
Abstract
Fourteen healthy adults were tested to assess the potential influence on stance maintenance of two parts of the visual feedback technique (display scale and time delay). The task consisted in their keeping a spot on the screen representing their center of pressure, CoP (i.e. successive points of application of the ground reaction forces detected by the force platform on which they were standing) to a minimum size. The analysis focused on elementary motions computed from the complex CoP trajectories, that is the horizontal motion of the center of gravity (CoGh) and the difference between the CoP and the vertical projection of the center of gravity (CoP−CoGv). The former is recognized as the main variable in postural control, and several interesting features can be extracted from the latter. The results indicate that setting a delay and increasing the display scale induce substantial reductions in CoP−CoGv and CoGh displacements, respectively. Interestingly, when the two effects are combined, these single effects cohabit quite happily. Fractional Brownian motion modeling of these trajectories revealed clearly that, in each case, these effects originate principally from poor or improved control, respectively. This feature confirms that these elementary motions are involved differently in the postural system and that study of the complex CoP might not be of great interest. By generating opposing but complementary trends, the visual feedback technique should thus be perceived as a promising tool for inducing particular postural behavior in healthy and disabled individuals.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Standing perfectly still in bipedal upright stance is not humanly possible. The main reason is the inability of whole body motion, expressed through that of the center of gravity (CoG), to accurately follow (because of inertia) displacements imposed by the points of application of the resultant reaction forces, i.e. the center of pressure (CoP). In addition, the farther the CoP is from the CoG, the larger the horizontal acceleration communicated to the latter (Brenière et al. 1987; Caron et al. 1997). From that, the neurophysiological basis which enables the body to be constrained in a low range of motion consists in detecting body motion as quickly as possible, through various sensory cues, and initiating precise motion aimed at limiting body acceleration. When problems arise in this organization or, more simply, when there is a desire to improve one’s capacity to stand still, one way to enhance postural performance is to provide the subject with additional perceptual information. Although our somesthetic (through plantar pressure and joint and limb motion), visual, and vestibular systems are able, through integrative processes, to communicate how our whole body moves to the central nervous system, the information obtained through a force platform device is much more sensitive than plantar pressures. From a study based on a two-point discrimination threshold measured on the soles (Weinstein 1968), the average 20 mm thresholds are far superior to the precision of a standard force platform. This principle, known as visual feedback (VFB), consists in providing the displacements of the CoP to the subject on the force platform via a visual display (Rougier 1999a, 2003a). Giving the output of the force platform in both medio-lateral (ML) and AP directions has indeed been common practice in all previous studies conducted on this topic.
Recent studies have highlighted two aspects of VFB which have opposite effects on the elementary motions which, reconstituted, correspond to the complex CoP displacements—the horizontal motion of the center of gravity (CoGh) and the distance in the plane of support between the CoP and the vertical projection of the CoG (CoP−CoGv). Delaying the display on the screen of the monitor significantly diminishes the amplitudes of CoP−CoGv motion whereas CoGh motion is barely affected (Rougier 2004). The main reason lies in the need, in this task, to reduce as much as possible the horizontal acceleration communicated to the CoG. On the other hand, increasing the display scale or zooming the CoP on the screen of the monitor induces the subjects to substantially diminish their CoGh motion without altering the CoP−CoGv motion (Rougier et al. 2004). In this case, the enhanced visibility of the spot improves the capacity of the subject to control his CoG during the correction process. At this stage it should be emphasized that the former is recognized by several authors (e.g. Horstmann and Dietz 1990) as the controlled variable in this task whereas the latter constitutes a fair expression of the resulting joint stiffness (Winter et al. 1998) and is linked to the level of neuro-muscular activity (Rougier et al. 2001). Modeling of the various trajectories as fractional Brownian motion demonstrates that delaying visual information induces subjects to reduce their control of CoP−CoGv motion over the shortest time intervals Δt. On the other hand, enlarging the display scale progressively improves the way the CoGh motion is controlled during the longest Δt. By inducing distinct behavior for each elementary trajectory, these results highlight the need to disentangle complex CoP motion. Complementarily, with one elementary motion (CoP−CoGv or CoGh motion) being controlled at a given time (Rougier and Caron 2000), these results emphasize that undisturbed upright stance is indeed controlled through two different mechanisms that are successively called into play (Collins and De Luca 1993; Rougier 1999a).
