BMD [11]. However, the influence of food restriction on bone in “young” female rats engaging in running exercise is unclear. Research on bone during the adolescent years has been regarded as important [2] because the adolescent period is critical for acquiring bone mass [15]. Therefore, it is essential to study this topic in an adolescent model such as young female rats.
Additionally, it is necessary to reveal the influence of running exercise on bone in female rats under food restriction conditions. In previous studies, it has been reported that running exercise under ad libitum feeding conditions in female rats induced high bone mass [16] or high BMD [3, 17]. However, the influence of running exercise on bone in female rats has not been reported under food restriction conditions. Engaging in running exercise in combination with low energy availability due to food restriction may result in lower bone strength than a sedentary lifestyle with food restriction because running exercise further decreases energy availability.
Accordingly, the aim of the present study was to examine the influence of the interaction of voluntary running exercise and food restriction on bone in young female rats.
Materials and Methods
Experimental Design
Female Sprague–Dawley rats (n = 32, 7 weeks old) were randomly divided into four experimental groups after a 1-week acclimatization period. The groups included a sedentary and ad libitum feeding group (SED, n = 8), a voluntary running exercise and ad libitum feeding group (EX, n = 8), a sedentary and 30 % food-restricted group (SED-FR, n = 8), and a voluntary running exercise and 30 % food-restricted group (EX-FR, n = 8). The experiment period was 12 weeks. The rats were purchased from CLEA Japan (Tokyo, Japan) and were fed the diet described in Table 1. The SED and SED-FR groups were housed individually in normal cages (15 × 25 × 19.5 cm), while the EX and EX-FR groups were housed individually with free access to permit wheel cage to voluntary running exercise (circumference 1 m; 27 × 35 × 35 cm). We used voluntary running so that the changing daily running distance could be assessed and to ensure momentum. The SED-FR and EX-FR groups were fed a 30 % restricted diet that was calculated to contain 70 % of the mean amount consumed in the previous week by the SED group. As a result, the EX-FR group was fed a diet with a mean 35 % reduction compared to the Ex group. The room was maintained at 22 ± 1 °C under a constant 12:12 h light–dark cycle (light 8:00–20:00).
Daily Data Collection and Specimen Harvesting
The body weight and amount of dietary intake were measured every second day, and the running distance was measured every day. On the day before the dissection, all rats fasted for 12 h. Whole blood samples were collected from the abdominal aorta using syringes under diethyl ether anesthesia. Serum samples were separated by centrifugation at 2500 rpm for 20 min at 4 °C. The serum was frozen at −80 °C for the determination of bone metabolic markers in the serum. The abdominal fat, uterus, adrenal gland, femurs, tibias, and lumbar spine were collected from each rat after death. Femurs were collected, freed from the adhering connective tissues, and their bone strength was immediately measured. The tibias and lumbar spines were stored in 70 % ethanol after being harvested and cleaned of soft tissue for measurement of the BMC, bone area, and BMD. The animals were placed in individual metabolic cages (24 × 20 × 18 cm3) on the 80th and 81st days, just before the end of the experimental period. Urine was collected over two 24-h periods. Urine was collected under acidic conditions using 2 ml 2N hydrochloric acid. All urine was centrifuged at 2500 rpm for 15 min to eliminate refuse, and tested by measuring bone turnover.
Calculation of Energy Availability
Energy intake was calculated by multiplying the amount of food intake in grams by the energy content of the food (3.73 kcal/g) (energy intake = food intake × 3.73 kcal/g). Exercise-induced energy expenditure due to daily wheel running was calculated by taking 5.0 kcal/kg body weight times the km run [18], as in a previous study [9] (energy expenditure = wheel-running distance × body weight × 5.0 kcal/kg body weight/km). Energy availability was calculated as energy intake minus exercise energy expenditure as previously described [9].
Evaluation of Estrous Cycle by Spectral Analysis of the Running Distance
This analysis was performed to examine whether the running distance increased every 4 or 5 days in order to assess the estrous cycle. To remove the slowly varying baseline from the data for the voluntary wheel-running distance, we used empirical mode decomposition (EMD) [19]. The EMD is a new adaptive data analysis method for analyzing nonlinear and non-stationary data. It decomposed the signal into several basic components called intrinsic mode functions (IMFs), and a residual understood as the signal trend.
