
Abstract
Individuals with reading fluency difficulty (RFD) show an impairment in the simultaneous processing of multiple elements, which could be reflected in their visual attention span (VAS) capacity. However, the relationship between VAS impairment and RFD is still controversial. A series of processes underlie VAS, such as the early stage of visual attentional processing and the late stage of allocating and maintaining attentional resources. Therefore, the present study explored the relationships between VAS skills and RFD through the event-related potential (ERP) technique to disentangle the contributing cognitive processes regarding VAS from a temporal perspective. Eighteen Chinese adults with poor reading fluency and 18 age-matched normal readers participated. Their VAS skills were measured by a visual one-back task with symbols as nonverbal stimuli and key pressing as nonverbal responses, while relevant electrophysiological signals were recorded. The results showed that lower d’ values and abnormal electrophysiological activities (especially weak amplitudes in the N1 and P3 components) in the VAS task were observed for the nonfluent readers compared with the controls. These findings suggested that the low VAS capacity in adults with poor reading fluency could be reflected by problems both in directing selective attention to visually discriminate stimuli within a multielement string at the early processing stage and in allocating attention to further encode targets at the late processing stage. Alternative explanations were further discussed. The current results provide theoretical explanations of the VAS-RFD relationship from a temporal perspective and provide insights for future remediation of reading fluency difficulty.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Reading fluency is a critical reading skill that refers to reading rapidly and accurately with adequate comprehension (Langer et al. 2015; Wang and Zhang, 2021). However, some individuals exhibit reading fluency difficulty (RFD). Impairment in reading fluency can lead to severe academic, economic, and psychosocial consequences (González et al. 2015). Therefore, it is important to investigate the mechanism underlying RFD to further guide the design of specific interventions to improve reading fluency. Researchers have claimed that the process of fluent reading should be explored from the aspect of visual decoding, because an important aspect of fluent reading is the need to simultaneously process several orthographic units in a very short time frame, namely, visual rapid simultaneous processing (Langer et al. 2015). The visual attention span (VAS) is usually considered an index of visual rapid simultaneous processing (Bosse et al. 2007), which is defined as the number of distinct visual elements in a briefly presented array (e.g., a letter string) that can be processed in parallel (Bosse et al. 2007; Lobier et al. 2012a2013). Bosse et al. (2007) proposed the VAS-deficit hypothesis, suggesting the influence of VAS dysfunction on reading efficiency. In detail, difficulty in this visual attentional processing may limit the number of letters/characters in parallel processing, which would consequently hinder the orthographic decoding of multiletter/multicharacter units and in turn affect fluent reading procedures (Bosse et al. 2007; Valdois et al. 2003; Zhao et al. 2019).
The VAS-deficit assumption has been supported by accumulating evidence from empirical studies that adopted traditionally whole-/partial-report tasks with five-letter strings as stimuli (e.g., Bosse et al. 2007; Germano et al. 2014; Valdois et al. 2014, 2019; Zoubrinetzky et al. 2016) and modified VAS tasks, such as a flanked visual categorization task (Lobier et al. 2012a, b), a visual one-back task (Lallier et al. 2016; Zhao et al. 2019) and a perceptual matching task (Reilhac et al. 2013), with symbols as nonverbal stimuli and key pressing as a nonverbal response to measure VAS ability. The results indicated that dysfluent readers had lower scores in the VAS tasks than the control readers. Further position-based analysis revealed that individuals with RFD exhibited an abnormal pattern in their visual attentional distribution (Zhao et al. 2018). It has been suggested that typically developing children exhibited an inverted-U shape in their distribution pattern of VAS (Huang et al. 2019; Zhao et al. 2018), while children with RFD showed a W-shaped pattern of attention allocation in VAS-related tasks, which might be associated with interference from their notable crowding effect (Zhao et al. 2018).
Compared with these findings, several studies have failed to observe VAS deficits in RFDs. Relevant studies reported that German children and Hebrew adults with reading fluency impairment did not differ from unimpaired readers in VAS-related tasks (Banfi et al. 2018; Hawelka and Wimmer 2008; Yeari et al. 2017). Since orthographic transparency has been suggested to potentially modulate the VAS-reading relationship (Awadh et al. 2016; Lallier et al. 2014), the inconsistencies in the literature described above might be associated with differences in the orthographic depth of background languages. Studies in which a significant relationship was observed between the VAS and fluent reading were usually performed in the context of languages with deep orthography (e.g., French and Chinese), and studies showing opposite results in which no significant relationship was observed between VAS and reading fluency were mostly performed in languages with shallow orthography (e.g., German and Hebrew). In languages with shallow orthography, readers apply the rules of grapheme-to-phoneme correspondence (GPC), in which phonological regularity characterizes small orthographic units. In contrast, in languages with deep orthography, larger orthographic units are more regular than smaller ones (Ziegler and Goswami 2005). Based on relevant studies (Huang et al. 2019; Zhao et al. 2017), the VAS has been suggested to have a significant relation to the global lexical reading strategy in the context of languages with deep orthographies, especially in the later developmental stages (e.g., adults). However, most of the relevant literature in languages with deep orthography examined the VAS skills of RFD in developing readers, such as children and teenagers (Lobier et al. 2013; van den Boer and de Jong 2018; Zhao et al. 2018, 2019); thus, the VAS capacity of adults with RFD at late developmental stages in language with deep orthography is still unclear. Whether adults with RFD exhibit notable VAS impairments or show normal VAS ability as they accumulate reading experience and literature training warrants further investigation.
Furthermore, these conflicting results suggest that we should further explore the mechanisms underlying this visual attentional capacity to verify the relationship between VAS deficits and RFD. VAS has been implicated in a series of cognitive processes, such as the early perceptual analysis of a string’s basic visual feature, further visual discrimination processing across items within the string, and the transformation of visual representations of the multielement string into visual short-term memory that serves subsequent comparisons with the codes stored in episodic memory (Bosse et al. 2007). Hence, it is interesting to explore the stage(s) at which the potential mechanisms of the VAS deficit in dysfluent readers manifest. To address this issue, the time course of cognitive processes underlying the VAS was further examined by using the event-related potential (ERP) technique with high temporal resolution, which was necessary because it could provide fine temporal information for different processing stages that contribute to the VAS before a motor response.