On the other hand, as demonstrated by decomposition of the CoP displacements into two elementary CoP−CoGv and CoGh motions, a dysfunction of posture control may have different origins. Overall, an unsteady individual is generally characterized by CoP displacements covering large territories. However, one should bear in mind that these excessive territories can be explained theoretically by either a single exaggerated elementary motion or by both of them. In the first case, the subject will be unable to reduce his/her neuro-muscular activity to a minimum or to control his/her body motion. In the second case, the problem essentially depends on the interaction between the two motions, because, as already stated, the overwhelming CoP−CoGv motion weakens the capacity to handle CoGh displacements efficiently. Naturally, these particularities are also of importance when considering appropriate rehabilitation procedures. Nevertheless, an important question, which remains unsolved and which constitutes the aim of this study, is the compatibility of the effects induced by the VFB technique. It could indeed be of prime interest to improve, for instance, both the neuro-muscular activity (by reducing its control) and body motion (by reinforcing its control) with a single combination of delay and display scale. In particular, could previously observed improved corrections as a result of the enlarged display on the monitor be altered by limiting the horizontal acceleration, and thus the CoG displacements resulting from the delayed VFB? Conversely, could the diminished CoP−CoG motion previously observed when the VFB is delayed be again lessened by enlarging the CoP display? Finally, this pilot study focused on normal subjects to avoid any distortion of the control mechanisms involved in undisturbed stance maintenance because of various possible diseases.
Methods
Experimental procedure
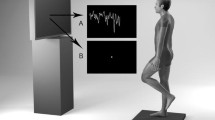
Fourteen healthy subjects, eight males and six females, ranging in age from 21 to 48 years (body weight 65.4±11.5 kg; height 175±7.6 cm; mean±standard deviation) with no known visual or balance pathology gave their written informed consent and were included in this study. As illustrated by Fig. 1, the subjects stood on a triangular force platform of 80 cm each side (PF01; Equi+, Aix les Bains, France) in a natural position (feet abducted at 30°, heels separated by 3 cm) and were asked to reduce the CoP displacements on the screen as much as possible with their arms at their sides. The VFB was implemented by displaying the horizontal trajectory of the CoP in real time through specific software (PROG01; Equi+) on the screen of an additional 21-inch monitor connected to the computer and placed at eye level 80 cm in front of the subjects. The signals issued from the load cells, on which the plate lay, were amplified and converted from analogue to digital form before being recorded on a personal computer with a 64 Hz sampling frequency. The AP and ML displacements of the CoP were depicted on the screen from top to bottom and from left to right, respectively. To facilitate the tracking, the data-acquisition software was programmed in such a way that the spot (4 pixels wide) was always positioned at the center of the screen at the onset of the trial and only the last 64 CoP positions (i.e. 1 s) were represented. The CoP trajectory was then processed in a number of different ways. ML and AP characterize, in the coordinates system, medio-lateral and forward–backward directions, respectively.

Schematic representation of the experimental set-up showing a subject standing on a force platform, beside the investigator, looking at the trajectory of his/her CoP projected on the screen of an additional monitor
The procedure, for the whole sample, included six experiments performed in a random order, all with the common characteristic of involving the visual feedback technique. These six conditions indeed correspond to all possible combinations including two levels of gain (2 and 20) and three time delays (0, 500, and 1,000 ms). Within this framework, the gain or display scale corresponds, in fact, to the ratio expressing the amplitudes of the displacements of the CoP, measured through the force platform, and the corresponding motions on the screen. For instance, a gain of 2 signifies that a CoP displacement of 1 cm will infer a 2 cm displacement of the spot on the monitor. Each condition was composed of five trials lasting 64 s, a rest period of a similar duration being allowed between each trial. At the end of each trial, knowledge of the results in terms of surfaces with a 90% confidence interval of both CoP−CoGv and CoGh motions was given to the subjects (Tagaki et al. 1985). The rest period between each experiment lasted at least 10 min.