First, we acquired the voluntary wheel-running distance date after 30 days (period which data were acquired was approximately 52 days), because estrous dysfunction due to food restriction with running exercise appears after 30 days according to a previous study [9]. Second, we analyzed the data using the EMD method and re-created the data set to extract the residual signal trend and the lowest-frequency IMF component from the original data. Third, we analyzed the data using the techniques of maximum entropy spectral analysis, using the final prediction error criterion for optimal order selection. Last, to exclude the inter-individual difference in the total power affecting the local power, we also computed the proportion of the power spectrum of the running distance from 0.2 to 0.3 Hz in the total power spectrum. We calculated the proportion of the power spectrum of the running distance from 0.2 to 0.3 Hz in the total power spectrum, because previous studies showed that female rats have a 4- or 5-day running cycle in association with the estrous cycle, and their running activity was observed to be high in proestrous [2, 9, 20]. If the running distance increased cyclically every 4 or 5 days, the proportion of the power spectrum of the running distance from 0.2 to 0.3 Hz in the total power spectrum was high. In contrast, wheel-running fluctuation in anestrous female rats was reported to be minimal [9]. So if female rats are anestrous, the proportion of the power spectrum of the running distance from 0.2 to 0.3 Hz in the total power spectrum is low.
Measurement of Bone Strength by Three-Point Bending Test
The strength of the femoral mid-shaft was assessed by a three-point bending test (DYN-1255, IIO DENKI, Tokyo, Japan) using previously described methods (distance between the fulcrums 1 cm; plunger speed 100 mm/min; full scale 50 kg; chart speed 120 cm/min) [21]. Breaking force refers to the loading weight (gravitational acceleration) required for bone breaking. Breaking energy refers to the workload that resulted in the breaking of the bone.
Measurement of BMC, Bone Area, and BMD by Dual-Energy X-ray Absorptiometry (DXA)
The BMC, bone area, and BMD values of the tibia and L3-L6 lumbar spines were measured by dual-energy X-ray absorptiometry (DXA; Aloka, DCS-600R, Tokyo, Japan) as previously described [22]. The tibia was divided into five divisions, and the first division from the upper side was considered to be the proximal metaphysis site. The second and third divisions from the upper side were considered to be the diaphysis site. The tibia at the metaphysis site contains mainly cancellous bone, and the tibia at the diaphysis site contains mainly cortical bone.
Bone Metabolic Markers
Serum bone-specific alkaline phosphatase (BAP) activity, a marker of bone formation, and tartrate-resistant acid phosphatase (TRAP) activity, a marker of bone resorption, were measured as previously reported [23]. The bone resorption marker deoxypyridinoline (Dpd, bone-specific type 1 collagen degradation product) was measured in the 24-h urine sample using commercially available kits (Metra Dpd EIA Kit, Quidel, San Diego, CA, USA). Urine calcium excretion (mg/d) was measured using the ICPS-8100 (Sequential Plasma Spectrometer, Shimadzu, Japan) according to the manufacturer’s protocol.
Statistical Analysis
All data were expressed as mean ± standard error (SE). Statistical analysis was carried out by two-way analysis of variance (ANOVA) with exercise (sedentary or exercise) and restriction (ad libitum feeding or food restriction) as between-subjects factors. In any analysis, if significant interactions were observed, the variables were analyzed with the Bonferroni multiple comparison test. Main effects were only considered when interactions were not significant. Running distance, exercise-induced energy expenditure, and the proportion of the power spectrum of the running distance from 0.2 to 0.3 Hz in the total power spectrum were analyzed using the Welch’s t test. The significance level was set at p < 0.05. All statistical analysis was performed using SPSS Statistical Packages (Ver. 19.0; SPSS Inc., Chicago, USA).
Results
Body Weight, Food Intake, Running Distance, Energy Availability, and Internal Organs Weight
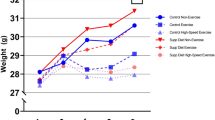
Figure 1 presents the results of the change in body weight, food intake, running distance, and energy availability during the experimental period. The body weight is expressed as the mean at the beginning of week. The food intake, running distance, and energy availability are expressed as the mean of the weekly average. The increase in body weight in the EX-FR group was suppressed. Food intake in the SED-FR and EX-FR groups continued to be restricted during the experimental period.

