
Abstract
Grasping a three-dimensional (3D) object results in the specification of motor output via absolute size information. In contrast, the impoverished visual cues (e.g., binocular and vergence) associated with grasping a two-dimensional (2D) object are reported to render aperture formation via an object’s perceptual and relative visual features. It is, however, important to recognize that 3D and 2D grasping differ not only in terms of their visual properties, but also because the latter does not entail the provision of haptic feedback. As such, the present work examined whether haptic feedback influences the nature of the information supporting 2D grasping. Participants grasped differently sized 3D objects (i.e., 3D task) and completed a ‘traditional’ 2D grasping task to line drawings without receiving haptic feedback (i.e., 2DH− task). As well, we included a separate condition using the same objects as the 2DH− task; however, the experimenter placed a 3D object (i.e., one corresponding to the size of the 2D object) between the thumb and forefinger of participants’ grasping limb once they completed their response (i.e., 2DH+ task). Thus, the 2DH+ task provided haptic feedback related to absolute object size. Notably, we computed just-noticeable-difference (JND) scores to determine whether the different tasks adhered to, or violated, the relative psychophysical principles of Weber’s law. JNDs for the 2DH− task adhered to Weber’s law, whereas 3D and 2DH+ tasks violated the law. Thus, results evince that 2DH− and 2DH+ tasks are specified via relative and absolute object size information, respectively. Accordingly, we propose that haptic feedback supports an absolute visuo-haptic calibration and contend that our results highlight the importance of multi-sensory cue integration in goal-directed grasping.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Reaching to grasp an object is a fundamental activity of daily living, requiring that the motor system be precisely calibrated to the extrinsic (e.g., height, weight) and intrinsic (e.g., density) properties of a to-be-grasped object. An extensive literature has shown that efficient and effective grasping is, in part, supported via absolute (i.e., metrically precise) visual information specified in an egocentric frame of reference via the dedicated visuomotor networks of the dorsal visual pathway (for review, see Goodale 2011). It is, however, important to recognize that mechano- and proprioceptive feedback (together referred to as haptic feedback) is associated with target-directed grasping. For example, touching an object with the thumb and forefinger (i.e., precision grasping) provides mechanoreceptor feedback related to the attainment of a stable grasp. Moreover, once an object has been securely grasped proprioceptive feedback related to thumb and forefinger position provides the motor system with absolute object size information (for review of haptic frames of reference, see Lederman and Klatzky 2009). The importance of haptic feedback in determining the nature of the information (i.e., absolute vs. relative) supporting grasping is demonstrated in Schenk’s (2012) work involving an individual (DF) with bilateral lesions to the lateral occipital cortex of her ventral visual pathway (James et al. 2003). Extensive work has shown that DF’s lesions impair her ability to use relative visual information (i.e., encoded in a scene-based/allocentric frame of reference) to support visual perceptions (e.g., discriminating the size and shape of an object). In contrast, DF’s grasping performance demonstrates metrical scaling to object size—a preserved ability attributed to the processing of absolute object information via her intact dorsal visual pathway (for extensive review of this issue, see Goodale and Milner 2013). Notably, Schenk’s work with DF entailed the use of a mirror box apparatus (see also Bingham et al. 2007), allowing the dissociation of the visual and physical location of a to-be-grasped object. Schenk reported that DF’s grasping performance in a block of trials wherein the object was physically unavailable to grasp was no better than her well-documented visuo-perceptual deficits—a result that was not unexpected given that Goodale et al. (1994) previously showed that DF exhibits impaired pantomime-grasping. More notably, Schenk reported that when a physical object was available at the movement goal—even when intermittently provided—DF’s grip aperture scaled to the absolute size of the object (but see Milner et al. 2012; Whitwell and Buckingham 2013). As such, Schenk proposed that DF’s impaired grasping performance occurs only in the absence of terminal haptic feedback and concluded that in grasping DF integrates haptic feedback to accommodate for her visuo-perceptual deficit. Although Schenk did not provide a mechanistic account for his findings, Whitwell et al. (2014) proposed that the importance of haptic feedback might arise over a series of trials by computing an error-related signal regarding the difference between an expected and an observed outcome that supports an absolute visuo-haptic calibration.
