
Abstract
It has been argued that representations of peripersonal space based on haptic input are systematically distorted by egocentric reference frames. Interestingly, a recent study has shown that noninformative vision (i.e., freely viewing the region above the haptic workspace) improves performance on the so-called haptic parallel-setting task, in which participants are instructed to rotate a test bar until it is parallel to a reference bar. In the present study, we made a start at identifying the different sensory integration mechanisms involved in haptic space perception by distinguishing the possible effects of orienting mechanisms from those of noninformative vision. We found that both the orienting direction of head and eyes and the availability of noninformative vision affect parallel-setting performance and that they do so independently: orienting towards a reference bar facilitated the parallel-setting of a test bar in both no-vision and noninformative vision conditions, and noninformative vision improved performance irrespective of orienting direction. These results suggest the effects of orienting and noninformative vision on haptic space perception to depend on distinct neurocognitive mechanisms, likely to be expressed in different modulations of neural activation in the multimodal parietofrontal network, thought to be concerned with multimodal representations of peripersonal space.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Representations of peripersonal space based on haptic input are systematically distorted, as has been demonstrated with haptic matching tasks: blindfolded subjects instructed to rotate bars until they are parallel or collinear make matching errors that vary systematically in magnitude and direction throughout peripersonal space (e.g., Kappers and Koenderink 1999; Kappers 2002). These errors have been shown to result from the use of biasing egocentric reference frames where an allocentric one should have been used (Zuidhoek et al. 2003). Interestingly, a recent study by Newport and colleagues (2002) demonstrated that freely viewing the region of space directly above the workspace reduced deviation size in haptic parallel-setting. The way in which this “noninformative” visual information exerted its effect is by no means clear, however. For one, considering that participants were allowed to freely move their heads and eyes during the task, it may be that orienting behavior during the task affected multimodal integration. In support of this, simple tactile detection and discrimination studies have shown that the orienting of head and eyes towards a tactilely stimulated body site without any visual input boosts tactile processing, which suggests hardwired multimodal connections between tactile, visual and proprioceptive processing (Honoré et al. 1989; Driver and Grossenbacher 1996).
The goal of the current study was to distinguish the possible effects yielded by orienting mechanisms from those of noninformative vision. The two factors might either show mutual dependencies or have independent effects. Discovering their relationship will give further insight in the mechanisms of multimodal integration and crossmodal facilitation. To this end, we examined the role of orienting direction in the bimanual haptic perception of parallelity of two bars in the horizontal plane, in no-vision and noninformative vision conditions. We reasoned that if the orienting direction of head and eyes affects haptic space perception, orienting to either of the hands handling the bars may improve the parallel-setting performance by enhancing tactile perception and possibly also proprioception (cf. Newport et al. 2001, but see also Van Beers et al. 1999). A neutral fixation point straight ahead above the workspace served as a baseline.
Materials and methods
This study has been approved by the local ethics committee and was performed in accordance with the Declaration of Helsinki of 1964.
Participants
Twelve paid undergraduates of Utrecht University (six male, six female, aged 18–30 years) participated in the experiment, after having given their informed consent. All were right-handed as assessed by means of a standard questionnaire (Annett 1970).
Apparatus and stimuli
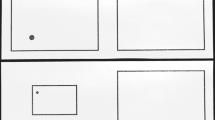
See Fig. 1. The participants were seated 3–5 cm from the middle of the long end of a table (160×80 cm). Two boards (28×28×0.9 cm) showing a protractor (diameter 20 cm) were fixed to the tabletop. The centers of the protractors were 14 cm from the table’s edge, which is closer than in previous studies (20.5 cm). Each protractor contained an aluminum bar (20×1.1 cm) that could be rotated around the center of the protractor. The distance between the centers of the bars was 120 cm. The bar to the left of the participant served as the reference bar. This bar was presented in one of eight orientations: 18, 42, 66, 84, 108, 132, 156, and 174° (0° being parallel to the long side of the table and increasing values signifying a rotation in counterclockwise direction). In each trial, the bar to the right—the test bar—was to be matched to the reference bar orientation by rotating it with the right hand. The test bar was presented in a random orientation, i.e., corresponding to the orientation of the seconds-hand of a clock at that particular time. In every condition, each reference bar orientation was presented three times, adding up to 24 trials per condition. The order of reference bar presentation was randomized and different for each participant.

Schematic top view of the experimental setup in noninformative vision conditions: an opaque cloth (transparent in the picture) covered participants’ shoulders and limbs, as well as the workspace. b Side view of the experimental setup in noninformative vision conditions
The experiment consisted of six conditions: three orienting directions (the “reference hand” (i.e., the hand handling the reference bar), a central “neutral” fixation point, the “test hand” (i.e., the hand handling the test bar)) in no-vision and noninformative vision situations. Orienting meant directing head and eyes. In no-vision conditions, participants were blindfolded. In noninformative vision conditions, an opaque cloth covered the workspace (20 cm above the table) as well as the participant’s limbs and shoulders (about 35–40 cm above the table), so that participants viewed the region of peripersonal space directly above the workspace (see Fig. 1b.). Conditions were blocked. Each participant was presented with a different sequence of conditions.
