
Abstract
Rationale
Neuropeptide Y (NPY) and its receptors are densely localized in brain regions involved in the mediation and modulation of fear, including the amygdala. Several studies showed that central NPY is involved in the modulation of fear and anxiety.
Objectives
In the present study, we investigated (1) whether intra-amygdala injections of NPY affect the expression of conditioned fear and (2) whether NPY Y1 receptors (Y1R) mediates the effects of these intra-amygdaloid NPY injections.
Results
Intra-amygdala NPY injections robustly decreased the expression of conditioned fear measured by conditioned freezing and fear-potentiated startle. These NPY effects were not mimicked by intra-amygdala injections of the Y1R agonists Y-28 or Y-36, and co-infusion of the Y1R antagonist BIBO 3304 did not block the NPY effects. Furthermore, we tested Y1R-deficient mice in conditioned freezing and found no differences between wild type and mutant littermates. Finally, we injected NPY into the amygdala of Y1R-deficient mice. Y1R deficiency had no effect on the fear-reducing effects of intra-amygdala NPY.
Conclusions
These data show an important role of the transmitter NPY within the amygdala for the expression of conditioned fear. Y1R do not appear to be involved in the mediation of the observed intra-amygdala NPY effects suggesting that these effects are mediated via other NPY receptors.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Neuropeptide Y (NPY) is one of the most abundant peptides found in the mammalian brain (Tatemoto et al. 1982). Its effects are mediated by its G-protein-coupled receptors, among which, the Y1, Y2, Y4, and Y5 subtypes are located in the brain (Holmes et al. 2003). During the last decade, a crucial role of NPY in fear and anxiety has been shown in animal research (summarized in Kask et al. 2002; Holmes et al. 2003). Furthermore, decreased NPY levels were found in the plasma and the cerebrospinal fluid of patients suffering from depression or anxiety disorders (Rasmusson et al. 2000; Heilig et al. 2004).
The present study investigated the role of NPY within the amygdala in conditioned fear. Fear, in contrast to anxiety, is induced in animals and humans by cues which they can clearly identify, i.e., fear is specific; whereas, anxiety is more generalized (e.g., Davis and Shi 1999; Fendt 2006). A variety of studies demonstrated an important role of NPY in fear and anxiety: For example, NPY infusions into the cerebral ventricle reduce fear and anxiety behavior in the elevated plus maze, the Vogel’s punished drinking test, the social interaction test, fear-potentiated startle (FPS), and the light–dark box (Heilig et al. 1989, 1993; Pich et al. 1993; Broqua et al. 1995; Gutman et al. 2008). Pharmacological studies suggest an important role of NPY Y1 receptors (Y1R) in mediating the anxiety- and fear-reducing effects of NPY: Y1R agonists mimic the effects of exogenous NPY in different anxiety models (Sorensen et al. 2004; Bacchi et al. 2006), and the anxiolytic-like effects of intra-ventricular NPY injections are absent in Y1R deficient mice (Karlsson et al. 2008).
The amygdala is a crucial region of the brain circuitry mediating fear and anxiety (e.g., Fendt and Fanselow 1999; LeDoux 2000). NPY and its receptors are present in the amygdala (Kask et al. 2002; Holmes et al. 2003; Wolak et al. 2003; Kishi et al. 2005) leading to the hypothesis that the anxiolytic-like effects of NPY are mediated in part by the amygdala. Indeed, local injections of NPY into the amygdala, as well as NPY over-expression within the amygdala, induced anxiolysis in different animal models of anxiety (Heilig et al. 1993; Primeaux et al. 2005). Surprisingly, there is only one recently published study investigating the effects of intra-amygdala NPY injections in a conditioned fear model (Gutman et al. 2008) despite the extensive evidence that the amygdala is the central brain site for conditioned fear (e.g., Davis 1994; Wilensky et al. 2006; Kim and Jung 2006). Conditioned fear models play a central role in the investigation of the neural circuitry of emotions and are used to probe potential anxiolytic treatments in preclinical research (Garakani et al. 2006).
The aim of the present study was to investigate the effects of intra-amygdala injections of NPY on the expression of conditioned fear in mice and to explore whether the observed effects of intra-amygdala NPY injections are mediated by Y1R. As behavioral fear models, we used FPS and conditioned freezing.