To our knowledge, only one previous ERP study explored VAS-related processes in bilingual readers with normal reading abilities (Lallier et al. 2013). A visual one-back task was adopted in that study, and a five-letter string represented the probe stimulus and a following single letter represented the target stimulus. The study focused on the probe-locked P1 and N1 components and the target-locked P3 component. The P1 and N1 components at the parietal and occipital sites have been identified as electrophysiological correlates of early visual attentional processing (Eimer 2010; Luck 1995). Specifically, the P1 component is typically related to early perceptual analysis of the attended target, and its amplitude appears to be modulated by the visual properties (e.g., visual complexity) of stimuli (Johannes et al. 1995; Kang et al. 2016). The N1 component has been suggested to be sensitive to general visual discrimination processing (Vogel and Luck 2000) and is associated with the refinement of visual processes to relevant elements required to perform a task (Maurer et al. 2006). Stronger N1 amplitudes correspond to greater involvement of visual processing. The P3 component has been used to index the quality of the mapping procedure between the current stimuli and the mental representations stored in visual short-term memory (Lallier et al. 2013). Larger P3 amplitudes are elicited when a match is detected between previously encoded and stored information (e.g., the probe of a letter string) and an ongoing event (e.g., a target letter; Lallier et al. 2013). Therefore, the different processes potentially involved in VAS can be separately explored by these different ERP components, which would help clarify relevant mechanisms of VAS deficits in individuals with RFDs from a temporal perspective.
Therefore, the first aim of the present study was to investigate the VAS capacity of dysfluent readers in Chinese, a logographic language with deep orthography, and the focus was on individuals at late developmental stages (i.e., adult readers). Adults with normal reading abilities exhibit an inverted-U shape in the attentional distribution of multielement processing, suggesting a fixation advantage (Huang et al. 2019). Thus, the attentional distribution in adults with poor reading fluency will also be addressed here. The second aim of the present study was to investigate the underlying contributing processes of VAS skills in dysfluent readers with an ERP technique to disentangle fine processes from a temporal perspective to deepen our understanding of the relationship between VAS and reading fluency. Consistent with previous research (Lallier et al. 2013), we focused on string-locked P1 and N1 components that reflect the early processing of strings’ visual features and target-locked P3 components that reflect attentional allocation to the target item within the string. In particular, we expected that possible position effects related to attentional distribution patterns would be closely associated with electrophysiological activity in the P3 component.
Materials and methods
Participants
A total of 36 Chinese adults were enrolled in this study, including 18 adults with poor reading fluency (12 females) and 18 age-matched normal readers (hereinafter referred to as NRs; 12 females). We recruited the present participants by posters and online dissemination. Adults with RFD were screened by reading fluency tests at the single-character and sentence levels. Details of the psychometric screening tests are described below. Referring to previous research (Bertoni et al. 2019; Kuhl et al. 2020), individuals were classified with RFD if their score on at least one of the reading tests was − 1 SD below the average score of the normal readers. All participants were right-handed and had normal or corrected-to-normal vision without ophthalmologic or neurological abnormalities. Meanwhile, the verbal working memory storage capacity and nonverbal intelligence were balanced between groups and separately measured by a backward digit span test and participants’ scores on the math test in the college entrance examination. Detailed information on the two groups is presented in Table 1. All participants were rewarded for their participation, and written informed consent was obtained from the participants prior to the experiment. The research was approved by the Research Ethics Committee of School of Psychology, Capital Normal University. This study was carried out in accordance with The Code of Ethics of the World Medical Association (Declaration of Helsinki) for experiments involving humans.
Psychometric tests for identifying individuals with poor reading fluency
Reading fluency test at the single-character level
The reading fluency test at the single-character level was a character-list reading task. The split-half reliability was 0.93 (Zhao et al. 2017). A list of 400 Chinese characters intermixed with 13 noncharacters consisted of the testing items. Participants were required to read the items silently and occasionally to cross-out the noncharacters within the time limit of one minute. The noncharacters were used to ensure the effectiveness of silent reading. At the end of this test, participants were asked to mark the last item they read silently. The score consisted of the number of items read minus the number of errors. Specifically, errors were defined as noncharacters that were not identified as well as real characters that were incorrectly crossed out. The score unit was the number of characters read correctly in 1 min (c/min).
Reading fluency test at the sentence level
A sentence-verification task was used to measure reading fluency at the sentence level. The split-half reliability was 0.85 (Zhao et al. 2017, 2018). A total of 54 sentences were constructed (four for the practice session and the remaining 50 sentences for the formal test). All sentences pertained to simple facts, and the length of each sentence varied from 7 to 22 characters. Half of the sentences were true, and the other half were false. This test was presented using a Dell laptop and was programmed by E-prime 1.0. When a complete sentence appeared in the screen center, the participants were instructed to read the sentence silently as quickly and accurately as possible and to press the space bar once they had finished reading the sentence. After pressing the space bar, an assessment was followed in which the participants were required to press different keys to judge the veracity of the sentence. The accuracy of the veracity assessment was recorded to ensure the validity of the performance on the silent reading task. Because the mean accuracy was higher than 0.9 for almost all the participants, we did not include it in the following analyses. The reading speed for one sentence was calculated based on the relative ratio of the number of Chinese characters in the sentence to the time taken to read the sentence; in addition, the mean reading speed for the sentences that were judged correctly was computed and regarded as the final score of the sentence reading test. The unit of reading speed was the number of characters read in 1 min (c/min).
Visual attentional tasks
VAS Task
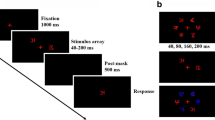
The stimuli in this test were ten symbols (Fig. 1a), and their visual complexity and familiarity were evaluated by another 35 undergraduates who did not participate in the formal experiment. The results of the rating scale with five points (1 = the symbol is not complex/familiar at all and 5 = the symbol is extremely complex/familiar) showed that the mean visual complexity was 2.30 and the mean visual familiarity was 2.36, thus revealing a middle level of visual complexity and familiarity for these items. The ratings of symbols were not significantly different between any two of these items (ps > 0.1), revealing the similarity and homogeneity across these stimuli. In this study, 300 5-figure strings were built from the 10 symbols, in which no string included the same symbol twice. Each symbol was used 150 times, with 30 times in each position. They were presented in black on a white screen. The visual angle of the strings was 7.9° × 0.8° at a distance of 65 cm. The center-to-center distance between two adjacent symbols was 1.7°.