Recent work by our group (Davarpanah Jazi et al. 2015b; see also Davarpanah Jazi and Heath 2014; Davarpanah Jazi et al. 2015a) involving neurologically intact individuals has also shown that haptic feedback influences the information mediating goal-directed grasping. In particular, Davarpanah Jazi et al. (2015b) had participants (1) grasp a physical object (i.e., naturalistic grasping, NG), (2) perform a pantomime-grasp wherein the object was removed from the grasping environment (i.e., pantomime-grasp without haptic feedback, PH-), and (3) perform a pantomime-grasp wherein the experimenter returned the physical object to participants’ thumb and forefinger once they had completed their response (pantomime-grasp with haptic feedback, PH+) (for review of pantomime-grasping in clinical and non-clinical populations, see Cavina-Pratesi et al. 2011; Fukui and Inui 2013; Goodale et al. 1994; Holmes et al. 2013; Westwood et al. 2000). Thus, the PH− and PH+ tasks differed with regard to the availability of absolute haptic feedback. Notably, Davarpanah Jazi et al. computed just-noticeable-difference (JND) scores at the time of peak grip aperture to determine whether grasping tasks differentially adhered to, or violated, the psychophysical principles of Weber’s law. In particular, Weber’s law asserts that a JND reflects the smallest difference that one can reliably discriminate between an original (e.g., the target object) and a comparator (e.g., grip aperture) stimulus and is in constant proportion to the magnitude of the original stimulus. Moreover, the law states that the sensitivity of detecting a perceptual change in a physical continuum is relative as opposed to absolute. As such, adherence to Weber’s law is reflected via a systematic increase in JNDs with increasing stimulus intensity (i.e., increasing object size) and is taken to evince motor output specified via the relative properties of an object. In turn, violation of the law is indicated by JNDs that do not vary with object size and is taken to indicate grip aperture specification via absolute object size. Results showed that JNDs for the PH− task adhered to Weber’s law, whereas values for the NG and PH+ tasks violated the law. Accordingly, it was proposed that the presence (i.e., PH+) and the absence (i.e., PH−) of terminal haptic feedback in a pantomime-grasp task renders grip aperture specification via absolute and relative information, respectively. In particular, our group has proposed that an error-related signal arising from visual and haptic feedback supports an absolute calibration for motor output (Davarpanah Jazi et al. 2015b; for tactile–haptic calibration, see Davarpanah Jazi and Heath 2014; Davarpanah Jazi et al. 2015a).
The studies described above, as well as the majority of the extant literature, have examined the sensory and contextual features that influence grasping a three-dimensional (3D) object, that is, an object that affords a physical interaction (for description of grasping affordances, see Gibson 1986). It is, however, important to recognize that some grasping studies have employed two-dimensional (2D) objects as a proxy for 3D objects (e.g., Vishton et al. 1999). This represents a salient issue because 2D objects offer impoverished visual information due to the absence of stereoscopic and vergence cues related to shape information and are logically devoid of intrinsic properties. Moreover, human neuroimaging evidence has shown that 3D objects do not yield the same consistent and well-documented (Kanwisher et al. 1996) activation within the ventral and dorsal visual processing streams as 2D objects (Snow et al. 2011), and electrophysiological evidence from non-human primates has shown that neurons in the analogue region of the dorsal visual pathway are selectively active in response to binocularity disparity cues (e.g., Maunsell and Van Essen 1983; Janssen et al. 2000). On the basis of these findings, Snow et al. proposed that the real-world nature of 3D objects and their affordance for action is associated with neural mechanisms distinct from 2D objects. In addition, behavioral work by Holmes and Heath (2013) computed the JNDs associated with grasping differently sized (20, 30, 40, and 50 mm) 2D and 3D objects and showed a respective adherence and violation to Weber’s law. Holmes and Heath therefore proposed that grasping a 2D object is a visuo-perceptual task supported via relative visual information. Further, Freud and Ganel (2015) recently showed that a 2D—but not 3D—grasping task is prone to Garner interference (i.e., the processing of object properties along an irrelevant task dimension). Thus, convergent evidence suggests that grasping 2D and 3D objects entail the processing of relative and absolute visual information, respectively (for results in a virtual grasping environment, see also Viau et al. 2004).
Of course, we recognize that 2D and 3D objects differ not only in terms of their visual properties but also in terms of the availability of terminal haptic feedback. As such, the absence of physical grasp points on a 2D object may influence not only how an individual structures their response but also their ability to determine the object’s absolute size. In other words, differences in the neural mechanisms and information supporting 2D and 3D grasping may not selectively relate to an object’s visual properties; rather, differences may also reflect the absence of terminal haptic feedback in the former task. Given this limitation, the present investigation had participants grasp the same 2D and 3D objects as Holmes and Heath (2013). Importantly, however, 2D grasps were performed in separate blocks that manipulated the availability of terminal haptic feedback in a fashion similar to Davarpanah Jazi et al. (2015b). In particular, participants (1) ‘grasped’ 2D objects without haptic feedback (i.e., 2D without haptic feedback task, 2DH−) and (2) ‘grasped’ the same 2D objects as the 2DH− task; however, at the end of participants’ response the experimenter placed a 3D object (i.e., one corresponding to the size of the visually presented 2D object) between participants’ thumb and forefinger (i.e., 2D with haptic feedback task, 2DH+). Indeed, if terminal haptic feedback supports an error-related and absolute visuo-haptic calibration, then JNDs for the 2DH+ and 2DH− tasks should violate and adhere to Weber’s law, respectively.
Methods
Participants
Eighteen individuals (13 females and 5 males; age range = 18–30 years of age) from the University of Western Ontario community that self-declared being right-hand dominant, with normal or corrected-to-normal vision participated in this experiment. All participants signed consent forms approved by the Office of Research Ethics, University of Western Ontario, and the procedures associated with this project conformed to the Declaration of Helsinki.
Apparatus and procedures
Participants sat for the duration of the experiment in a height-adjustable chair placed in front of a tabletop (height = 780 mm, depth = 760 mm, width = 1060 mm) and reached to grasp—via precision grip (i.e., thumb and forefinger)—the long axis of a target object presented at their midline and 450 mm from the front edge of the table. Target objects were 3D and 2D and were painted/printed flat black. 3D target objects were weight-matched (i.e., 7 g) acrylic blocks 20, 30, 40, and 50 mm in width and 10 mm in height and depth and were secured to a 76.2 by 127.0 mm sheet of white paper (i.e., a cue card). 2D target objects were printed on the same surface as used to secure the 3D target objects (i.e., 2D objects were printed on a cue card) and were of the same width and depth as 3D target objects (i.e., 20, 30, 40, and 50 mm in width and all 10 mm in depth). Target objects were oriented perpendicular to participants’ midline. A pressure-sensitive switch located at midline and 50 mm from the front edge of the table served as the start location for each trial. Vision of the grasping environment was controlled via liquid-crystal shutter goggles (PLATO Translucent Technologies, Toronto, ON, Canada), and all computer-related experimental events were controlled via MATLAB (7.9.0, the MathWorks, Natick, MA, USA) and the Psychophysics Toolbox extensions (version 3.0) (Brainard 1997).