Procedure
Participants were instructed to use their right hand to rotate the test bar parallel to the reference bar, which they simultaneously felt with their left hand. Before the start of every trial they were instructed to orient head and eyes in the direction corresponding to the condition at hand. One experimenter monitored the direction of head and eyes. Of course, in blindfolded conditions, the direction of the eyes could not be observed. Note, however, that with the head directed at one of the fixation points, it was impossible for the participant to direct the eyes to one of the other fixation points due to the large distance between fixation points.
After having oriented head and eyes to the appropriate location, participants were allowed to touch and explore the bars freely, as long as they did not change the orientation of the reference bar, did not touch the edges of the table, and remained seated. The time per trial was restricted to 10 s, which proved to be more than sufficient. In between trials, participants were to place their hands right in front of them on the table and to look straight up. The participants never received any feedback on their performance.
Data analyses
For all analyses below we computed signed average errors in degrees, assigning a positive value to deviations in the expected direction (i.e., the direction of the systematic deviations found in previous parallel-setting studies), and a negative value to deviations in the opposite direction.
Results
A 3 (orienting direction) × 2 (input condition, i.e., no vision vs. noninformative vision) within-subjects ANOVA was conducted for the mean signed errors. Significant main effects were found for orienting direction (F (2,22)=5.1, p=.016) and input condition (F (1,11)=37.9, p<.001). The main effect of orienting direction was expressed in different average deviations for the three orienting directions: 54.1, 56.7, and 59.9° for orienting to the reference hand, central fixation point and the test hand, respectively. Post-hoc testing, with Bonferroni correction lowering significance criterion α to 0.017, showed that performance with reference hand orienting was significantly better than performance with test bar orienting: t (11)=−3.7, p=.004. Differences for reference hand and test hand orienting with respect to central fixation point orienting were not significant: t (11)=−1.4, p=.2 and t (11)=−1.7, p=.11, respectively. Furthermore, the average deviations for the three orienting directions showed a significant linear trend (F (1,11)=13.9, p=.004), implying that the facilitating effect of orienting decreases with absolute distance from the reference hand.
The main effect of input condition signified that allowing the processing of noninformative vision improved performance (F (1,11)=37.9, p<.001): it reduced deviation size from 59.5 to 54.3°. Importantly, orienting direction and input condition showed no interaction (F (2,22)=.19, p=.83), yet an additive relation (see Fig. 2), implying that orienting direction and noninformative vision affect haptic spatial performance independently.

Orienting direction of head and gaze affect haptic parallel-setting in both no-vision and noninformative vision conditions. Error bars represent ± standard error of mean
Discussion
The primary goal of the present study was to identify multimodal mechanisms involved in constructing representations of peripersonal space from haptic input. We focused on distinguishing possible effects yielded by orienting mechanisms from those of the processing of visual input that was not directly relevant for the task (‘noninformative vision’). To this end, we presented participants with a bimanual parallel-setting task, in which participants were instructed to orient head and eyes towards either the reference hand, the test hand, or a neutral fixation point, in both a no-vision and a noninformative vision condition.
We found that orienting direction affected the perception of space based on haptic input: orienting towards the hand handling the reference bar yielded significantly smaller deviations than orienting towards the test hand. Moreover, the average deviations for the three orienting directions showed a significant linear trend, suggesting that the facilitating effect of orienting decreases with distance from the point of fixation to the reference hand. Furthermore, we observed that the availability of noninformative visual input facilitated parallel-setting performance, a finding that replicates Newport and colleagues (2002). Importantly, the effects of orienting direction and input condition (noninformative vision vs. no vision) proved to be independent, suggesting distinct neurocognitive mechanisms. An additional observation is that the deviations found were relatively large as compared to those in previous studies. This can be explained by the relatively extreme placing of the stimuli, with the bars 120 cm apart and closer to the body than in previous studies. This led to large orientation differences between the hands, which has been suggested to be the main cause of deviations in haptic orientation perception (Zuidhoek et al. 2003).
An interesting question, of course, is how orienting direction and noninformative vision contribute to the representation of haptic space. Noninformative vision may improve haptic perceptions of peripersonal space by providing information about the space between and beyond the bars (although above the actual workspace), and as such serving as an allocentric reference frame. At the neural level of multimodal representation, noninformative vision then may be reflected in the increase of the number of multimodal cells activated in a parietofrontal multimodal network suggested to mediate the coding of peripersonal space and our limbs in it by integrating visual, proprioceptive, tactile and possibly also auditory inputs (Làdavas et al. 1998; Obayashi et al. 2000; Newport et al. 2001; Graziano 2002; Lloyd et al. 2003).