Stimuli and presentation format of each trial in the visual one-back task and the control task. a All ten figures used in the visual one-back task and the control task are shown. b The procedure in the visual one-back task is presented. c The presentation format of each trial in the control task of single-figure recognition is displayed. d Each level of eccentricity in one string
A visual one-back paradigm was adopted here. In each trial (Fig. 1b), a 500-ms fixation was first presented in the screen center, and then a blank screen was shown for 100 ms, which was followed by a probe of the five-figure string centering on the screen for 200 ms. The string was replaced by a 100-ms mask with asterisks, and finally, a target of a single figure appeared below or above (half of the trials) the median horizontal line for a time window of 2000 ms. Participants were asked to press corresponding keys as quickly and accurately as possible when the target symbol appeared. Participants were asked to press the “Z” key if the target symbol was present in the above string and to press the “B” key if it was absent. The target symbol disappeared after the response or at the end of the time window 2000 ms. The interval between each trial randomly ranged from 200 to 500 ms. The formal experiment consisted of three blocks, with each block containing 100 trials. Half of the trials were for the “present” decisions, and the other half of the trials were for the “absent” decisions. The sequence of three blocks was balanced across participants. There were 20 trials for practice before the formal task. The reaction times and accuracy were recorded. Moreover, the d’ scores were computed based on accuracy in each position of the five-figure strings, which was a bias-free estimate of task sensitivity (Lallier et al. 2016; Pammer et al. 2004, 2005; Yeari et al. 2017). Because the d’ value and accuracy greatly overlapped, we finally utilized the bias-free estimate of d’ values as well as the reaction times in the following analyses.
Control task
A single-figure recognition task was used to assess whether the participants efficiently identified an individual symbol. The stimuli were the same ten symbols as mentioned above (Fig. 1a). The presentation duration and procedure for each trial (Fig. 1c) were the same with that used in the visual one-back task, except that the probe consisted of only one symbol rather than a five-symbol string. The participants were required to judge whether the probe and the target were the same. “Same” decisions were indicated by pressing the “Z” key, and “different” decisions were indicated by pressing the “B” key. The formal experiment consisted of 60 trials, with 30 trials for the “same” decisions (thrice for each figure) and 30 trials for the “different” decisions. There were ten practice trials before the formal experiment. The visual angle of the single symbol was 1.1° × 0.8°. Similarly, the reaction times and d’ values were the relevant measurements.
EEG recording and processing
EEG signals were recorded as the participants performed the visual attentional tasks. The participants were seated comfortably in a quiet, separated room. The procedures in the two visual attentional tasks were programmed by E-Prime 2.0 software and presented on a 21.5-inch liquid crystal display (LCD) monitor (1280 × 1024 pixels; 75 Hz frame rate) with a white background at a 65-cm viewing distance. The EEG signal was acquired using a 64-channel system (HydroCel Geodesic Sensor Net, Electrical Geodesics, Inc., Eugene, OR) with Net Station EEG software. The EEG signal was amplified with a bandpass of 0.1–30 Hz and digitized online at a sampling rate of 1000 Hz. The electrode impedance level was kept under 50 kΩ, which is an acceptable level for this system. All 64 electrodes were physically referenced to the average of a global field power reference (fixed by the EGI system) during EEG recordings.
Net Station EEG software was used to analyze the EEG data. The data were filtered off-line with a 30-Hz low-pass zero-phase shift digital filter and segmented into 1100-ms epochs that included a 100-ms prestimulus baseline and 1000-ms period after the onset of the string. ERPs containing artifacts (signal max–min > 200 µV), eye blinks (max–min > 140 µV in windows of 640 ms), and eye movements (max–min > 140 µV in windows of 640 ms) were removed from the analysis. Channels with more than 20% artifacts across the task were replaced with data interpolated from surrounding channels using spherical splines. Baseline correction was performed in reference to prestimulus activity, and individual averages were similarly referenced to the global field power reference.
Data analyses
Behavioral analysis
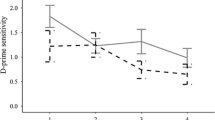
Trials with correct responses in both the control and VAS tasks were first selected. The absolute values of correct reaction times (RTs) longer than three standard deviations above the average were removed, and the remaining data were input into the following analyses. Independent t tests were submitted to examine the group differences separately in correct RTs and d’ values in the control task. The mean RTs and d’ values in the visual one-back task were evaluated with a 2 (group: RFD, NR) × 3 (eccentricity: inner = performance in position 3, middle = mean performance in position 2 and position 4, and outer = mean performance in position 1 and position 5) mixed measure analysis of variances (ANOVAs). Pilot analyses reported that participants’ performances exhibited bilateral symmetry in the VAS task, in which mean RTs and d’ values did not differ significantly between the first and fifth positions or between the second and fourth positions. Therefore, we composited the participants’ performances in the first and fifth positions and in the second and fourth positions and generated a variable of eccentricity to examine the position effect here in combination with the performance in the third position (Fig. 1d). Furthermore, a Pearson correlation analysis was conducted to examine the possible relationship between VAS task scores and reading skills.