Estimation of CoGh and CoP–CoGv motion
CoGh and CoP−CoGv motion were estimated from the CoP trajectories measured through the force platform. To be more precise, a relationship between the amplitude ratio of the horizontal motion of the center of gravity (CoGh), CoP motion (CoGh/CoP), and sway frequencies was used to determine CoGh and consequently CoP−CoGv motion. From this CoGh/CoP relationship, displayed graphically in Fig. 2 and initially proposed by Brenière (1996), it can be logically deduced that CoP oscillations operating over too high frequencies would not incur appreciable CoG movements. This ratio seems maximum for the lower frequencies (CoGh and CoP are characterized by similar positions at 0 Hz) and tends towards zero above 3 Hz. Estimation of the CoGh consists in multiplying the complex data, transformed in the frequency domain through a fast Fourier transform (FFT), by the above-mentioned low-pass filter and recovering to the time domain by processing an inverse FFT. All of this data processing was automatically performed through the Equi+ PROG01 software. The article by Caron et al. (1997) provides the details of this procedure.

Amplitude ratio between CoGh and CoP motion as a function of sway frequency. This filter shows that CoGh and CoP have similar positions at 0 Hz whereas CoGh motion relative to CoP becomes smaller, thus determining increased CoP−CoGv amplitudes, as displacement frequency increases (from Rougier and Caron 2000)
Signal processing
The signal processing used in this study has been described in detail in previous studies (Rougier and Caron 2000; Rougier and Farenc 2000). Three approaches were adopted to study the CoP−CoGv and CoGh elementary motion:
-
1)
a classical method based on data such as the surface of an ellipse calculated with a confidence interval (Tagaki et al. 1985) and mean velocity,
-
2)
data (root mean square (RMS) and mean power frequency (MPF)) calculated from mean frequency decompositions of the various motions on specific bandwidths (0–3 Hz for CoP−CoGv and 0–0.5 Hz for CoGh), and
-
3)
a mathematical model, termed fractional Brownian motion (Mandelbrot and Van Ness 1968).
The RMS and the MPF express a mean amplitude independent of the frequency bandwidth, and the respective contribution of lowest and highest frequencies, respectively. Fractional Brownian motion (fBm) modeling enables specification of the extent to which a trajectory is controlled. Its general principle is that the aspect of a trajectory, expressed as a function of time, may be quantified by a fractional, i.e. non-finite, integer space dimension. The latter thus provides a quantitative measurement of wiggle in the trajectory in a single direction. This fractional dimension D, in a single direction, is, in fact, linked to the Hurst scaling exponent H, because D=1−H for this case. This scaling regime corresponds graphically to the half slope of the line portions constituting a variogram depicted bi-logarithmically. The latter in fact expresses the mean-square displacements <Δx2> as a function of increasing time intervals (1/64 s<Δt<10 s) and is given by the formula:
Note that the squared distances are simply used to prevent a null mean displacement. A median value of 0.5 for H indicates a linear relationship between the two variables and thus a lack of correlation between past and future increments, suggesting that the trajectory is totally uncontrolled. On the other hand, i.e. if H differs from 0.5, positive (0.5<H) or negative (H<0.5) correlation can be inferred, which is indicative of a given part of determinism in the control. Depending on how H is positioned with respect to the median value 0.5, it can be inferred that the trajectory is more or less controlled—the closer to 0.5, the less the control. In addition, depending on whether H is superior or inferior to the 0.5 threshold, persistent (the point is drifting away) or anti-persistent behavior (the point retraces its steps) can be revealed, respectively.