Material and methods
Animals
Experimentally naive male mice with an age of 2–3 months were used: experiment 1: DBA/1J (n = 34), supplied by Janvier, Le Genest Saint Isle, France; experiments 2–4: C57Bl/6J (n = 200), supplied by Janvier, Le Genest Saint Isle, France; and experiment 5 and 6: Y1R deficient mice (n = 84). Y1R-deficient mice (B6-TgH(npyy1KO)) were purchased from the University of Lausanne, Switzerland, and bred in Novartis facilities (backcrossed for at least ten generations onto the C57BL/6JNpa background). Genotyping was performed using a Taqman assay (2 min, 50°C; 10 min 95°C, 40 cycles 15 s, 95°C; and 1 min, 60°C) on an ABI Prism 7900HT. Knockout allele-specific Neo primers used were TGGATTGCACGCAGGTTCT (sense), GTGCCCAGTCATAGCCGAAT (anti-sense), and CGGCCGCTTGGGTGGAGAGG (probe labeled 5′FAM and 3′BHQ1). Primers used for the glucocorticoid receptor gene as reference were CGGGACCACCTCCCAAA (sense), CCCCATAATGGCATCCCGAA (anti-sense), and CTTCATCGGAGCACACCAGGCAGA (probe labeled 5′YY and 3′BHQ1).
The animals were housed in groups of 2–4 in a humidity (55%) and temperature (22°C)-controlled room under a 12/12 h day-night cycle with lights on at 07:00 a.m. Water and food were available ad libitum. Experiments were in accordance with the Swiss law and international guidelines for the care and use of animals and covered by license #1988 and #2152 (Kantonales Veterinäramt Basel-Stadt, Switzerland).
Surgery (experiments 1–4, 6)
Mice were anesthetized with ketamine/xylazine (110 mg/kg, 10:1, i.p.) and placed into a stereotaxic frame. The skull was exposed, and stainless steel guide cannulae (diameter: 0.35 mm; length: 6 mm) were bilaterally implanted aiming to the amygdala. The following coordinates were used (Paxinos and Franklin 2001): 1.5 mm caudal from Bregma, ±3.5 mm lateral from Bregma, and −3.7 mm ventral from dura. The guide cannulae were fixed to the skull with dental cement and two to three anchoring screws (2 × 2.75 mm; Laubscher, Täuffelen, Switzerland). To prevent post-surgery pain, the analgesic Buprenorphine (0.01 mg/kg, i.p.) was given twice a day on the first 2 days following surgery. Behavioral tests started following full recovery (5–6 days).
Apparatus
Fear-potentiated startle (experiment 1)
For this experiment, a startle system with eight chambers (35 × 35 × 38 cm) was used (SR-LAB, San Diego Instruments, San Diego, CA, USA). Each chamber contained a small animal enclosure made of transparent Plexiglas cylinder with 4 cm inner diameter and 10 cm inner length. Movements of the animals were detected by motion-sensitive transducers mounted underneath the animal enclosure. For data acquisition, the output signal of the transducers was digitized (sampling rate 24 bit, 1 kHz) and stored on a computer. Stored responses were expressed in arbitrary units.
For fear conditioning, electric foot shocks and light stimuli were used. The light stimulus was presented to the animals with 10 W bulbs (ca. 700 lux) mounted to the back of the test chambers. Scrambled foot shocks were administered by a floor grid (seven parallel bars, 8 mm apart and 4 mm diameter) and had an intensity of 0.6 mA and a duration of 1 s. White background noise (50 dB sound pressure level (SPL)) and the acoustic noise stimuli (96 dB SPL) were generated by high-frequency loudspeakers mounted in the center of the ceiling of the test chambers.
Conditioned freezing (experiment 2–6)
To measure conditioned freezing, a computerized fear-conditioning system (TSE, Bad Homburg, Germany) was used. The apparatus consisted of four identical transparent Perspex boxes (46 × 46 × 32 cm) placed inside animal detection infrared sensors frame; each box was located in a sound-attenuating chamber provided with loudspeakers for the acoustic stimuli (background noise of 60 dB SPL and the tone stimuli for fear conditioning), light sources (continuous illumination of 10 lux), and a ventilation fan for air exchange. The floor of the boxes consisted of removable stainless steel grids (bars, 4 mm diameter; distance from rod center to rod center, 8.9 mm), which was connected to a shock unit and able to deliver foot shocks of defined duration and intensity. Delivery of all stimuli was controlled by a personal computer. Four additional boxes of the same size as those described above but made of black Perspex (including the floor) served to create a different test context.