ERP analysis
The EEG signals in the control and VAS tasks were separately computed for every participant at each of the relevant electrode sites by averaging the trials with correct responses. The datasets of all 36 participants were included in the following analyses, because the percentage of valid trials in the inner level of eccentricity was above 50% for each of the participants. String-locked ERPs were extracted to examine the P1 and N1 components. According to the topographic map (Fig. 2c), the P1 and N1 components were recorded in the posterior sites (i.e., P7, Pz, P8; O1, Oz, O2). Visual inspection of the EEG data across participants allowed us to identify a P1 component as the first visible positive peak within the 90–150-ms time window after string onset and a follow-up N1 component, which was identified as the first negative peak within the 150–210-ms time window after string onset. Moreover, referring to the relevant literature (Lallier et al. 2013) and the topographic map (Fig. 2c), the time window of the target-locked P3 component was 300–500 ms after the onset of the target item, which was measured at electrodes in the frontal–central sites (including FC1, FCz, FC2, C1, Cz, and C2). Considering the task setting, the 100-ms duration before the target onset corresponded to the presentation of the mask; therefore, this interval was not appropriate to be used as a baseline for the P3 component. In the current study, we utilized the baseline of string-locked P1/N1 components as the baseline of the target-locked P3 component; consequently, the time window of the P3 component was 600–800 ms after string onset (i.e., 200 ms string + 100 ms mask + 300–500 ms after the target onset) for both the control task and the visual attention span task. We separately conducted two-way repeated-measures ANOVAs with target eccentricity (three levels: inner, middle, and outer) as a within-subject factor and group (two levels: RFD and NR) as a between-subject factor on the peak amplitude of the P1 component and the mean amplitudes of the P3 component in the VAS task. Left lateralization may occur in the N1 component based on its relation with reading expertise (Brem et al. 2009; Lochy et al. 2016); thus, we added another between-subject factor, i.e., electrode site (three levels: left hemisphere, midline, and right hemisphere), and conducted a three-way ANOVA to analyze the peak amplitude of the N1 component to examine the possible influence of reading experience on VAS-related processing. Moreover, we conducted independent-sample t tests on the P1 and P3 amplitudes of the control task. Similar to the data analysis of the N1 component in the experimental task, the electrode site factor was added and a two-way ANOVA (3 electrode site × 2 group) was carried out on the N1 peak amplitude of the control task. The Greenhouse–Geisser correction was applied when the sphericity assumption was violated in the ANOVA. Bonferroni correction was adopted for multiple comparisons in post hoc analyses of ANOVAs. Moreover, Pearson correlation analyses were utilized to examine the relationship between the ERP measures and behavioral performances on the VAS task as well as on the reading tasks.

Comparisons of grand-averaged ERPs at relevant electrodes in different conditions. P1 and N1 components are string-locked (marked by “string onset”), and the corresponding electrodes include P7, Pz, P8, O1, Oz, and O2; P3 components are target-locked (marked by “target onset”), which were recorded at FC1, FCz, FC2, C1, Cz, and C2 electrodes. RFD individuals with poor reading fluency; NR age-matched normal readers. a The comparisons between individuals with poor reading fluency and normal readers within each level of eccentricity for different ERP components. Significant group differences were observed in string-locked N1 components at middle and outer positions and in target-locked P3 component at the outer position. b Significantly main effects of position in string-locked N1 component for the normal readers and in target-locked P3 component for the poor readers. c The significantly main effect of electrode site, with greater N1 amplitude in the left site than that in the midline and the right site. d The topographic maps for each component. *p < 0.05; **p < 0.01; ***p < 0.001
Results
Behavioral results
Control task
The results showed a significant difference between groups only in RTs (t34 = 2.69, p = 0.011) but not in d’ values (t34 = 0.20, p = 0.85). Adults with poor reading fluency identified an individual figure more slowly than NRs did (Table 1).
VAS task
Given that there was a significant group difference for the reaction times in the control task, reaction times for the control task were regarded as a covariate in the two-way ANOVA on reaction times for the VAS task. Relevant results showed that there were no significant main effects or interactions (ps > 0.1).
The results for the d’ values showed that the main effect of group was significant [F(1, 34) = 4.09, p = 0.04, η2 = 0.11]. Specifically, adults with poor reading fluency exhibited lower d’ values than normal readers (Table 1). The main effect of eccentricity was significant [F(2, 68) = 76.94, p < 0.001, η2 = 0.69]. Post hoc analyses showed that the d’ values in the inner level of eccentricity were higher than those in the other levels of eccentricity (ps < 0.001), while the d’ values did not significantly differ between the middle and outer levels (p > 0.1). There were no other significant comparisons or interaction effects (ps > 0.1). The results of the correlation analysis showed that single-character reading speeds were significantly correlated with the average d’ values (r = 0.45, p < 0.01) and d’ values in the noncenter positions (middle level: r = 0.42, p = 0.011; outer level: r = 0.56, p < 0.001); that is, higher reading speed corresponded to better performance in the VAS task.
ERP results
Control task
The grand-average ERPs recorded at relevant electrode sites in the control task for each group are shown in the Supplemental Materials. Group differences in the P1, N1, or P3 amplitudes were not significant (p > 0.1). The main effect of electrode site was significant for the N1 amplitude [F(2,68) = 23.35, p < 0.001, η2 = 0.41], with a more negative N1 amplitude in the left hemisphere than in the midline and right hemisphere (p < 0.05).
VAS task
Figure 2 displays the grand-average ERP waveforms between groups at different levels of eccentricity. We did not observe main effects (ps > 0.1) or group-by-eccentricity interactions [F(2,68) = 0.49, p = 0.62] on the string-locked P1 amplitude.
The N1 peak amplitude results showed that the main effect of group was significant [F(1,34) = 9.56, p = 0.004, η2 = 0.22], with a more negative amplitude for NRs than adults with poor reading fluency. The main effect of electrode site was significant [F(2,68) = 37.25, p < 0.001, η2 = 0.52], in which the peak amplitude in the midline was lower than that in both hemispheres (ps < 0.001) and the peak value in the left site was more negative than that in the right site (p = 0.04). The main effect of eccentricity was significant [F(2,68) = 4.34, p = 0.02, η2 = 0.11], with a more negative amplitude in the outer level of eccentricity than in the inner level of eccentricity. The eccentricity × group interaction was significant [F(2,68) = 3.78, p = 0.03, η2 = 0.10]. The simple effect analysis showed that in the inner level of eccentricity, significant differences did not occur between groups (p > 0.1); however, in the middle and outer levels, normal readers exhibited a more negative N1 amplitude than the poor readers (ps < 0.01). Meanwhile, the eccentricity effect was only significant for normal readers (p < 0.01) but not for RFD-risk individuals (p > 0.1), with a more negative N1 amplitude in the outer level of eccentricity than that in the inner level of eccentricity for the normal readers.
The mean amplitude results for the P3 component showed a significant group effect [F(1,34) = 6.40, p = 0.02, η2 = 0.16], in which the mean amplitude of age-matched controls was significantly greater than that of RFD adults. The main effect of eccentricity was also significant [F(2,68) = 5.32, p = 0.007, η2 = 0.14]. Post hoc analyses showed that the P3 amplitudes induced by targets appearing in the inner eccentricity were stronger than those in the outer and middle levels of eccentricity (ps < 0.05), with no significant differences between the other two conditions (p > 0.1). The group × eccentricity interaction was significant [F(2,68) = 3.27, p = 0.04, η2 = 0.09]. Simple effect analyses showed that a significant group effect was only observed in the outer level of eccentricity (NR > RFD, p < 0.001) but not in the other levels of eccentricity (ps > 0.1). Adults with RFD exhibited differences across eccentricities with lower P3 amplitude in the outer level than in the other levels (ps < 0.05), while the eccentricity effect was not significant in the normal readers (p > 0.1).