Because two straight-line portions generally characterize variograms relative to undisturbed upright stance maintenance (a quite flat line preceding or succeeding a steeper one), a final step consists in the determination of the transition point for both directions, i.e. the point corresponding to the slope inflection. CoP and CoGh displacements being by definition in phase, the temporal coordinate of the transition points on the variograms characterizing the CoP trajectories will also be that of the CoGh and CoP−CoGv motion. The method used for this purpose is based on the evolution, as a function of increasing Δt, of the distance between the CoP variogram and a completely stochastic process. In this case, the stochastic process corresponds to the ordinary Brownian motion and is characterized by a 1 slope (inferring, in that case, a 0.5 value for H). The retained principle is that the Δt for which this distance is maximum is the Δt of the transition point (Rougier 1999b). However, it should be pointed out that the stochastic behavior, taken as a reference, is itself modified by the low or high-pass filter used for the computation of CoGh and CoP−CoGv motion. As seen in the upper part of Fig. 3, instead of the straight line characterizing the CoP trajectories, the filter effect leads, in fact, to curvilinear functions moving progressively away from or closer to the 1 slope mentioned above (because 2×(H=0.5)=1). Precisely, the temporal order of the increments from CoP trajectories have been randomly shuffled and then recombined to generate stochastic trajectories which were then processed like the experimental ones to obtain CoGh and CoP–CoGv motion. Scaling regimes relative to “average stochastic variograms” over the same Δt must therefore be taken as a reference in such a way that:
where Hcal, Hexp, and Hsto represent the calculated, experimental, and stochastic scaling regimes, respectively. Thus, for each of the two motions investigated and each ML and AP component, two scaling exponents (indexed as short and long latencies: Hsl and Hll) and the coordinates of the transition point were extracted. Figure 3 illustrates this procedure with an example of CoP, CoGh, and CoP−CoGv trajectories.

Method used to determine, in one direction, the transition between successive scaling regimes for the CoGh and CoP−CoGv trajectories from one subject’s data. The upper part represents the three variograms, computed for a single direction ML or AP, characterizing CoP (gray) and CoGh and CoP–CoGv (black), i.e. the mean-square displacements as a function of increasing Δt. The tilted or curvilinear dashed curves which pass through each variogram at the point with the shortest Δt theoretically express a pure stochastic process. The lower part displays the distances, in arbitrary units, between variograms and the dashed lines mentioned above. The maximum distance relative to the CoP trajectory is taken to be the transition points’ Δt coordinate. Thus, the dotted horizontal and vertical straight lines correspond to the mean square displacement and Δt coordinates of the different transitions between successive regimes involved in undisturbed upright stance control (from Rougier and Caron 2000)
To evaluate the effects of the interaction between time delays and scaling display of the VFB on undisturbed stance control, all data were analyzed by two-factor analysis of variance (ANOVA) with repeated measures, the first level of significance being set at P<.05.
Results
Before presenting the results of both types of elementary motion, it is relevant to mention that modifying the two FBV variables does not affect the mean position of CoGh motions in both ML and AP directions. Consequently, the various effects observed through this investigation cannot be explained by modification of the position of the body with respect to the base of support.
Classical data
An interesting result is that the ANOVA is incapable of displaying the smallest effect when only some of the data obtained from the CoP displacements are considered (left column of Fig. 4). This is true for both the surfaces of the ellipses and the mean velocities. On the other hand, opposite effects can be observed for both CoP−CoGv and CoGh motion. One may note, in particular, for the surfaces of the ellipses that modifying the VFB gain has an effect on the CoGh motion (F(1,78)=5.906, P<.017) whereas the time delay interferes principally with the CoP−CoGvmotions (F(2,78)=4.285, P<.017). Overall, increasing the VFB delay induces an increase of CoGh motion and a decrease of CoG−CoPv motion. Because the CoP displacements constitute the algebraic sum of these two elementary motions, the two previous effects, of opposite signs, tend to counterbalance each other.

Group means and standard deviations for classical and frequency data displayed as a function of increasing time delay. In each panel, the values are in black and gray for gains of 2 and 20, respectively. The significant effects assessed by the ANOVA and the nature of the data involved are presented above or below the plots (***P<.001; **P<.01; *P<.05). Note the differences according to the motion involved: although the CoP (left column) does not provide statistically significant results, CoGh (middle column) and CoP−CoGv (right column) give some significant results: the former mainly by modifying the gain and in some cases the delay, the latter by modifying solely the delay
On the other hand, an insignificant effect is seen for the mean velocities. As the delay increases, these velocities seem to diminish for all motion. Modifying the gain does not seem to alter this feature even though these velocities, for a given delay, are always lower for the CoGh motions when the gain is reduced.
Frequency data
With regard to the RMS data, some opposing effects are seen depending on the elementary motion taken into consideration (lower part of Fig. 4). From that, the RMS values computed from the CoP displacements are unaffected by the gain or delay modifications. On the other hand, the former plays a significant role for the CoGh motion for both ML (F(1,78)=4.491, P<.037) and, in particular, AP directions (F(1,78)=7.404, P<.008). The larger the visual display of the VFB, the smaller the CoGh motion in these directions. On the other hand, the CoP−CoGv motion tends to diminish with the setting of a VFB delay, whatever the gain values. This feature concerns both ML (F(2,78)=3.445, P<.037) and AP directions (F(2,78)=4.151, P<.019).