Changes in body weight (a), food intake (b), running distance (c), energy availability (d). Values are mean ± SE. However, the SE might not be noticeable in the figure if it was small. SED sedentary group, EX exercise group, SED-FR sedentary + food restriction group, and EX-FR exercise + food restriction group. Body weight, food intake, and energy availability were measured every other day, and running distance was measured every day. The value of body weight is expressed as the mean at the beginning of every week. The values of food intake, running distance, and energy availability are expressed as the mean of the weekly average for each parameter
Moreover, the body weight and internal organ weight at the dissection, the food intake, running distance, and related categories of energy availability are presented in Table 2. There was no significant interaction of exercise and restriction on abdominal fat weight, but there were significant main effects of exercise or restriction on causing lower abdominal fat weight.
There were significant interactions of exercise and restriction on average of food intake and energy intake. The average food intake and energy intake in the EX group were significantly higher than those in the SED group. The average food intake and energy intake in the SED-FR group were significantly lower than those in the SED group. The average food intake and energy intake in the EX-FR group were significantly lower than those in the EX group (Table 2). The average wheel-running distance and exercise-induced energy expenditure were not significantly different between the EX group and the EX-FR group. The average energy availability (kcal/day) levels of each group were as follows: SED; 66.3 ± 1.8, EX; 62.0 ± 1.4, SED-FR; 47.3 ± 0.1, and EX-FR; 36.2 ± 0.6. There was a significant interaction of exercise and restriction on average of energy availability. The average of energy availability in the SED-FR group was significantly lower than that in the SED group, and the average energy availability in the EX-FR group was significantly lower than that in the EX and SED-FR groups.
Reproductive Function
There was significant interaction of exercise and restriction on uterus weight (Table 2). The uterus weight in the SED-FR group was significantly higher than that in the SED group, and that in the EX-FR group was significantly lower than that in the EX and SED-FR groups. Additionally, the proportion of the power spectrum of the running distance from 0.2 to 0.3 Hz in the total power spectrum in the EX-FR group was significantly lower than that in the EX group. This result showed that rats in the EX-FR group showed estrous cycle abnormality.
Bone strength, BMD, BMC, and Bone Area
The breaking force and breaking energy of the femur are presented in Fig. 2. There were significant interactions of exercise and restriction on breaking force, and there were also significant interactions of exercise and restriction on breaking energy. The breaking force and breaking energy in the EX group were significantly higher than those in the SED group, while they were significantly lower in the SED-FR group than those in the SED group and significantly lower in the EX-FR group than those in the EX group. Moreover, the breaking force in the EX-FR group was significantly lower than that in the SED-FR group.

Breaking force and breaking energy of femur. a Breaking force of femur. b Breaking energy of femur. c Data were analyzed by two-way ANOVA. Values are expressed as mean ± SE. Data were analyzed by two-way ANOVA. SED sedentary group, EX exercise group, SED-FR sedentary + food restriction group, EX-FR exercise + food restriction group. If a result showed significant interaction (p < 0.05), the Bonferroni multiple comparison test was used to determine specific differences between means. # p < 0.05 for versus SED group, *p < 0.05 for versus EX group
The BMD of the lumbar spine, total tibia, proximal metaphysis tibia, and diaphysis tibia are presented in Fig. 3. There were significant interactions of exercise and restriction on the BMD of the lumbar spine, total tibia, proximal metaphysis tibia, and diaphysis tibia. Those in the EX-FR group were significantly lower than those in the EX and SED-FR groups.