Further correlation analyses showed that the peak N1 amplitude in the outer condition was negatively correlated with the d’ values of the VAS task (r = − 0.47, p = 0.04) and reading speed (single-character: r = − 0.33, p = 0.053, marginally significant; sentence: r = − 0.42, p = 0.011), in which a more negative N1 component when the target was presented in the outer position corresponded with a better performance in the VAS and reading tasks. The mean amplitude of the P3 component in the outer eccentricity condition was positively correlated with the d’ values of the VAS task (r = 0.32, p = 0.056, marginally significant) and single-character reading speed (r = 0.34, p = 0.046), in which a greater P3 component in the outer condition corresponded to higher scores in the VAS and single-character-reading tests.
Discussion
After excluding the possibility of group differences in identifying single symbols (i.e., performance in the control task), the present study found that the performance in the VAS test was worse (i.e., lower task sensitivity) for Chinese adults with poor reading fluency than for the age-matched controls, thus reflecting that weakness in VAS capacity was still remarkable in these adult poor readers at a late developmental stage. Meanwhile, we explored the internal mechanism underlying this poor VAS skill by disentangling the temporal processing of VAS via an ERP technique. The current findings showed that abnormal electrophysiological activities were observed in the adults with poor reading fluency compared to the age-matched normal readers from the early stages of visual attentional processing with revealing an inability to sufficiently process the visual features of the string, to the later stage with reflecting problems in further encode the targets at different spatial positions.
At the behavioral level, a low score in the VAS task was reported in adults with poor reading fluency and the behavioral measure of VAS skill was significantly correlated with Chinese reading fluency in the present study. The adult participants here received higher education and thus had accumulated reading experience. Their low VAS capacity suggested that the enhanced reading experience probably could not compensate for their VAS problems. The current result was consistent with the prediction stated in the Introduction section; that is, the low VAS capacity in RFDs may remain remarkable in the context of languages with deep orthographies, even at the later stage of development, which could be attributed to the global reading strategy that processes large units of orthographic input in languages with deep orthographies (Antzaka et al. 2018). Chinese has a particularly deep orthography, and reading Chinese depends a great deal on global processing through the lexical route, especially in terms of a global visual analysis of Chinese characters’ forms and the direct mapping between the whole character and relevant semantics, even for adults (Huang et al. 2019; Liu et al. 2016). VAS could play an important role in this processing. Thus, VAS problems in Chinese adults with RFD could be clearly observed.
The electrophysiological results showed that adults with poor reading fluency exhibited weaker N1 amplitudes than normal readers, and further analyses revealed that this group difference mainly existed in the middle and outer levels of eccentricity but not in the inner level of eccentricity. The N1 component is mainly related to early visual processing and specifically linked to selective attention of relevant elements required to perform a task (Beaucousin et al. 2013). Amplitudes of the N1 component elicited by visual stimuli are typically enhanced when selective attention is directed to the target stimulus, which has been generally interpreted as attention-related facilitation of visual processing (Wiegand et al., 2014). The weaker amplitude of the N1 component for adults with poor reading fluency might reflect their inefficiency in directing selective attention to visually discriminate stimuli within the string, especially for stimuli presented in the noninner (middle and outer) levels of eccentricity within the attentional span. Furthermore, the N1 component has been suggested to change with the enhancement of reading experience and could be interpreted as the refinement of visual processes, especially for script materials (Bach et al. 2013; Bentin et al. 1999; Kim and Strakova 2012; Maurer et al. 2006; Yoncheva et al. 2015). Given the usage of script-like symbols in the current study, the N1 component might reflect the potential link with reading skills. Particularly, the significant left-lateralized pattern in the present N1 component, which was consistently found in previous studies exploring visual form processing of words and characters (Lin et al. 2011; Sacchi et al. 2016), supported the possibility that this N1 component is associated with reading. Accordingly, another possible explanation for the group difference in the N1 component could be provided. In detail, nonfluent readers may be unable to sufficiently process this visual string due to their lower reading level as opposed to the abilities of fluent readers, and the discrepancy in reading abilities between groups might cause a difference in the N1 component. However, the results of the current control task showed that significant group differences did not occur in the N1 amplitudes. If different levels of reading expertise could cause group differences in the N1 amplitudes in the visual one-back task, a similar pattern of group differences should also be observed in the control condition. The present finding of a nonsignificant group difference in the N1 amplitudes in the control task may exclude this possibility of the influence of different visual processing efficiencies related to reading expertise to some extent.
Additionally, significant group differences at the early stages of multielement processing were only observed in N1 amplitudes and not in P1 amplitudes. This result may be accounted for by the link between P1/N1 and local/global processing (Beaucousin et al. 2013). It has been suggested that the P1 component might be more involved in local processing and that N1 is modulated by the spread of attention for global processing (Beaucousin et al. 2013; Marini and Marzi 2016). In the VAS task, participants were required to simultaneously process multielement strings involved in global visual decoding of the elements in parallel; therefore, this global processing could be greatly reflected by the N1 component. Moreover, group differences on the N1 component but not the P1 component in this study could alternatively be explained by the greater involvement of N1 in the combination of bottom-up and top-down processing and the greater involvement of P1 on attentional bottom-up processing (Schiff et al. 2006). Accordingly, in the present study, poor readers might have difficulty in more complex attentional processing, such as linking script-like symbols in a string with similar representations stored in long-term memory, while their ability to encode the basic features of the items in one string seemed to be normal.
Moreover, larger P3 amplitudes were observed in normal readers than in adult poor readers. The study of Lallier et al. (2013) indicated that the target-locked P3 amplitude was viewed as an index of how well the target letter was encoded in the string, in which a higher P3 amplitude corresponded to better target letter encoding. Therefore, a better match was observed between the target letter and the probe strings for normal readers than for the adults in the RFD group. In addition, the P3 component was considered an index of the later cognitive processes related to allocating and maintaining attentional resources (Berndt et al. 2019; Corbetta et al. 2008). The weaker P3 amplitudes for the RFD group might indicate that the nonfluent readers loaded insufficient attention resources while detecting targets in the visual string. The combination of the findings based on N1 and P3 components revealed that the VAS deficit in nonfluent readers could be reflected both at the early stage of directing visual spatial attention to globally process the visual features of the multielement string and at the late processing stage of further encoding targets within the string.