The MPF values, uniquely, elicit statistically significant results for the elementary CoGh motion only, as seen from the lower panels of Fig. 4. Increasing the VFB gain induces some enhanced values for these MPF in both ML and AP directions. Complementarily, significantly delaying the VFB produces a decrease of these values. Interestingly, one may emphasize that the F values are higher when the VFB gain is modified (ML: (F(1,78)=15.979, P<.000); AP: (F(1,78)=50.589, P<.000) than when a delay is set (ML: (F(2,78)=9.873, P<.000); AP: (F(2,78)=6.236, P<.003). Finally, opposing effects, although without any statistical trend, are observed for the CoP−CoGv motion.
Data from the fBm modeling
Modifying the display scale and/or delaying the VFB on the screen alters the postural behavior of the subjects, as highlighted by the fBm modeling. If the temporal coordinate Δt, which indicates the onset of the corrective process, remains unaffected by the VFB, the spatial coordinates <Δx2> have a variety of statistically significant effects. However, these latter are quite opposite in nature depending on the direction. One may note indeed, from the ANOVA, that modifying the delay induces smaller <Δx2> in the ML (F(2,78)=4.814, P<.011) and, more particularly, in the AP direction (F(2,78)=6.835, P<.002) for the CoP–CoGv motion (Fig. 5). The same feature is also observed for the CoP displacement, but solely in the ML direction (F(2,78)=4.692, P<.012). On the other hand, displaying the VFB through different scales results in appreciable modification in the AP direction alone. This is true for both CoP (F(1,78)=4.827, P<.031) and CoGh motion (F(1,78)=4.335, P<.041).

Group means and standard deviations for data obtained from fBm modeling displayed as a function of increasing time delay. As for Fig. 4, significant effects assessed by ANOVA and the nature of the data involved are presented above or below the plots (***P<.001; **P<.01; *P<.05). Note the most striking effects on the long latency scaling regimes for both CoP (left column) and CoGh (middle column) motion which occur particularly in the AP direction. Note also that the main effect of delaying the display, at least for the ML direction, is lessened control of the CoP−CoGv motion and reduced spatial transition point coordinates <Δx2>, as revealed through scaling regimes Hsl (right column)
Finally, some effects on the slopes of the variograms should also be emphasized. To be precise, these slopes, as mentioned in the methods section, are used to compute the various scaling regimes aimed at characterizing the successive line portions usually depicted in the variograms for such signals. From our results it seems that the observed effects are not identical for the two directions. In the ML direction, modification of the delay provokes a decrease of the short latency scaling regimes Hsl for both CoP−CoGv (F(2,78)=6.534, P<.002) and CoP displacements (F(2,78)=4.768, P<.011). As seen from Fig. 5, the longer the delay, the smaller the Hsl values. Conversely, modifying the gain has no particular effect, even though some slight differences can be observed for the long scaling regimes Hll computed from the CoGh motion. In the AP direction, changing the delay only induces effects on the long latency scaling exponents Hll for the global CoP displacements (F(2,78)=4.287, P<.017). It is worth noting from the lower panels of Fig. 5 that smaller Hll, which are supposed to reveal an enhanced control of this motion, are found for the 500 ms delay. Finally, altering the gain of the VFB influences the values of the scaling regimes Hll for both CoGh (F(2,78)=80.230, P<.000) and CoP motion (F(2,78)=2.458, P<.000). As seen from the plots of Fig. 5, the larger the gain of the VFB, the smaller the Hll values.
Interestingly, all the tests conducted on the interaction between time delay and display scale do not lead to any statistically significant results for any of the classical, frequency or fBm data.
Discussion
The results of this study clearly indicate that displaying visual feedback to healthy individuals leads them to adopt specific postural behavior depending on the characteristics of the feedback. On the whole, the results are in complete accordance with the two previous studies on the subject and serve to confirm that:
-
1)
displaying visual feedback through a monitor screen with a time delay causes the individual to reduce the amplitude of the CoP−CoGv motion (Rougier 2004) on the one hand, and
-
2)
magnifying the scale of the visual display predominantly affects the CoGh motion (Rougier et al. 2004).