Bone mineral density of lumbar spine and tibia spine. a BMD of lumbar spine. b BMD of total tibia. c BMD of proximal metaphysis tibia. d BMD of diaphysis tibia. e Data were analyzed by two-way ANOVA. Values are expressed as mean ± SE. Data were analyzed by two-way ANOVA. SED sedentary group, EX exercise group, SED-FR sedentary + food restriction group, and EX-FR exercise + food restriction group. If a result showed significant interaction (p < 0.05), the Bonferroni multiple comparison test was used to determine specific differences between means. # p < 0.05 for versus SED group, *p < 0.05 for versus. EX group, † p < 0.05 for versus SED-FR group
The BMC values and bone areas of the lumbar spine, total tibia, proximal metaphysis tibia, and diaphysis tibia are presented in Table 3. There were significant interactions of exercise and restriction on the BMC and bone area of the lumbar spine, total tibia, proximal metaphysis tibia and diaphysis tibia but not on the bone area of diaphysis tibia. The BMC and bone area of lumbar in the SED-FR group were significantly lower than those in the SED group, and those in the EX-FR group was significantly lower than those in the EX and SED-FR groups. The BMC of total tibia, proximal metaphysis tibia, and diaphysis tibia in the EX group were significantly higher in the SED group, and those in the EX-FR group was significantly lower than that in the EX and SED-FR groups. Additionally, the BMC of proximal metaphysis tibia in the SED-FR group was significantly lower than those in the SED group. The bone area of total tibia in the EX group was significantly higher than those in the SED group, while they were significantly lower in the SED-FR group than in the SED group and significantly lower in the EX-FR group than in the EX group. The bone area of diaphysis tibia in the EX group was significantly higher in the SED group, and those in the EX-FR group was significantly lower than that in the EX and SED-FR groups. Finally, food restriction had a significant main effect on decreasing the bone area of proximal metaphysis tibia.
Bone Metabolic Markers
Table 4 presents the results for serum BAP, serum TRAP, urine DPD, and urine calcium excretion. There were no significant interactions of exercise and restriction on serum BAP, serum TRAP, urine DPD, and urine calcium excretion. Exercise or food restriction had significant main effects on increasing BAP and TRAP, and these effects in the EX-FR group were higher than those in the other groups. Meanwhile, exercise had a significant main effect on causing higher DPD. There was no significant main effect of exercise or food restriction on urine calcium excretion.
Discussion
The aim of the present study was to examine the influence of the interaction of voluntary running exercise and food restriction on bone in young female rats. Our data demonstrated that the interaction of voluntary running exercise and food restriction resulted in lower energy availability, lower bone strength, or lower BMD than exercise alone or food restriction alone in young female rats.
In the present study, our data showed that the interaction of the combination led to lower energy availability compared with food restriction alone in female rats. Interaction of voluntary running exercise and food restriction induced lower energy availability compared with only exercise in female rats (Table 2), which was similar to previous study [9]. Energy availability is defined as energy intake minus exercise energy expenditure, and low energy availability causes suppression of the physiological system, including, cellular maintenance and growth [24], thus decreasing total energy expenditure [25]. Thus, low energy availability is a critical problem in female rats with that engage in exercise and undergo food restriction. We considered that the suppression of the physiological system due to low energy availability would occur in female rats as a result of the interaction of running exercise and food restriction in our study. As an indication of this physiological suppression, the increase in body weight in the EX-FR group was inhibited, and the uterus weight in the EX-FR group was significantly lower than that in the EX and SED-FR groups. It was reported that low uterus weight can be caused by estradiol deficiency [26]. Thus, in the present study, it may be that rats in the EX-FR group exhibited abnormal estradiol secretion due to low energy availability.
Moreover, we considered that reproductive dysfunction occurred in the EX-FR group due not only to the uterus weight but also to the proportion of the power spectrum of the running distance from 0.2 to 0.3 Hz in the total power spectrum. This proportion of the power spectrum of the running distance from 0.2 to 0.3 Hz in the total power spectrum in the EX-FR was low in the period from 30 to 82 days (Table 2). The disruption of the voluntary running distance peak in the EX-FR group appeared to be anestrous. Dimarco et al. [9] reported that anestrous female rats showed estradiol deficiency, so it possible that rats in the EX-FR group showed abnormal estradiol secretion.
Meanwhile, previous studies in the activity-stress paradigm reported that wheel-running distance was greatly increased in rats with food restriction [27, 28]. In contrast, in our study, although the wheel-running distance in the EX-FR group was 2.5 km/day longer than that in the EX group, this difference was not significant, and the result was similar to the results of previous studies using the same food restriction method [9, 11]. The difference in the extent of causing longer wheel-running distance results may have been caused by the difference in the amount of food restriction. The food intake of the EX-FR groups in the present study was 12.6 ± 0.0 g/day, whereas the food intake in the activity-stress paradigm was about 7 g/day or less [27]. We suggested that the food restriction in the present study was not strict enough to cause an extreme longer running distance.