In the present study, a position-based analysis of behavioral d’ values in the VAS task showed that both adults with poor reading fluency and age-matched normal readers presented a similar pattern of visual attention distribution, that is, a fixation advantage and a decreasing gradation with eccentricity enlargement. This distribution pattern was different from that in some other studies, in which a left-to-right bias can be seen (Awadh et al. 2016; Lallier et al. 2015; Valdois et al. 2014; van den Boer and de Jong 2018). This inconsistency might be associated with the differences in the characteristics of background languages. The leftward distribution of VAS resources has primarily been reported in studies in alphabetic language systems with left-to-right reading direction (Awadh et al. 2016; Banfi et al. 2018; Lallier et al. 2015; Valdois et al. 2014; van den Boer and de Jong 2018), in which readers utilized the sublexical decoding strategy of letter-by-letter spelling during reading (Frith 1985). This reading strategy would probably exert an influence on processing the string in VAS tasks, resulting in allocating more attention to the left part of the string (i.e., the starting point in reading habit). In contrast, readers in Chinese rely on the globally lexical route during reading (Huang et al. 2019) and usually decode a word as a whole. Similarly, these readers globally processed the string in VAS tasks and distributed more attention on the fixation point. In particular, this left bias has been found to be more remarkable in VAS tasks using letters as stimuli (Awadh et al. 2016; Banfi et al. 2018; Valdois et al. 2014; van den Boer and de Jong 2018) and global reports as oral responses (Valdois et al. 2014; van den Boer and de Jong 2018) but without task instructions for focusing on the screen center throughout the test (Banfi et al. 2018). These settings make VAS-related processing closer to the reading procedure, which would boost the influence of reading experience on visual simultaneous processing. In contrast, the present study used nonverbal symbols as stimuli and partial reports as nonverbal responses. Meanwhile, current participants were required to fixate on the screen center, which would make the VAS processing in the present study distinct from reading processes. Moreover, the differences in distribution patterns of VAS resources could also be related to the string spanning a larger visual angle in the current study (7.9°) than in other studies (e.g., approximately 5.5°; Bosse et al. 2007; Valdois et al. 2014; van den Boer and de Jong 2018). A smaller visual angle of the visual string could enable fovea to process the letters presented in the endpoints of the string (including the left endpoint), which might contribute to the leftward bias to some extent. Whereas a larger visual angle of the string in the present study might make the endpoints of the string locating in the parafoveal visual field, which would weaken the advantage of endpoint processing.
On a related note, the ERP results suggested that there were significant group differences in eccentricity (position) effects from a temporal perspective, which were reflected not only at the late processing stages of further encoding targets within strings (i.e., P3 component), as predicted in the Introduction section, but also at the early stage of selective attention processing (i.e., N1 component). In detail, NRs exhibited an eccentricity effect in N1 amplitudes but not in P3 amplitudes, with more negative N1 amplitudes in the outer level of eccentricity than in the inner level of eccentricity. However, an opposite pattern was observed in adults with poor reading fluency, and a significant eccentricity effect was observed in the P3 component instead of the N1 component, in which stronger amplitudes were observed in the outer level of eccentricity than in the other levels of eccentricity. This finding suggested a temporal dissociation of the eccentricity effect between the RFD and NR groups. In particular, the normal readers efficiently directed their selective attention to relevant locations within a string according to the experimental context, whereas the RFDs did not exhibit distinguished responses to items across eccentricities within a string until the late stage of further target encoding. N1 amplitudes were assumed to index object discrimination processes within the attentional focus (Vogel and Luck 2000). In particular, the N1 response has been shown to increase with increasing difficulty of the required visual discriminations (Wiegand et al. 2014). The target was difficult to identify when it was presented in the outer level of eccentricity within a string compared to the inner level of eccentricity in the present study, and thus, the N1 amplitudes in the outer level of eccentricity were greater than those in the inner level. This significant eccentricity effect in the normal readers suggested that they have attentional weight in allocating selective attention across items within a string at the early stage of visual processing; however, adults with poor reading fluency could not effectively direct their attention at different levels of eccentricity within a multielement string until the late processing stages (i.e., P3 component). Since the P3 component has been used to examine the efficiency of target encoding within a string (Lallier et al. 2013), the lower P3 amplitude in the outer level of eccentricity observed in the RFDs revealed problems in processing the items at the outer location of a multielement string, further suggesting their narrow span in visual spatial attention; in contrast, the degrees of target encoding were high and similar across each eccentricity condition for the normal readers, revealing that their visual attentional window may be larger than the length of the current string. Furthermore, the group-comparison findings revealed that the poor readers showed weaker N1 and P3 amplitudes in the noncenter positions (especially in the outer level) compared to the normal readers, and these weaker amplitudes were correlated with worse performances in the VAS and reading tests, as suggested by the present results of the relationship between ERP and behavioral measures. N1 is related both to attention and expertise in processing horizontally presented script-like symbols (Bach et al. 2013; Lin et al. 2011), and P3 is closely associated with whether the participant identifies a match between the target symbol and a previously seen symbol in the string (Lallier et al. 2013). Accordingly, an alternative explanation for the worse performance in the VAS task of these poor readers could be proposed. That is, because these dysfluent readers did not efficiently process the first presentation of the symbol within the string (especially for these outer position targets), they could not identify the “match” between the target and the symbol that appeared in the symbol string.