With respect to the effect of orienting direction, the current findings showed that—although the task comprised the perception of two bars—orienting towards the reference bar enhanced performance, whereas orienting towards the test bar did not. Importantly, this implies that the mere alignment of sensory systems through orienting is not sufficient to result in an improvement in (spatial) perception (cf. Kennett et al. 2001). Rather, it seems that orienting enhances perception by facilitating additional processing, which—in the parallel-setting task—is performed exclusively on reference hand input. Two underlying mechanisms may be considered here. First, visual imagery has been suggested to play a role in improving allocentric representations of space based on haptic input (Zuidhoek et al. 2003). It may be that performing the parallel-setting task comprises imagery of the reference bar which is stimulated and/or facilitated by orienting towards it. Second, orienting may facilitate the allocation of attentional resources (Honoré et al. 1989; Driver and Grossenbacher 1996). Then, the improvement in performance with reference hand orienting may reflect the attentional focus being primarily on the reference hand during task performance. Possibly, these two mechanisms are interlinked: imagery of the reference hand may depend on attentional resources, and in turn, attending a (visuo-tactile) image of the limb itself may be critical, since visually attending a neutral object at the location of tactile stimulation has been found not to boost perception (Kennett et al. 2001; Taylor-Clarke et al. 2002).
It is unknown how the facilitating effect of orienting on haptic perception is mediated at a neural level. However, Taylor-Clarke and colleagues (2002) have argued that the effects of vision on tactile perception are reflected by a modulation of cell activity in the primary and secondary somatosensory cortex (SI and SII), brought about by back projections from multimodal cortical areas. Importantly, they showed that S1 processing (N80 component) is only enhanced by vision when tactile stimulation is task-relevant, suggesting top-down attentional selection. It might be that the contribution of visual imagery operates in a similar manner, i.e., via back projections from those multimodal areas concerned with visuo-tactile imagery of our moving limbs in space like the intraparietal sulcus (Obayashi et al. 2000; Lloyd et al. 2003) to S1 and S2, with those to S1 depending on task-dependent selection processes.
References
Annett M (1970) A classification of hand preference by association analyses. Br J Psychol 61:303–321
Driver J, Grossenbacher PJ (1996) Multimodal constraints on tactile selective attention. In: Inui T, McClelland JL (eds) Attention and performance XVI. MIT Press, Cambridge, MA, pp 209–235
Graziano MS (2002) Where is my arm? The relative role of vision and proprioception in the neuronal representation of limb position. Proc Natl Acad Sci USA 96:10418–10421
Honoré J, Bourdeaud’hui M, Sparrow L (1989) Reduction of cutaneous reaction time by direction eyes towards the source of stimulation. Neuropsychologia 27:367–371
Kappers AML (2002) Haptic perception of parallelity in the midsagittal plane. Acta Psychol 109:25–40
Kappers AML (2003) Large systematic deviations in a bimanual parallelity task: further analysis of contributing factors. Acta Psychol 114:131–145
Kappers AML, Koenderink JJ (1999) Haptic perception of spatial relations. Perception 28:781–795
Kennett S, Taylor-Clarke M, Haggard P (2001) Noninformative vision improves the spatial resolution of touch in humans. Curr Biol 11:1188–1191
Làdavas E, Di Pellegrino G, Farnè A, Zeloni G (1998) Neuropsychological evidence of an integrated visuotactile representation of peripersonal space in humans. J Cog Neurosci 10:581–589
Lloyd DM, Shore DI, Spence C, Calvert GA (2003) Multisensory representation of limb position in human premotor cortex. Nat Neurosci 6:17–18
Newport R, Hindle JV, Jackson SR (2001) Links between vision and somatosensation: vision can improve the felt position of the unseen hand. Curr Biol 11:975–980
Newport R, Rabb B, Jackson SR (2002) Noninformative vision improves haptic spatial perception. Curr Biol 12:1661–1664
Obayashi S, Tanaka M, Iriki A (2000) Subjective image of invisible hand coded by monkey intraparietal neurons. Neuroreport 11:3499–3505
Taylor-Clarke M, Kennett S, Haggard P (2002) Vision modulates somatosensory cortical processing. Curr Biol 12:233–236
Van Beers RJ, Sittig AC, Denier van der Gon JJ (1999) Localisation of a seen finger is based exclusively on propioception and on vision of the finger. Exp Brain Res 125:43–49
Zuidhoek S, Kappers AML, Van der Lubbe R, Postma A (2003) Delay improves performance on a haptic spatial matching task. Exp Brain Res 149:320–330
Acknowledgement
This research was supported by a grant from the Netherlands Organization for Scientific Research (NWO: 440-20-000).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Zuidhoek, S., Visser, A., Bredero, M.E. et al. Multisensory integration mechanisms in haptic space perception. Exp Brain Res 157, 265–268 (2004). https://doi.org/10.1007/s00221-004-1938-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-004-1938-6