Movements of the animals were detected by the infrared sensors. The time spent freezing (immobility) was automatically recorded (no infrared beam crosses for more than 1 s) during all phases of the experiments.
Drugs
Sterile saline (NaCl 0.9%) was used as a vehicle for all experiments. The following concentrations were used for the different compounds: NPY (Tocris via Lucerna Chem AG, Lucerne, Switzerland): 0.125, 0.25, and 0.5 μg/0.3 μl saline (i.e., 30, 60, and 120 pmol/brain side or 97.5, 185, and 390 μM); Y-28 (Des-AA11–18 [Cys7,21, d-Lys9 (Ac), d-His26, Pro34]-NPY, a specific Y1R agonist (Mullins et al. 2001)): 0.01, 0.1, or 0.4 μg/0.3 μl (i.e., 0.9, 9, and 36 pmol/brain side or 2.9, 29, and 116 μM); Y-36 ([d-Arg25, d-His26]-NPY, a specific Y1R agonist (Mullins et al. 2001)): 0.62 μg/0.3 μl (i.e., 120 pmol/brain side or 120 μM); and BIBO 3304 ((R)-2-Diphenylacetylamino-5-guanidino-pentanoic acid 4-ureidomethyl-benzylamide trifluoroacetate, a specific Y1R antagonist (Wieland et al. 1998)): 130 ng/0.3 μl (i.e., 60 pmol/brain side or 202 μM). Y-28 and Y-36 were synthesized by NeoMPS, Strasbourg, France, and the BIBO 3304 were synthesized by Novartis.
Experimental procedures
Experiment 1 (effects of intra-amygdala NPY on FPS expression)
On the first experimental day, the mice were tested for baseline acoustic startle response to ten startle stimuli and then matched into two treatment groups with similar mean baseline startle magnitudes. On the following 2 days, the animals were fear-conditioned. On each day, after a habituation period of 5 min, they received ten pairings of a 30-s light stimulus and a foot shock which was presented in the last second of the light stimulus. The mean intertrial interval was 140 s (range: 100–180 s).
On the fourth day, NPY or vehicle was injected into the amygdala during a short anesthesia with 1.5–3% isofluran applied through a mask. Therefore, injectors with a diameter of 0.15 mm (connected to Hamilton syringes by tubes) were introduced into the guide cannulae, and a total volume of 0.3 μl solution/side was injected with a speed of 0.1 μl/min, controlled by a microinfusion pump (CMA100, CMA, Stockholm, Sweden). The injector was removed after additional 60 s, and the mice were returned in their home cage for 10 min. Then, the animals were put into the startle devices and after an acclimatization period of 5 min, 12 startle stimuli of 96 dB SPL were administered, half of them alone, the other half of them were preceded by the conditioned light stimulus (in a pseudorandomized order). Inter-stimulus interval was 120 s, and no shocks were administered on this retention test day.
Experiment 2 (effects of intra-amygdala NPY injections on conditioned freezing)
Fear conditioning was performed on the first experimental day using the transparent Perspex boxes. The animals were individually placed into the boxes. Sixty seconds later, the first of six pairings of a tone stimulus (8 kHz, 80 dB, 30 s, pulsed with an interval of 100 ms) and a scrambled foot shock (0.4 mA, during the last second of the tone stimulus) were presented. The inter trial intervals were 60 s. Thirty seconds after the last pairing, the mice were returned to the home cage. Then, the boxes were cleaned with 70% ethanol and dried.
On the next day, NPY or vehicle was injected into the amygdala during a short anesthesia with 1.5–3% isofluran applied through a mask. Therefore, injectors with a diameter of 0.15 mm (connected to Hamilton syringes by tubes) were introduced into the guide cannulae, and a total volume of 0.3 μl solution/side were injected with a speed of 0.1 μl/min, controlled by a microinfusion pump. The injector was removed after additional 60 s, and the mice were returned to their home cage for 10 min. Then, the animals were placed in the black Perspex boxes, and after a habituation period of 2 min, five tone stimuli (without foot shocks) were presented with an interstimulus time of 1 min (retention test).