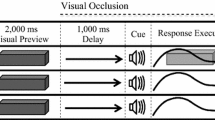
At the start of a trial, the goggles were set to the translucent state and participants rested the medial surface of their grasping (i.e., right) hand on the start location. During this time, the experimenter placed an appropriate target object on the tabletop. Once the target object was positioned, the experimenter cued a trial sequence wherein the goggles were set to their transparent state for a 2000-ms visual preview. At the end of the preview, a tone (2900 Hz for 10 ms) cued participants to initiate a grasping response (see details below). Participants were instructed to complete their response in a 600–800 ms movement time criterion, and received oral feedback following each trial (i.e., ‘too fast,’ ‘too slow,’ or ‘good’). Any trial falling outside of the movement time criterion was reentered into the trial matrix. At movement offset (see details below), the goggles reverted to their translucent state; thus, participants were provided online vision throughout their response. Notably, the aforementioned 2000-ms preview period and movement time criterion were used so that the current study directly matched the methods employed in a previous 2D grasping study (e.g., Holmes and Heath 2013). The movement time criterion was also used to avoid any possible confounds associated with distinct movement durations for grasping 3D and 2D objects. Moreover, and in line with Holmes and Heath, the basic instruction provided to participants was to ‘grasp the target object in the instructed movement time criterion.’
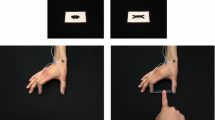
Grasping responses were completed in each of three tasks (see Fig. 1 for schematic). For the 3D task, participants grasped—but did not lift—and held the presented target object for 2000 ms before returning to the start location in preparation for a subsequent trial. For the 2D without haptic feedback task (i.e., 2DH−), participants were instructed to ‘grasp’ the target object and hold their final aperture for 2000 ms before returning to the start location. In this task, participants’ thumb and forefinger were in contact with the tabletop surface at the end of their response; however, the absence of a physical target precluded the integration of absolute haptic feedback related to object size. Moreover, and in line with our group’s previous work (Holmes and Heath 2013), we simply instructed participants to ‘grasp’ the 2D target object without providing further details—an instruction set designed to limit any potential bias related to task goals. For the 2D with haptic feedback task (i.e., 2DH+), the same procedures were used as per the 2DH− task; however, after movement offset (and when the goggles reverted to the translucent state), the experimenter placed a 3D target object (i.e., the same objects as used in the 3D task and one corresponding to the width of the presented 2D target object) between participants’ thumb and forefinger. More specifically, the experimenter placed the appropriate 3D target object on the tabletop and slid it along the surface until it contacted participants’ thumb and then positioned the opposite side to contact their forefinger. When the target object had been appropriately placed, participants adjusted their thumb and forefinger to achieve a stable grasp (i.e., a posture permitting stable lifting of the target object). The target object was then held—but not lifted—for 2000 ms before the participant released the target object and returned to the start location. Thus, the 2DH− and 2DH+ conditions provided equivalent grasping postures at movement offset; however, the provision of a physical object in the latter task provided absolute haptic feedback related to object size.

Schematic of visual, auditory and haptic events for 3D, 2DH− (i.e., 2D grasping without haptic feedback), and 2DH+ (i.e., 2D grasping with haptic feedback) tasks. For all trials, participants were provided a 2000-ms preview of either a 3D or 2D target object (i.e., 2DH− and 2DH+) and were subsequently cued via a tone to complete their grasping response. In the 3D task, the target was available to grasp at the movement goal location and therefore provided immediate terminal haptic feedback. In turn, for the 2DH+ task the experimenter placed an appropriately sized 3D object between participants’ thumb and forefinger once they achieved their movement goal location. As such, the 2DH+ condition provided experimenter-induced (and delayed) terminal haptic feedback. For the 2DH− task, the response was completed in the absence of any haptic feedback. Notably, for all tasks vision of the grasping environment was occluded at movement offset. The pictures below the schematic provide the participant’s egocentric view in each of the 3D (left panel), 2DH− (middle panel), and 2DH+ (right panel) tasks. For the 2DH+ task, the experimenter’s limb is visible, positioning the 3D object between the participant’s thumb and forefinger
3D, 2DH−, and 2DH+ tasks were performed in separate and randomly ordered blocks completed across two sessions separated by at least 24 h (i.e., one session involved the completion of two tasks, whereas the other session involved the completion of one task). Each block of trials required approximately 35 min to complete; hence, the separate sessions were used to limit mental fatigue. Within each block, each object size (i.e., 20, 30, 40, and 50 mm) was randomly presented 20 times (i.e., 80 trials per block and 240 total experimental trials). Further, we note that previous studies by our group (Davarpanah Jazi et al. 2015a, b) showed that the ordering of tasks performed with and without haptic feedback does not influence within-task performance. As well, we performed a blocked design as opposed to randomly interleaving 3D, 2DH−, and 2DH+ trials based on a previous 3D grasping study by Bingham et al. (2007). In particular, Bingham et al. showed that participants were able to produce an absolute visuo-haptic calibration even when haptic feedback was unpredictably varied from trial to trial. Thus, the blocked design used here provided the necessary framework to establish the independent role of haptic calibration for 2D grasping.