In summary, this study found a VAS deficit in the current cohort of Chinese adults with poor reading fluency, revealing that VAS problems occur in adult nonfluent readers in languages with particularly deep orthography, even at later developmental stages. The electrophysiological results, especially those based on the N1 and P3 components, revealed the possible underlying mechanism of the low VAS capacity in individuals with reading dysfluency from the temporal aspect; that is, their poor VAS skills could be reflected from the early stage of directing selective attention within the multielement string (i.e., the N1 component) to the late processing stage of allocating attention resources to encode the target out of surrounding elements (i.e., the P3 component). The present findings extend our understanding of the role of VAS in fluent reading from a temporal perspective and suggest the impact of language transparency on this relationship. Moreover, the current results may provide useful directions for future interventions on reading dysfluency, which could be designed to focus on the training of visual attention processes to further improve reading proficiency. It should be noted that the current study does not provide insights into the directionality of the relationship. Future longitudinal and intervention studies are required to better clarify the causality of the relationship. Additionally, the current study classified individuals as having poor reading fluency if their scores on at least one of the reading tests were − 1 SD below the average scores of the normal readers, which is consistent with previous research (Bertoni et al. 2019; Kuhl et al. 2020). Moreover, poor readers in the present study exhibited lower d’ values on the VAS task, revealing a low capacity in VAS-related processing. However, this group could not be confidently classified as having a visual attention span deficit. Therefore, future studies can apply more systematic and rigorous procedures to identify participants with RFD or with a VAS deficit to further ensure the mechanism underlying the relationship between VAS deficit and RFD. Furthermore, the current results of correlation results did not keep significant after Bonferroni correction. Thus, we will recruited a larger sample size of participants so as to further examine the relationship between VAS and reading in the following studies. Meanwhile, there are some limitations in the settings of VAS tasks, which might exert an influence on the final conclusion. For example, only a 100-ms duration was allowed before the onset of the target item, and the participants were required to respond as soon as the target stimulus appeared. Further studies could extend the interval between the onsets of the string and target stimuli and adopt a delayed response to ensure a flatter waveform before the target onset, which is more suitable as a baseline and to decrease the influence of motor responses on EEG recording.
Data availability
The datasets in this study are available on request to the corresponding author.
References
Antzaka A, Acha J, Carreiras M, Lallier M (2018) Does the visual attention span play a role in the morphological processing of orthographic stimuli? Q J Exp Psychol 72(7):1704–1716
Awadh F, Phenix T, Antzaka A, Lallier M, Carreiras M, Valdois S (2016) Cross-language modulation of visual attention span: an Arabic-French-Spanish comparison in skilled adult readers. Front Psychol 7:307
Bach S, Richardson U, Brandeis D, Martin E, Brem S (2013) Print-specific multimodal brain activation in kindergarten improves prediction of reading skills in second grade. Neuroimage 82:605–615
Banfi C, Kemeny F, Gang M, Schulte-Korne G, Mo K, Landerl K (2018) Visual attention span performance in german-speaking children with differential reading and spelling profiles: no evidence of group differences. PLoS One 13(6):e0198903
Beaucousin V, Simon G, Cassotti M, Pineau A, Houde O, Poirel N (2013) Global interference during early visual processing: ERP evidence from a rapid global/local selective task. Front Psychol 4:539
Bentin S, Mouchetant-Rostaing Y, Giard MH, Echallier JF, Pernier J (1999) ERP manifestations of processing printed words at different psycholinguistic levels: time course and scalp distribution. J Cogn Neurosci 11(3):235–260
Berndt M, Bäuml JG, Menegaux A et al (2019) Impaired structural connectivity between dorsal attention network and pulvinar mediates the impact of premature birth on adult visual–spatial abilities. Hum Brain Mapp 40(14):4058–4071
Bertoni S, Franceschini S, Ronconi L, Gori S, Facoetti A (2019) Is excessive visual crowding causally linked to developmental dyslexia? Neuropsychologia 130:107–117
Bosse M, Tainturier MJ, Valdois S (2007) Developmental dyslexia: the visual attention span deficit hypothesis. Cognition 104(2):198–230
Brem S, Halder P, Bucher K, Summers P, Martin E, Brandeis D (2009) Tuning of the visual word processing system: distinct developmental ERP and fMRI effects. Hum Brain Mapp 30(6):1833–1844
Corbetta M, Patel G, Shulman GL (2008) The reorienting system of the human brain: from environment to theory of mind. Neuron 58(3):306–324
Eimer M (2010) Attending to quadrants and ring-shaped regions: ERP effects of visual attention in different spatial selection tasks. Psychophysiology 36(4):491–503
Frith U (1985) Beneath the surface of developmental dyslexia. In: Patterson KG, Marshall JC, Coltheart M (eds) Surface dyslexia: studies of phonological reading. Cambridge University Press, Cambridge, pp 301–330
Germano GD, Reilhac C, Capellini SA, Valdois S (2014) The phonological and visual basis of developmental dyslexia in Brazilian Portuguese reading children. Front Psychol 5:1169
González GF, Zaric G, Tijms J, Bonte M, Blomert L, van der Molen MW (2015) A randomized controlled trial on the beneficial effects of training letter-speech sound integration on reading fluency in children with dyslexia. PLoS One 10(12):e0143914
Hawelka S, Wimmer H (2008) Visual target detection is not impaired in dyslexic readers. Vision Res 48(6):850–852
Huang C, Lorusso ML, Luo Z, Zhao J (2019) Developmental differences in the relationship between visual attention span and Chinese reading fluency. Front Psychol 10:2450
Johannes S, Munte TF, Heinze HJ, Mangun GR (1995) Luminance and spatial attention effects on early visual processing. Brain Res Cogn 2:189–205
Kang J, Lee S, Park E, Leem H (2016) Event-related potential patterns reflect reversed hemispheric activity during visual attention processing in children with dyslexia: a preliminary study. Clin Psychopharm Neu 14(1):33–42
Kim AE, Strakova J (2012) Concurrent effects of lexical status and letter-rotation during early stage visual word recognition: evidence from ERPs. Brain Res 1468:52–62
Kuhl U, Neef NE, Kraft I et al (2020) The emergence of dyslexia in the developing brain. Neuroimage 211:1–11
Lallier M, Carreiras M, Tainturier M, Savill N, Thierry G (2013) Orthographic transparency modulates the grain size of orthographic processing: behavioral and ERP evidence from bilingualism. Brain Res 1505:47–60