The novelty of this study’s results is that these effects remain even though another aspect of VFB, i.e. display scale or time delay, is modified. In other words, these data indicate that the reduced CoP−CoGv motion usually observed during delayed VFB is still present, irrespective of the scale of CoP visualization. Concomitantly, the effect of the gain (i.e. the improved control for correcting CoGh motion) is preserved throughout the various time delays. Another interesting trend highlighted by the results is that both classical (i.e. based on summary statistics) and frequency analyses applied to the complex CoP trajectories do not reveal any statistically significant trend. On the whole, these features thus emphasize the importance of having recourse to data obtained from fractional Brownian motion modeling and/or focusing on elementary CoGh and CoP−CoPv motion.
Modifying the time delay predominantly affects the way CoP−CoGv motion is controlled
When graphically displaying the variograms to illustrate the relationship between the mean square distances <Δx2> and the increasing time intervals Δt, one of the main modifications concerns the slopes of the linear portions computed for the shortest Δt. As seen by the scaling regimes Hsl, whose values express the half slopes of these line portions, setting a time delay in the visualization of the VFB, whatever the value of the gain, induces a decrease in these slopes and consequently reduced Hsl values. From the fBm modeling, these latter express a larger contribution of stochastic processes in the control of CoP−CoGv motion. Because the latter motion, during these shortest Δt, constitutes the main part of the variograms of the complex CoP trajectories, similar trends can be found for the Hsl scaling coefficients aimed at characterizing these CoP variograms. The slight difference comes from the fact that the CoGh components, which seem uncontrolled in all such postural studies during this period (because the Hsl scaling exponents are close to the median 0.5 value), are suppressed when studying the CoP−CoGv motions. Biomechanically, these reduced Hsl can be interpreted as a larger probability for the trajectories to wiggle until a corrective process begins to operate. In other words, for a given time interval, the distance covered by the trajectory in a single direction will be lessened. This principle explains why the spatial coordinates of the transition points are also reduced under these conditions. One should note, at this stage, that increasing the gain induces a tendency for the Hsl regimes to be reduced. It can be thus hypothesized that the improved sensitivity of the visualized CoP trajectories enables the subjects to better appreciate the advantage of diminishing the CoP−CoGv motion as much as possible, and from that to better appreciate the neuro-muscular activity in his lower limbs. These effects seem to be progressive in nature for the ML direction whereas the opposite is true in the AP direction for the longest delay of 1 s. This tendency for effects to be more significant the ML direction than in the AP direction is also a common trend in this kind of study. For instance, it has previously been shown that the VFB effects, when compared to a referenced eyes-open condition, were always more pronounced in this direction (Rougier 2003a). Similarly, it is also in the ML direction that the Hsl scaling exponents are reduced when the time delay is progressively increased (Rougier 2004). Accordingly, some future experiments consisting in furnishing information relative to a single direction should be planned to better assess these different effects.
On the other hand, it should be borne in mind that the CoP−CoGv amplitudes are thought to express the level of neuro-muscular activity developed for this equilibrium task. According to the size principle initially described by Henneman et al. (1965), the muscular force increase is achieved by recruiting additional motor units whose mechanical properties in terms of contraction times tend to progressively decrease. From the CoGh/CoP ratio, the higher the frequency, the smaller the relative displacements of CoGh and thus, the larger the amplitudes and the higher the MPF of the CoP−CoGv spectra (Winter et al. 1998; Caron et al. 2000). Thus, one of the main consequences of setting a delay could be a reduced muscular activity in the subjects.
Despite this reduced initial horizontal acceleration communicated to the CG, the correction of the latter seems to be initiated after a similar distance has been covered when the delay is lengthened. This is indeed highlighted by the <Δx2> coordinates. However, one should note that neither the temporal coordinates Δt nor the degree to which the CoGh motions are controlled in order to return to their initial position, are affected by the time delay.