The bone strength results showed that the voluntary running exercise led to high bone strength with ad libitum feeding conditions, and the interaction of voluntary running exercise and food restriction led to lower bone strength than the food restriction alone (Fig. 2). Our group reported that the combination of voluntary running exercise and food restriction resulted in low bone strength in young male rats [29], similar to finding in the present study with young female rats. Moreover, in the present study, the two-way ANOVA statistical methods, revealed that an interaction occurred as a result of combination of conditions. Although it is known that exercise causes higher bone strength with ad libitum feeding [20], the effect was the opposite with food restriction in young female rats. To the best of our knowledge, there has been no study that has reported the influence of the interaction of voluntary running exercise and food restriction on bone strength.
BMD is frequently used as a proxy measure for bone strength and accounts for approximately 70 % of bone strength [30]. BMD is the main factor that determines bone strength. The effect of voluntary running exercise on BMD has been reported to be positive [3, 20]. In contrast, the influence of food restriction on BMD has been reported to be negative in sedentary rats [12, 14] and in exercising rats [11]. In the present study, we revealed that there were interactions of voluntary running exercise and food restriction on the BMD of the lumbar spine and tibia; the BMD values of the lumbar spine and tibia in the EX-FR group were lower than those in the SED-FR group (Fig. 3). These results demonstrated that the interaction of the combination caused the negative influence of voluntary running exercise on BMD in young female rats. Dimarco et al. [9] reported that the BMD values of the femur and tibia were not significantly different between ad libitum intake and food restriction conditions in exercising female rats, a different result than that found in the present study. This difference might have been caused by the difference in the age of the rats. The age of the rat in the exercise and food restriction phase was lower in our study (8–20 weeks old) than in Dimarco et al.’s study (17–30 weeks old). Our findings could suggest that young female rats may be sensitive to the negative effects of the interaction of voluntary running exercise and food restriction. Meanwhile, Exercise did not have positive impact on BMD in rats fed ad libitum while bone strength in the EX group was higher than that in the SED group. Bone strength reflects not only BMD but also bone quality [30]. These results suggest that bone features beside BMD influence bone strength in our study. Additionally, our studies showed that the interaction of voluntary running exercise and food restriction resulted in low BMC and bone area in young female rats. The BMC of proximal metaphysis tibia was in tandem with the bone breaking force of femur, and the bone area of total tibia was in tandem with the bone breaking energy of femur. However, the BMC and bone area of diaphysis tibia, which is the same cortical bone site as the femur used for bone strength test, were not completely in tandem with the bone breaking force and bone breaking energy of femur. These results suggest both theories: Bone mass and bone size influence bone strength, and bone features other than those also influence bone strength in our study.
We consider that one of the factors of leading to the bone fragility resulting from the interaction of voluntary running exercise and food restriction is low energy availability. Low energy availability induces suppression of physiological systems [24], including the secretion of hormone, such as estradiol and IGF-1, which affect bone turnover and promote the strengthening of bone [31]. In the present study, it is possible that the rats in the EX-FR group exhibited abnormal estradiol secretion as suggested by the uterus weight and the proportion of the power spectrum of the running distance from 0.2 to 0.3 Hz in the total power spectrum. In support of this possibility, Dimarco et al. [9] and Yanaka et al. [11] reported that lower estradiol resulted from the combination of exercise and food restriction in mature female rats. Additionally, Barrack et al. [32] reported that adolescent female runners with elevated bone turnover showed low energy intake and low estradiol levels. Low energy availability may induce the elevation of bone turnover and low bone strength. In the present study, main effect of exercise or food restriction caused higher BAP and higher TRAP (Table 4). Further investigation of the alteration of hormone secretion, bone turnover, and the mechanism of bone fragility resulting from the combination of exercise and food restriction is necessary.