Data analysis
Infrared-emitting diodes (IREDs) were placed on the medial surface of the distal phalanx of the thumb, the lateral surface of the distal phalanx of the forefinger, and the styloid process of the radius. IRED position data were recorded at 400 Hz via an OPTOTRAK Certus (Northern Digital Inc., Waterloo, ON, Canada). A dual-pass Butterworth filter employing a low-pass cutoff frequency of 15 Hz was used to filter position data, and instantaneous velocities were computed from the position data via a five-point central finite difference algorithm. Movement onset was determined when participants released pressure from the start location switch, and movement offset was marked as the first frame wherein velocity fell below 50 mm/s for 20 consecutive frames (i.e., 50 ms).
Dependent variables and statistical analyses
Dependent variables included reaction time (RT: time from response cuing to movement onset), movement time (MT: time from movement onset to movement offset), peak grip aperture (PGA: maximal resultant distance between thumb and forefinger), and percent time to peak grip aperture (%tPGA).Footnote 1 As with previous work (Davarpanah Jazi and Heath 2014; Davarpanah Jazi et al. 2015a, b; Ganel et al. 2008; Heath et al. 2011, 2012; Holmes and Heath 2013; Holmes et al. 2011, 2013; Pettypiece et al. 2010), JNDs were computed as the within-participant standard deviations of PGAs. This technique is rooted in the Fechnerian principle that variability increases with stimulus uncertainty, and according to Ganel et al., represents a principle derived from the classic method of adjustment task wherein variance provides a measure of visuomotor uncertainty. Accordingly, the present investigation interprets a linear scaling of JNDs to increasing object size as adherence to Weber’s law. To further demonstrate our JND approach, the main panels of Fig. 2 provide trial-to-trial PGA values for an exemplar participant as a function of each task and object size used here. The figure shows that trial-to-trial PGAs for the 3D and 2DH+ tasks did not systematically vary with object size, whereas for the 2DH− task, trial-to-trial differences in PGAs increased with increasing object size. The smaller offset panels of Fig. 2 show the standard deviations (i.e., the JND values) associated with the PGA values presented in the main panels and demonstrate that JNDs for the 2DH− task, but not for the 3D or 2DH+ tasks, increased linearly with increasing object size.

Main panels present trial-to-trial peak grip aperture (PGA, mm) for an exemplar participant in the 3D (top panel), 2D without haptic feedback (i.e., 2DH− and see middle panel), and 2D with haptic feedback (i.e., 2DH+ and see bottom panel) tasks as a function of object size (20, 30, 40, and 50 mm). The smaller offset panels for each figure represent the participant's mean just-noticeable-difference (JND) scores (i.e., the standard deviations of PGA for each object size). The dashed line in the smaller panels represents the linear regression of JNDs to object size, and the top of each panel presents the associated linear regression equation and proportion of explained variance. The figure is designed to provide a demonstration of our computation and interpretation of JNDs. In particular, a linear scaling of JNDs to object size (see middle panel) is taken as adherence to Weber’s law, whereas JNDs that do not systematically vary with object size is taken as a violation of the law (see top and bottom panels)
Dependent variables were examined via 3 (task: 3D, 2DH−, 2DH+) by 4 (object size: 20, 30, 40, and 50 mm) repeated-measures ANOVA. Significant main effects and interactions (p < 0.05) were decomposed via paired-sample tests and/or power polynomials (i.e., trend analyses; see Pedhazur 1997).
Results
The grand means for RT and MT were 289 ms (SD = 46) and 696 ms (SD = 25), respectively, and these variables did not yield reliable main effects or interactions, Fs < 2.23, ps ≥ 0.10, all η 2 p < 0.16. Results for %tPGA indicated main effects for task, F(2,34) = 14.35, p < 0.001, η 2 p = 0.46, and object size, F(3,51) = 11.83, p < 0.001, η 2 p = 0.41: %tPGA occurred earlier for the 3D (74 %, SD = 6) than 2DH− (84 % ms, SD = 10) or 2DH+ (82 % ms, SD = 12) tasks [ts(17) = 5.09 and 3.42, ps < 0.01], which did not differ (t(17) = 1.61, p = 0.26), and %tPGA increased linearly with increasing object size for all tasks (only significant linear effect: F(1,17) = 14.81, p < 0.01).