Modifying the display scale predominantly affects the way the CoGh motions are controlled
The main advantage for the individual to see the displacements of its CoP on a larger scale is that the CoGh motion becomes further reduced in amplitude, as seen, for instance, by the surface or RMS data. This effect is seen for both ML and AP directions, even though a more pronounced reduction is, a priori, observed for the latter. Another interesting result concerns the MPF which seem to increase with the gain, signifying that the delay needed by the CoGh to return to its initial position is diminished. These modifications in postural strategies are plainly explained by fBm modeling. Indeed, even though a slight effect is observed for the spatial coordinates of the transition points in the AP direction, the main effect remains a huge diminution of the long latency scaling exponents for both ML and AP directions. In other words, these features show that when the display scale is high, the individuals become more able to control their CoGh motions in order to make it return to a place where body acceleration is temporally lowered. This feature alone could explain the reduced MPF mentioned above. On the other hand, the previous effects induced by an increased gain appear to be unaffected by the setting of the time delay. This suggests that the individuals, even though the given information is delayed, are still able benefit from informed knowledge of the CoP position. On the whole, these results are in complete accordance with previous results which describe that these postural effects were progressively observed as the display scale was increased (Rougier et al. 2004). Lastly, because the CoGh motion constitutes the main contribution of the CoP trajectories during the longest Δt, as seen through the variograms computed from the various displacements (Fig. 3), these effects on the scaling regimes Hll are also, logically, found for the CoP displacements.
Despite reduced initial horizontal acceleration, the CoGh is not necessarily better handled to maintain balance during VFB
As shown by the results, the amplitude of CoP−CoGv motion is reduced when the VFB is delayed. Considering that, biomechanically, this latter motion expresses the initial horizontal acceleration communicated to the CoG (Brenière et al. 1987), one may thus hypothesize that the control of body motion should, in turn, be facilitated. One of the most interesting findings of modeling the trajectories (issued from a force platform) as an fBm is that the CoGh motion seems to be controlled only for the longest Δt. The previous period, the shortest Δt, is stochastic in nature in all cases (Rougier and Caron 2000). Consequently, the only theoretical possibility for modulating the control characteristics of this motion includes the spatio-temporal coordinates of the transition point (<Δx2> and Δi) and the extent to which the CoGh motion is corrected (Hll). One of the main characteristics of the VFB technique, when compared with an eyes-open condition, is to induce an increase in the distance covered by the CoGh motion until correction occurs (Rougier 2003a). Because this feature is observed whatever the delay or the display scale, this would seem to suggest that the ability to limit the distance covered until the onset of the corrective mechanism is not related to the forces applied on the CoG at this precise moment. This would suggest that a minimum distance needs to be covered before engaging the corrective process and/or the CNS needs to integrate other perceptual cues from the sensory receptors. This differentiation thus legitimizes the value of computing these mean square distances for both CoP−CoGv and CoGh motion and, on the whole, validates the dissociation of the CoP trajectories into two elementary motions. Consequently, even though both elementary CoP−CoGv and CoGh motion is estimated on a similar basis, i.e. the trajectories of the CoP, the postural behavior encountered in this study demonstrates that controls of these two motions are completely separate from one another and thus establishes the need to disassociate the CoP. These results also emphasize that neither CoG nor CoP−CoG motion can be viewed as the single variable controlled in this undisturbed upright stance experiment, thus challenging the hypotheses formulated for instance by Horstmann and Dietz (1990) for the former and Winter et al. (1998) for the latter. As shown by the variograms, each elementary motion is indeed the controlled variable but for a given time and in alternating fashion.
Another interesting result of this study is the nature of the links between these distances and the extent to which the CoGh motion is then controlled during the longest Δt. In most of the studies conducted on this topic and analyzed by use of the same methodology, one of the common features generally observed is that the longer this distance, the more anti-persistent this process, and vice versa (Rougier 2003a, 2003b; Rougier et al. 2004). The decreased tendency for the CoGh motions to wiggle as the scaling display increases and the unchanging distance from which this control operates highlights the independence of these variables.