This study has the following limitation. With regard to the influence of food restriction, we focused on low energy availability because we considered it to be a critical problem in female rats that engage in exercise and are subject to food restriction. However, the food restriction involved not only energy restriction but also the reduction of the intake of all nutrients. Talbott et al. [33] reported that 40 % energy restriction and 80 % calcium restriction resulted in lower BMD than 40 % energy restriction alone in young female rats. Thus, it is possible that a reduction in nutrient intake, not just caloric intake, also impaired bone development.
In conclusion, the interaction of voluntary running exercise and food restriction resulted in lower energy availability, lower bone strength, and lower BMD than exercise alone and food restriction alone in young female rats. These results suggested that food restriction induced low bone strength in young female rats that engaged in voluntary running exercise. Additionally, unlike under ad libitum feeding conditions, the voluntary running exercise with food restriction led low bone strength and low BMD in young female rats due to the interaction of exercise and food restriction.
References
Nattiv A, Loucks AB, Manore MM, Sanborn CF, Sundgot-Borgen J, Warren MP, American College of Sports Medicine (2007) American College of Sports Medicine the female athlete triad. Med Sci Sports Exerc 39(10):1867–1882
Barrack MT, Ackerman KE, Gibbs JC (2013) Update on the female athlete triad. Curr Rev Musculoskelet Med 6(2):195–204
Newhall KM, Rodnick KJ, van der Meulen MC, Carter DR, Marcus R (1991) Effects of voluntary exercise on bone mineral content in rats. J Bone Miner Res 6(3):289–296
Hinda K, Burrowsb M (2007) Weight-bearing exercise and bone mineral accrual in children and adolescents: a review of controlled trials. Bone 40(1):14–27
Nichols DL, Sanborn CF, Essery EV (2007) Bone density and young athletic women. Sports Med 37(11):1001–1014
Borer KT (2005) Physical activity in the prevention and amelioration of osteoporosis in women. Sports Med 35(9):779–830
Lauder TD, Dixit S, Pezzin LE, Williams MV, Campbell CS, Davis GD (2000) The relation between stress fractures and bone mineral density: evidence from active-duty army women. Arch Phys Med Rehabil 81(1):73–79
Boutroy S, Van Rietbergen B, Sornay-Rendu E, Munoz F, Bouxsein ML, Delmas PD (2008) Finite element analysis based on in vivo HR-pQCT images of the distal radius is associated with wrist fracture in postmenopausal women. J Bone Miner Res 23(3):392–399
Dimarco NM, Dart L, Sanborn CB (2007) Modified activity-stress paradigm in an animal model of the female athlete triad. J Appl Physiol 103(5):1469–1478
Swift SN, Baek K, Swift JM, Bloomfield SA (2012) Restriction of dietary energy intake has a greater impact on bone integrity than does restriction of calcium in exercising female rats. J Nutr 142(6):1038–1045
Yanaka K, Higuchi M, Ishimi Y (2012) Effect of long-term voluntary exercise and energy restriction on bone mineral density in mature female rats. J Phys Fit Sports Med 1(4):695–702
Talbott SM, Cifuentes M, Dunn MG, Shapses SA (2001) Energy restriction reduces bone density and biomechanical properties in aged female rats. J Nutr 131(9):2382–2387
Mardon J, Trzeciakiewicz A, Habauzit V, Davicco MJ, Lebecque P, Mercier S, Tressol JC, Horcajada MN, Demigné C, Coxam V (2009) Dietary protein supplementation increases peak bone mass acquisition in energy-restricted growing rats. Pediatr Res 66(5):513–518
Hawkins J, Cifuentes M, Pleshko NL, Ambia-Sobhan H, Shapses SA (2010) Energy restriction is associated with lower bone mineral density of the tibia and femur in lean but not obese female rats. J Nutr 140(1):31–37
Weaver CM (2002) Adolescence: the period of dramatic bone growth. Endocrine 17(1):43–48
Holy X, Zérath E (2000) Bone mass increases in less than 4 wk of voluntary exercising in growing rats. Med Sci Sports Exerc 32(9):1562–1569