In terms of PGA and JND, both variables yielded main effects for object size, Fs(3,51) = 1847.97 and 11.52, ps < 0.001, η 2 p = 0.99 and 0.40, and interactions involving task by object size, Fs(6,102) = 6.28 and 2.22, ps < 0.001 and 0.05, η 2 p = 0.27 and 0.12. In addition, PGA produced a main effect for task, F(2,34) = 97.91, p < 0.001, η 2 p = 0.85. The right ordinate of each panel in Fig. 3 presents mean PGA values and shows the expected increase in PGA with increasing object size for all tasks [significant linear effect: F(1,17) = 2632.93, p < 0.001] (see also top panel of Fig. 4). In addition, the figure demonstrates that PGAs for the 3D task were larger than for the 2DH+ task at each matched object size [ts(17) > 10.39, all ps < 0.001]. In turn, PGAs for the 2DH+ and 2DH− tasks did not reliably differ for the 20 mm object size [t(17) = 1.69, p = 0.10]; however, PGAs for the 2DH+ task were larger than for the 2DH− task at matched object sizes of 30, 40, and 50 mm [ts(17) = 2.75, 2.67, and 3.12, all ps < 0.02]. Notably, the left ordinate of each panel in Fig. 3 shows mean JNDs and demonstrates that values for the 3D and 2DH+ tasks did not reliably vary with object size [nonsignificant linear effects: Fs(1,17) = 2.33 and 1.27, ps = 0.14 and 0.27], whereas values for the 2DH− task increased with increasing object size [significant linear effect: F(1,17) = 27.10, p < 0.001]. In addition, the bottom panel of Fig. 4 presents mean participant-specific slopes and associated 95 % confidence intervals relating JNDs to object size for each task. The figure provides a graphical demonstration that JNDs for the 2DH− task increased systematically with increasing object size, whereas values for the 3D and 2DH+ tasks did not reliably vary with object size.

Mean just-noticeable-difference scores (JND in mm: see left ordinate) and peak grip aperture (PGA in mm: see right ordinate) as a function of object size (20, 30, 40, and 50 mm) for the 3D (top panel), 2D without haptic feedback (i.e., 2DH− and see middle panel), and 2D with haptic feedback (i.e., 2DH+ and see bottom panel) tasks. To highlight the difference between JND and PGA values within each panel, the former are presented as dark filled circles, whereas the latter are depicted as open light-gray squares. The dark and light hatched lines in each figure represent the respective regression lines for JND and PGA values to object size, and their associated regression equations and proportion of explained variance are presented at the top of each panel. Errors bars represent 95 % within-participant confidence intervals as a function of the mean-squared error term for object size computed separately for each task (Loftus and Masson 1994). Note: error bars for PGA are less than the size of the symbol associated with each mean value

Mean participant-specific slopes relating peak grip aperture (PGA: top panel) and just-noticeable-difference (JND: bottom panel) values to object size for the 3D, 2D without haptic feedback (i.e., 2DH−), and 2D with haptic feedback (i.e., 2DH+) tasks. Error bars represent the 95 % between-participants confidence intervals (Cumming 2013). The absence of overlap between error bars and zero represents a reliable linear effect that can be interpreted inclusive to a test of the null hypothesis
Last, participant-specific slopes relating PGA and JND to object size were examined via one-way repeated-measures ANOVA and results yielded reliable main effects, Fs(2,34) = 10.95 and 7.17, ps < 0.01. In particular, the mean PGA slope for the 2DH− task (0.81, SD = 0.07) was shallower than either the 3D (0.90, SD = 0.08) or 2DH+ (0.87, SD = 0.08) tasks [ts(17) = 4.35 and 3.82, ps < 0.01], and the latter two tasks did not reliably differ [t(17) = 1.48, p = 0.16]. In turn, the mean JND slope for the 2DH− task (0.04, SD = 0.02) was steeper than the 3D (0.007, SD = 0.02) and 2DH+ (0.008, SD = 0.02) tasks [ts(17) = 3.90 and 2.87, ps < 0.01], and the latter two tasks did not reliably differ [t(17) < 1]. In other words, mean PGA slopes for the 3D and 2DH+ tasks scaled to object size more than for the 2DH− task, and results for mean JND slopes support the assertion raised in the previous paragraph that the 2DH− task adhered to Weber’s law, whereas the 3D and 2DH+ tasks violated the law.
Discussion
Object dimension and terminal haptic feedback influence the timing and magnitude of peak grip aperture
The timing and magnitude of PGA increased linearly with increasing object size for 2D (2DH− and 2DH+) and 3D tasks. These expected results indicate that the visuomotor system contains the resolution necessary to differentiate between the differently sized objects used here (for resolution of visuomotor system, see Ganel et al. 2012). Moreover, such results indicate that 2D and 3D grasping are associated with similar kinematic features (e.g., Freud and Ganel 2015; Holmes and Heath 2013; Westwood et al. 2002). Notably, however, the 3D task produced larger and earlier occurring PGAs than either 2D task—a result supporting the view that grasping a physical object requires orthogonal thumb and forefinger approach vectors (i.e., double-pointing hypothesis; Smeets and Brenner 1999). Such a control strategy prevents an early collision with the object (for timing of 3D grasping, see Jeannerod 1984) and decreases spatial variability at the time of contact to ensure that the forces applied by the thumb and forefinger are opposite to one another (i.e., prevents the object from slipping). In turn, the smaller and later occurring PGAs for the 2D tasks indicate that orthogonal approach vectors are not required because: (1) the absence of a physical target precludes the risk of an object collision, and (2) the participant, and not the physical parameters of the object, determines task-based precision requirements for a successful 2D grasp. In other words, results for PGA indicate that the strategy and underlying control of grasping 2D and 3D objects are distinct.
An interesting difference between 2D tasks completed with (2DH+) and without (2DH−) terminal haptic feedback was that the former produced larger PGAs at matched object sizes (see Fig. 3).Footnote 2 These results are consistent with previous work by our group showing that haptic feedback provided after visually based (Davarpanah Jazi et al. 2015b) and tactile-based (Davarpanah Jazi et al. 2015a) pantomime-grasping renders larger PGAs than counterparts performed without haptic feedback. Accordingly, and as will be outlined in more detail in the following section, we propose that haptic feedback influences the manner a grasping response is structured and results in an improved—albeit incomplete—calibration of aperture kinematics (see also Bingham et al. 2007).