Lallier M, Valdois S, Lassus-Sangosse D, Prado C, Kandel S (2014) Impact of orthographic transparency on typical and atypical reading development: evidence in french-spanish bilingual children. Res Dev Disabil 35(5):1177–1190
Lallier M, Acha J, Carreiras M (2015) Cross-linguistic interactions influence reading development in bilinguals: a comparison between early balanced French-Basque and Spanish-Basque bilingual children. Developmental Sci 19:1–14
Lallier M, Acha J, Carreiras M (2016) Cross-linguistic interactions influence reading development in bilinguals: a comparison between early balanced French-Basque and Spanish-Basque bilingual children. Dev Sci 19(1):76–89
Langer N, Benjamin C, Minas J, Gaab N (2015) The neural correlates of reading fluency deficits in children. Cereb Cortex 25(6):1441–1453
Lin SE, Chen HC, Zhao J, Li S, He S, Weng XC (2011) Left-lateralized N170 response to unpronounceable pseudo but not false Chinese characters—the key role of orthography. Neuroscience 190:200–206
Liu D, Chen X, Wang Y (2016) The impact of visual-spatial attention on reading and spelling in Chinese children. Read Writ 29(7):1435–1447
Lobier M, Zoubrinetzky R, Valdois S (2012) The visual attention span deficit in dyslexia is visual and not verbal. Cortex 48(6):768–773
Lobier M, Dubois M, Valdois S (2013) The role of visual processing speed in reading speed development. PLoS One 8(4):e58097
Lochy A, Van Reybroeck M, Rossion B (2016) Left cortical specialization for visual letter strings predicts rudimentary knowledge of letter-sound association in preschoolers. PNAS 113(30):8544–8549
Luck SJ (1995) Multiple mechanisms of visual-spatial attention: recent evidence from human electrophysiology. Behav Brain Res 71(1):113–123
Marini F, Marzi CA (2016) Gestalt perceptual organization of visual stimuli captures attention automatically: electrophysiological evidence. Front Hum Neurosci 10:446
Maurer U, Brem S, Kranz F et al (2006) Coarse neural tuning for print peaks when children learn to read. Neuroimage 33(2):749–758
Pammer K, Lavis R, Hansen P, Cornelissen PL (2004) Symbol-string sensitivity and children's reading. Brain Lang 89(3):601–610
Pammer K, Lavis R, Cooper C, Hansen PC, Cornelissen PL (2005) Symbol-string sensitivity and adult performance in lexical decision. Brain Lang 94(3):278–296
Reilhac C, Peyrin C, Demonet J, Valdois S (2013) Role of the superior parietal lobules in letter-identity processing within strings: FMRI evidence from skilled and dyslexic readers. Neuropsychologia 51(4):601–612
Sacchi E, Laszlo S (2016) An event-related potential study of the relationship between N170 lateralization and phonological awareness in developing readers. Neuropsychologia 91:415–425
Schiff S, Mapelli D, Vallesi A, Orsato R, Gatta A, Umilta C, Amodio P (2006) Top-down and bottom-up processes in the extrastriate cortex of cirrhotic patients: an ERP study. Clin Neuropshysiol 117:1728–1736
Valdois S, Bosse M, Ans B, Carbonnel S, Zorman M, David D, Pellat J (2003) Phonological and visual processing deficits can dissociate in developmental dyslexia: evidence from two case studies. Read Writ 16(6):541–572
Valdois S, Peyrin C, Lassus-Sangosse D, Lallier M, Demonet JF, Kandel S (2014) Dyslexia in a French-Spanish bilingual girl: behavioural and neural modulations following a visual attention span intervention. Cortex 53(4):120–145
Valdois S, Lassus-Sangosse D, Lallier M, Moreaud O, Pisella L (2019) What bilateral damage of the superior parietal lobes tells us about visual attention disorders in developmental dyslexia. Neuropsychologia 130:78–91
van den Boer M, de Jong P (2018) Stability of visual attention span performance and its relation with reading over time. Sci Stud Read 22(5):434–441
Vogel EK, Luck SJ (2000) The visual N1 component as an index of a discrimination process. Psychophysiology 37:190–203
Wang C, Zhang Q (2021) Word frequency effect in written production: evidence from ERPs and neural oscillations. Psychophysiology 58:e13775
Wiegand I, Töllner T, Habekost T, Dyrholm M, Müller HJ, Finke K (2014) Distinct neural markers of TVA-based visual processing speed and short-term storage capacity parameters. Cereb Cortex 24:1967–1978
Yeari M, Isser M, Schiff R (2017) Do dyslexic individuals present a reduced visual attention span? Evidence from visual recognition tasks of non-verbal multi-character arrays. Ann Dyslexia 67(2):128–146
Yoncheva YN, Wise J, McCandliss B (2015) Hemispheric specialization for visual words is shaped by attention to sublexical units during initial learning. Brain Lang 145–146:23–33
Zhao J, Kwok R, Liu M, Liu H, Huang C (2017) Underlying skills of oral and silent reading fluency in Chinese: perspective of visual rapid processing. Front Psychol 7:2082
Zhao J, Liu M, Liu H, Huang C (2018) The visual attention span deficit in chinese children with reading fluency difficulty. Res Dev Disabil 73:76–86
Zhao J, Liu HL, Li JX, Sun HX, Liu ZH, Gao J, Liu Y, Huang C (2019) Improving sentence reading performance in Chinese children with developmental dyslexia by training based on visual attention span. Sci Rep 9(1):18964–19019
Ziegler JC, Goswami U (2005) Reading acquisition, developmental dyslexia, and skilled reading across languages: a psycholinguistic grain size theory. Psychol Bul 131(1):3–29
Zoubrinetzky R, Collet G, Serniclaes W, Nguyen-Morel M, Valdois S (2016) Relationships between categorical perception of phonemes, phoneme awareness, and visual attention span in developmental dyslexia. PLoS One 11(3):e0151015
Acknowledgements
We thank all the adults who participated in this study and the experimenters for their hard work during the data collection. This work was supported by the National Natural Science Foundation of China [Grant Numbers: 31871117].
Funding
This work was supported by National Natural Science Foundation of China [Grant Numbers: 31871117].
Author information
Authors and Affiliations
Contributions
JL conceptualization; investigation; methodology; data curation; Formal analysis; Software; validation; visualization; writing—original draft preparation. JZ conceptualization; funding acquisition; project administration; resources; visualization; supervision; validation; writing—reviewing and editing. JH supervision; validation; visualization. HL methodology; data curation; software.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
The research was approved by the Research Ethics Committee of School of Psychology, Capital Normal University. This study was carried out in accordance with The Code of Ethics of the World Medical Association (Declaration of Helsinki) for experiments involving humans.
Consent to participate
Informed consent was obtained from all individual participants included in the study.
Consent for publication
The participants signed informed consent regarding publishing their data.
Additional information
Communicated by Francesca Frassinetti.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Li, J., Zhao, J., Han, J. et al. Electrophysiological correlates of visual attention span in Chinese adults with poor reading fluency. Exp Brain Res 239, 1987–1999 (2021). https://doi.org/10.1007/s00221-021-06115-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-021-06115-7