Rehabilitation applications of the VFB technique with display scale and time delay data
The complementary nature of the postural behavior induced by this technique makes it a promising tool for rehabilitation purposes. Indeed, apart from the incapacity to properly initiate postural correction after a reduced distance has been covered, whatever the cause of the balance problem (excessive muscular activity, difficulty, on a regular basis, to bring the CoG back to a position where the external forces acting upon it will be reduced), VFB with these two variables is thought to provide a promising way forward for most patients. More pragmatically, modulating the display scale and time delay at the same time, might be time-saving for patients needing to reduce their neuro-muscular activity (by delaying the VFB) whilst at the same time improving their capacity to appropriately correct their body motions. Further investigations are, however, needed to assess whether these results, obtained with healthy adults, can be observed as clearly with people with various pathologies or with elderly individuals.
References
Brenière Y (1996) Why we walk the way we do? J Mot Behav 28:291–298
Brenière Y, Do MC, Bouisset S (1987) Are dynamic phenomena prior to stepping essential to walking? J Mot Behav 19:62–76
Caron O, Faure B, Brenière Y (1997) Estimating the center of gravity of the body on the basis of the center of pressure in standing posture. J Biomech 30:1169–1171
Caron O, Gélat T, Rougier P, Blanchi JP (2000) A comparative analysis of the centre of gravity and the centre of pressure sway paths in standing posture: an estimation of active stiffness. J Appl Biomech 16:234–247
Collins JJ, De Luca CJ (1993) Open-loop and closed-loop control of posture: a random-walk analysis of center of pressure trajectory. Exp Brain Res 95:308–318
Henneman E, Somjen G, Carpenter DO (1965) Functional significance of cell size in spinal motoneurons. J Neurophysiol 28:560–580
Horstmann A, Dietz V (1990) A basic posture control mechanism, the stabilization of the center of gravity. Electroencephalogr Clin Neurophysiol 76:165–176
Mandelbrot BB, Van Ness JW (1968) Fractional Brownian motions, fractional noises and applications. SIAM Rev 10:422–437
Rougier P (1999a) Influence of visual feedback on successive control mechanisms in upright stance in humans assessed by fractional Brownian motion modelling. Neurosci Lett 266:157–160
Rougier P (1999b) Automatic determination of the transition between successive control mechanisms in upright stance assessed by modelling of the centre of pressure. Arch Physiol Biochem 107:35–42
Rougier P (2003a) Visual feedback induces opposite effects on elementary centre of gravity and centre of pressure minus centre of gravity motions in undisturbed upright stance. Clin Biomech 18:341–349
Rougier P (2003b) The influence of having the eyelids open or closed on undisturbed postural control. Neurosci Res 47:73–83
Rougier P (2004) Optimising the visual feedback technique for improving upright stance maintenance by delaying its display: behavioural effects on healthy adults. Gait Posture 19:154–163
Rougier P, Caron O (2000) Center of gravity motions and ankle joint stiffness control in upright undisturbed stance modeled through a fractional Brownian motion framework. J Mot Behav 32(4):405–413
Rougier P, Farenc I (2000) Adaptative effects of loss of vision on upright undisturbed stance. Brain Res 871:165–174
Rougier P, Burdet C, Farenc I, Berger L (2001) How leaning backward or forward affects undisturbed stance control in humans. Neurosci Res 41(1):41–50
Rougier P, Farenc I, Berger L (2004) Modifying the gain of the visual feedback affects undisturbed upright stance control. Clin Biomech 19:858–867
Tagaki A, Fujimara E, Suehiro S (1985) A new method of statokinesigram area measurement: application of a statistically calculated ellipse. In: Igarashi M, Black O (eds) Vestibular and visual control on posture and locomotor equilibrium. Karger, Basel/Suisse, pp 74–79
Weinstein S (1968) Intensive and extensive aspects of tactile sensitivity as a function of body part, sex, and laterality. In: Kenshalo DR (ed) The skin senses. Thomas, Springfield IL USA, pp 195–222
Winter DA, Patla AE, Prince F, Ishac M, Gielo-Perczak K (1998) Stiffness control of balance in quiet standing. J Neurophysiol 80:1211–1221
Acknowledgments
The author is pleased to acknowledge D. Goodhew for correction of the English text and the referees for their helpful comments
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Rougier, P. Compatibility of postural behavior induced by two aspects of visual feedback: time delay and scale display. Exp Brain Res 165, 193–202 (2005). https://doi.org/10.1007/s00221-005-2288-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-005-2288-8