JNDs: haptic feedback and an absolute visuo-haptic calibration
JNDs for the 3D task did not reliably vary with object size, whereas values for the 2DH− task systematically increased with increasing object size. In other words, the 3D and 2DH− tasks violated and adhered to Weber’s law, respectively. These results match an earlier study by our group (Holmes and Heath 2013) and Freud and Ganel (2015) and support the contention forwarded by both groups that absolute visual information mediates the grasping of a physical object (i.e., 3D task), whereas a 2D object renders aperture shaping via holistic and relative visual information. In other words, the dissociable adherence of 3D and 2DH− tasks to Weber’s law has been taken to selectively reflect the use of dissociable visual codes (i.e., absolute versus relative visual information). Notably, however, JNDs for the 2DH+ tasks did not systematically vary with object size and elicited values on a par with their 3D counterparts. This finding indicates that the difference between the 2DH− and 3D tasks reported here and previously (i.e., Freud and Ganel 2015; Holmes and Heath 2013; Westwood et al. 2002) is not entirely visually based; rather, results indicate that terminal haptic feedback plays an important role in determining the nature of the integrative (i.e., visual and haptic) sensory cues supporting grasping control (Davarpanah Jazi et al. 2015b). Furthermore, we note that the difference between 2DH− and 2DH+ tasks cannot be attributed to a speed–accuracy trade-off in movement planning or control (Fitts 1954); after all, reaction time and movement time did not vary across the different experimental conditions used here. Moreover, results cannot be attributed to a range effect in aperture size (Lemay and Proteau 2001). Indeed, in spite of the fact that the 3D and 2DH+ tasks produced larger PGAs than the 2DH− task (see exception for 20-mm object in Footnote 2) only the latter task showed a systematic increase in JNDs with object size. Thus, the present findings in combination with our group’s previous pantomime-grasping study (Davarpanah Jazi et al. 2015b) demonstrate that the provision of terminal haptic feedback renders motor output that violates Weber’s law.
In accounting for the null scaling of JNDs in the 2DH+ task, it is important to recognize that the different object sizes used here were randomly varied from trial to trial. Thus, it was not possible for participants to rely on haptic feedback from trial N-1 to plan a subsequent response. Instead, the random presentation mandated a visuo-haptic calibration. Thus, and as proposed by Whitwell et al. (2014), the absolute scaling of 2DH+ trials may reflect that haptic information derived from finger and thumb orientation (i.e., when holding a physical object) serves to generate an error-related signal that is used in a predictive fashion to support an absolute visuo-haptic calibration. In this formulation, a difference between visual and haptic feedback arising from trial-to-trial performance serves as an information source to refine motor output. Alternatively, and although not mutually exclusive to the account described above, haptic feedback in a 2DH+ task may serve as a predominant (and absolute) information source in minimizing motor variance and optimizing motor output (i.e., maximum likelihood integrator model; Ernst and Banks 2002). Thus, haptic feedback may be used to refine/calibrate a response the next time the performer encounters the same object, and support the egocentric specification of object size (Lederman and Klatzky 2009). In other words, the 2DH+ task elicits a violation of Weber’s law on a par with 3D grasping because object size for each task type is specified via an egocentric—and absolute—frame of reference. In turn, because the 2DH− task does not provide integrative visuo-haptic cues, the impoverished visual information (e.g., the absence of stereoscopic vergence and disparity cues) associated with the task may render motor output selectively mediated by the object’s relative visual properties. Regardless of the explanation, we believe that the present results add importantly to the literature insomuch as they demonstrate that terminal haptic feedback plays an important role in accounting for differences in the information supporting 2D and 3D grasping tasks. As well, our results serve the general grasping literature in demonstrating that multi-sensory cues play an important role in determining the strategy and nature of information supporting aperture shaping.
Three final issues require addressing. The first relates to a recent study by Christiansen et al. (2014) reporting that JNDs for 2DH− and 3D tasks do not vary with object size (Note: Christiansen et al. did not employ a 2DH+ task). Accordingly, Christiansen et al. proposed that a common visual code (i.e., absolute) supports 2DH– and 3D grasping—a result countering the current and previous (Freud and Ganel 2015; Holmes and Heath 2013) work. In reconciling this discrepancy, Christiansen et al. presented 2D objects in the picture plane, whereas the present and previous work presented objects in the depth plane (Freud and Ganel 2015; Holmes and Heath 2013). Given this methodological difference, it could be argued that a 2D object in the picture plane permits the extraction of depth and surface information (as well as other stereoscopic cues) that are unavailable when the same object is presented in the depth plane (Turnbull et al. 2004). Thus, future work should directly examine whether the plane (depth vs. picture) in which a 2D object is presented influences the visual information supporting motor output. That being said, it is unclear from Christiansen et al.’s work at what point in the grasping response JNDs were computed. This is a salient issue because if JNDs were computed at the time of final grip aperture (i.e., 100 % of movement time) rather than at PGA, then results would not represent a predictive determination of object size in line with that quantified in the extant literature (see Jeannerod 1984). The second issue to address relates to Kwok and Braddick’s (2003) work showing that PGAs for grasps to 2D and 3D objects embedded in the Titchener circles illusion were refractory to the scene-based (i.e., relative) properties of the stimulus. Indeed, a prediction drawn from the present results would be that the relative features of the Titchener circles would result in 2D grasps that were ‘tricked’ in a direction consistent with the illusion’s perceptual effects. Notably, however, Kwok and Braddick did not report the timing of PGA (see Fig. 6 of their work), and it is therefore unclear from their work whether a late PGA onset for their 2D task precluded their ability to identify a reliable illusion effect. Of course, the present results and previous work (Freud and Ganel 2015; Holmes and Heath 2013; Westwood et al. 2002) suggest that the late onset of PGA in a 2D grasping task may exclude the variable from serving as a reliable metric for quantifying the influence of scene-based visual cues. The third issue to address relates to the null difference in the magnitude of JNDs between the 3D and 2DH+ tasks. Indeed, it could be predicted that the overall magnitude of JNDs for the 2DH+ task would be larger than the 3D task given that the former is less ‘practiced.’ In spite of such a prediction, the magnitude of JNDs across 3D and 2DH+ tasks did not differ. As such, we conclude that terminal haptic feedback not only supports an absolute visuo-haptic calibration, but also supports the efficiency and effectiveness of motor output.