Shiga K, Hara H, Okano G, Ito M, Minami A, Tomita F (2003) Ingestion of difructose anhydride III and voluntary running exercise independently increase femoral and tibial bone mineral density and bone strength with increasing calcium absorption in rats. J Nutr 133(12):4207–4211
Anantharaman-Barr HG, Decombaz J (1989) The effect of wheel running and the estrous cycle on energy expenditure in female rats. Physiol Behav 46(2):259–263
Huang NE, Shen Zheng, Long SR, Wu MC, Shih HH, Zheng Q, Yen NC, Tung CC, Liu HH (1998) The empirical mode decomposition and the Hilbert spectrum for nonlinear and non-stationary time series analysis. Proc R Soc Lond A 454(1971):903–995
Wang G (1923) The relation between “spontaneous” activity and oestrous cycle in the white rat. Comp Psychol Monogr 2:1–27
Ezawa I, Okada R, Nozaki Y, Ogata E (1979) Breaking-properties and ash contents of the femur of growing rat fed a low calcium diet. Nippon Eiyo Shokuryo Gakkaishi. J Jpn Soc Food Nutr 32(5):329–335
Omi N, Morikawa N, Ezawa I (1994) The effect of voluntary exercise on bone mineral density and skeletal muscles in the rat model at ovariectomized and sham stages. Bone Miner 24(3):211–222
Omi N, Goseki M, Oida S, Sasaki S, Ezawa I (1994) The nutritional evaluation of globin on maintenance of bone metabolism in ovariectomized osteoporotic rats. J Nutr Sci Vitaminol 40(5):443–457
Loucks AB, Kiens B, Wright HH (2011) Energy availability in athletes. J Sports Sci 29(Suppl 1):S7–S15
Stubbs RJ, Hughes DA, Johnstone AM, Whybrow S, Horgan GW, King N, Blundell J (2004) Rate and extent of compensatory changes in energy intake and expenditure in response to altered exercise and diet composition in humans. Am J Physiol Regul Integr Comp Physiol 286(2):R350–R358
Erben RG, Brunner KS, Breig B (2004) Long-term sensitivity of uterus and hypothalamus/pituitary axis to 17 beta-estradiol is higher than that of bone in rats. J Bone Miner 19(11):1827–1832
Routtenberg A, Kuznesof AW (1967) Self-starvation of rats living in activity wheels on a restricted feeding schedule. J Comp Physiol Psychol 64(3):414–421
Woods DJ, Routtenberg A (1971) “Self-starvation” in activity wheels: developmental and chlorpromazine interactions. J Comp Physiol Psychol 76(1):84–93
Hattori S, Park JH, Agata U, Oda M, Higano M, Aikawa Y, Akimoto T, Nabekura Y, Yamato H, Ezawa I, Omi N (2014) Food restriction causes low bone strength and microarchitectural deterioration in exercised growing male rats. J Nutr Sci Vitaminol 60(1):35–42
NIH Consens Statement (2000) Osteoporosis prevention, diagnosis, and therapy. NIH Consens Statement 17(1):1–45
Ihle R, Loucks AB (2004) Dose-response relationships between energy availability and bone turnover in young exercising women. J Bone Miner Res 9(8):1231–1240
Barrack MT, Van Loan MD, Rauh MJ, Nichols JF (2010) Physiologic and behavioral indicators of energy deficiency in female adolescent runners with elevated bone turnover. Am J Clin Nutr 92(3):652–659
Talbott SM, Rothkopf MM, Shapses SA (1998) Dietary restriction of energy and calcium alters bone turnover and density in younger and older female rats. J Nutr 128(3):640–645
Acknowledgments
We thank Dr. Kumpei Tokuyama for his helpful comments on methods for evaluating the estrous cycle using spectral analysis of the running distance.
Conflict of interest
Yuki Aikawa, Umon Agata, Yuya Kakutani, Michito Higano, Satoshi Hattori, Hitomi Ogata, Ikuko Ezawa, and Naomi Omi declare that they have no conflict of interest.
Human and Animal Rights and Informed Consent
Animal care and experimental procedures were approved by the Animal Experimental Committee of the University of Tsukuba. We did not performed our study with human.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Aikawa, Y., Agata, U., Kakutani, Y. et al. The Interaction of Voluntary Running Exercise and Food Restriction Induces Low Bone Strength and Low Bone Mineral Density in Young Female Rats. Calcif Tissue Int 97, 90–99 (2015). https://doi.org/10.1007/s00223-015-0005-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00223-015-0005-6