Conclusions
The present study provides the first examination of the influence of terminal haptic feedback in a 2D grasping task. Results showed that 2D grasps performed with (i.e., 2DH+) and without (i.e., 2DH−) terminal haptic feedback violated and adhered to Weber’s law, respectively. Given these findings, we propose that terminal haptic feedback supports an absolute visuo-haptic calibration. What is more, the present findings not only serve to better understand how the dimensional properties of an object influence grasping, but also extend to the general grasping literature in highlighting the importance of multi-sensory feedback integration in goal-directed grasping.
Notes
We report %tPGA so that our findings can be conceptualized within the well-known normalized time frame for PGA (i.e., 76 % of MT) (see Jeannerod 1984). That said, the results and interpretation for absolute time to PGA mirrored those of %tPGA. In particular, results yielded main effects for task, F(2,34) = 15.94, p < 0.001, η 2p = 0.48, and object size, F(3,51) = 8.18, p < 0.001, η 2p = 0.32: time to PGA occurred earlier for the 3D (515 ms, SD = 45) compared to the 2DH− (585 ms, SD = 69) and 2DH+ (577 ms, SD = 87) tasks [ts(17) = 5.69 and 3.69, ps < 0.001] which did not differ (t(17) < 1), and values increased linearly with increasing object size [only linear effect significant: F(1,17) = 11.08, p < 0.01].
Although post hoc analyses showed that PGAs for the 20-mm object did not reliably differ between the 2DH− and 2DH+ tasks, Fig. 3 shows that the mean value for the latter task was larger. Thus, the direction of the difference between the 2DH− and 2DH+ tasks for the 20-mm object is consistent with the statistically reliable between-tasks differences reported for the larger object sizes used here (i.e., 30, 40, and 50 mm).
References
Bingham G, Coats R, Mon-Williams M (2007) Natural prehension in trials without haptic feedback but only when calibration is allowed. Neuropsychologia 45:288–294
Brainard DH (1997) The psychophysics toolbox. Spat Vis 10:433–436
Cavina-Pratesi C, Kuhn G, Ietswaart M, Milner AD (2011) The magic grasp: motor expertise in deception. PLoS ONE 6:e16568
Christiansen JH, Christensen J, Grünbaum T, Kyllingsbæk S (2014) A common representation of spatial features drives action and perception: grasping and judging object features within trials. PLoS ONE 9:e94744
Cumming G (2013) Understanding the new statistics: effect sizes, confidence intervals, and meta-analysis. Routledge, New York
Davarpanah Jazi S, Heath M (2014) Weber’s law in tactile grasping and manual estimation: feedback-dependent evidence for functionally distinct processing streams. Brain Cogn 86C:32–41
Davarpanah Jazi S, Hosang S, Heath M (2015a) Memory delay and haptic feedback influence the dissociation of tactile cues for perception and action. Neuropsychologia 71:91–100
Davarpanah Jazi S, Yau M, Westwood DA, Heath M (2015b) Pantomime-grasping: the ‘return’ of haptic feedback supports the absolute specification of object size. Exp Brain Res 233:2029–2040
Ernst MO, Banks MS (2002) Humans integrate visual and haptic information in a statistically optimal fashion. Nature 415:429–433
Fitts PM (1954) The information capacity of the human motor system in controlling the amplitude of movement. J Exp Psychol Gen 47:381–391
Freud E, Ganel T (2015) Visual control of action directed toward two-dimensional objects relies on holistic processing of object shape. Psychon Bull Rev 22:1377–1382
Fukui T, Inui T (2013) How vision affects kinematic properties of pantomimed prehension movements. Front Psychol 4:44
Ganel T, Chajut E, Algom D (2008) Visual coding for action violates fundamental psychophysical principles. Curr Biol 18:R599–R601
Ganel T, Freud E, Chajut E, Algom D (2012) Accurate visuomotor control below the perceptual threshold of size discrimination. PLoS ONE 7:e36253
Gibson JJ (1986) The ecological approach to visual perception. Routledge, Boston
Goodale MA (2011) Transforming vision into action. Vision Res 51:1567–1587
Goodale M, Milner D (2013) Sight unseen: an exploration of conscious and unconscious vision. Oxford University Press, Oxford
Goodale MA, Jakobson LS, Keillor JM (1994) Differences in the visual control of pantomimed and natural grasping movements. Neuropsychologia 32:1159–1178
Heath M, Mulla A, Holmes SA, Smuskowitz LR (2011) The visual coding of grip aperture shows an early but not late adherence to Weber’s law. Neurosci Lett 490:200–204
Heath M, Holmes SA, Mulla A, Binsted G (2012) Grasping time does not influence the early adherence of aperture shaping to Weber’s law. Front Hum Neurosci 6:332
Holmes SA, Heath M (2013) Goal-directed grasping: the dimensional properties of an object influence the nature of the visual information mediating aperture shaping. Brain Cogn 82:18–24
Holmes SA, Mulla A, Binsted G, Heath M (2011) Visually and memory-guided grasping: aperture shaping exhibits a time-dependent scaling to Weber’s law. Vision Res 51:1941–1948
Holmes SA, Lohmus J, McKinnon S, Mulla A, Heath M (2013) Distinct visual cues mediate aperture shaping for grasping and pantomime-grasping tasks. J Mot Behav 45:431–439
James TW, Culham J, Humphrey GK, Milner AD, Goodale MA (2003) Ventral occipital lesions impair object recognition but not object-directed grasping: an fMRI study. Brain 126:2463–2475
Janssen P, Vogels R, Orban GA (2000) Selectivity for 3D shape that reveals distinct areas within macaque inferior temporal cortex. Science 288:2054–2056
Jeannerod M (1984) The timing of natural prehension movements. J Mot Behav 16:235–254
Kanwisher N, Chun MM, McDermott J, Ledden PJ (1996) Functional imaging of human visual recognition. Brain Res Cogn Brain Res 5:55–67
Kwok RM, Braddick OJ (2003) When does the Titchener Circles illusion exert an effect on grasping? Two- and three-dimensional targets. Neuropsychologia 41:932–940
Lederman SJ, Klatzky RL (2009) Haptic perception: a tutorial. Atten Percept Psychophys 71:1439–1459
Lemay M, Proteau L (2001) A distance effect in a manual aiming task to remembered targets: a test of three hypotheses. Exp Brain Res 140:357–368
Loftus GR, Masson ME (1994) Using confidence intervals in within-subject designs. Psychon Bull Rev 1:476–490
Maunsell JH, Van Essen DC (1983) Functional properties of neurons in middle temporal visual area of the macaque monkey. II. Binocular interactions and sensitivity to binocular disparity. J Neurophysiol 49:1148–1167
Milner AD, Ganel T, Goodale MA (2012) Does grasping in patient D.F. depend on vision? Trends Cogn Sci 16:256–257 (discussion 258–259)
Pedhazur EJ (1997) Multiple regression in behavioral research: explanation and prediction. Harcourt Brace College Publishers, Orlando
Pettypiece CE, Goodale MA, Culham JC (2010) Integration of haptic and visual size cues in perception and action revealed through cross-modal conflict. Exp Brain Res 201:863–873
Schenk T (2012) No dissociation between perception and action in patient DF when haptic feedback is withdrawn. J Neurosci 32:2013–2017
Smeets JB, Brenner E (1999) A new view on grasping. Mot Control 3:237–271
Snow JC, Pettypiece CE, McAdam TD, McLean AD, Stroman PW, Goodale MA, Culham JC (2011) Bringing the real world into the fMRI scanner: repetition effects for pictures versus real objects. Sci Rep 1:130
Turnbull OH, Driver J, McCarthy RA (2004) 2D but not 3D: pictorial-depth deficits in a case of visual agnosia. Cortex 40:723–738
Viau A, Feldman AG, McFadyen BJ, Levin MF (2004) Reaching in reality and virtual reality: a comparison of movement kinematics in healthy subjects and in adults with hemiparesis. J Neuroeng Rehabil 1:11
Vishton PM, Rea JG, Cutting JE, Nuñez LN (1999) Comparing effects of the horizontal-vertical illusion on grip scaling and judgment: relative versus absolute, not perception versus action. J Exp Psychol Hum Percept Perform 25:1659–1672
Westwood DA, Chapman CD, Roy EA (2000) Pantomimed actions may be controlled by the ventral visual stream. Exp Brain Res 130:545–548
Westwood DA, Danckert J, Servos P, Goodale MA (2002) Grasping two-dimensional images and three-dimensional objects in visual-form agnosia. Exp Brain Res 144:262–267
Whitwell RL, Buckingham G (2013) Reframing the action and perception dissociation in DF: haptics matters, but how? J Neurophysiol 109:621–624
Whitwell RL, Milner AD, Cavina-Pratesi C, Byrne CM, Goodale MA (2014) DF’s visual brain in action: the role of tactile cues. Neuropsychologia 55:41–50
Acknowledgments
This work was supported by a Discovery Grant from the Natural Sciences and Engineering Research Council of Canada and Faculty Scholar and Major Academic Development Fund Awards from the University of Western Ontario. SH was supported by an Undergraduate Student Research Award from the Natural Sciences and Engineering Research Council of Canada.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Hosang, S., Chan, J., Davarpanah Jazi, S. et al. Grasping a 2D object: terminal haptic feedback supports an absolute visuo-haptic calibration. Exp Brain Res 234, 945–954 (2016). https://doi.org/10.1007/s00221-015-4521-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-015-4521-4