Experiment 3 (effects of intra-amygdala injections of Y-28 on conditioned freezing)
Except for the following details, the same procedure as in experiment 2 was used. For fear conditioning, the foot shocks had a duration of 2 s and an intensity of 0.6 mA, and the frequency of the tone stimulus was 10 kHz. Furthermore, the injections before the retention test were performed without anesthesia.
In this experiment, fear conditioning was again without any injections. On the next day, the mice received injections of 0.3 μl/side vehicle, 0.01, 0.1, or 0.4 μg/side Y-28 (Y1R agonist), or 0.5 μg/side NPY (as a positive control) into the amygdala. Fifteen minutes later, they were tested for expression of conditioned freezing.
Experiment 4 (effects of intra-amygdala injections of Y-36 and BIB0 3304 on conditioned freezing)
In this experiment, 0.3 μl/side vehicle, 0.62 μg/side Y-36 (Y1R agonist), 0.5 μg/side NPY (as a positive control), 130 ng/side BIBO 3304 (Y1R antagonist), or a combination of NPY and BIBO 3304 was injected into the amygdala. In contrast to the previous experiment, the retention test now consisted of 15 tone stimulus presentations to evaluate potential effects of the injections on extinction of conditioned fear. Furthermore, a second retention test with five tone stimulus presentations was carried out on the third experimental day (without any treatment) to probe fear extinction memory.
Experiment 5 (effects of Y1R deficiency on conditioned freezing)
In this experiment, the animals had no cannulae implanted and were not treated. Fear conditioning was carried out as in experiments 3 and 4. On day 2, the animals were first exposed to the conditioning context for 5 min. Three hours later, the conditioned stimulus (CS) was presented ten times in the black Perspex boxes. This CS exposure was identically repeated on the next 3 days (extinction).
Experiment 6 (effects of intra-amygdala NPY injections in Y1R deficient mice on conditioned freezing)
Again, the behavioral procedure of experiment 3 was used. Animals received intra-amygdala injections of 0.3 μl/side vehicle or 0.5 μg/side NPY.
Histology (experiments 1–4, 6)
Immediately after the final behavior test, all mice with intra-amygdala cannulae were euthanized, and the brains were removed for histological analysis in order to verify the injection sites within the amygdala. The brains were immersion-fixed with 4% formaldehyde and stored in fixative until further processing. Prior to cutting, the brains were transferred to phosphate buffered 30% sucrose for at least 12 h. Frontal sections (100 μm) were cut on a freezing microtome and stained with cresyl violet. The injection sites were localized, and the extent of tissue lesions due to cannulation was examined under a light microscope. The injection sites were drawn onto plates taken from a mouse brain atlas (Paxinos and Franklin 2001).
Statistical analysis
All reported statistical tests were performed using the program SYSTAT (Statistical Package for the Social Sciences Inc., version 11). An analysis of variance (ANOVA) was used with the between-subject factor treatment (experiments 1–4, 6) or genotype (experiment 5 and 6) and the within-subject factors trial type (experiment 1) or trial number (experiments 2–6). In case of significance (p < 0.05), post hoc Dunnett’s or Tukey tests were carried out.
Results
Histology
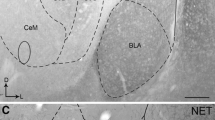
All injection sites from mice that were included in the statistical analysis were bilaterally localized within the amygdala, mainly in the central, lateral, and basolateral subnuclei (Fig. 1). Thirty-six animals were excluded from final analysis due to amygdaloid lesions caused by cannulation or due to misplaced injection sites.

Frontal sections of the amygdala (adapted from Paxinos and Franklin 2001) depicting the injection sites of the different experiments into the amygdala. The photomicropraphs show representative examples of two different animals. BLA basolateral amygdala, CA central amygdala, LA lateral amygdala
Experiment 1: effects of intra-amygdala NPY on FPS expression
The aim of this experiment was to test whether conditioned fear, measured by FPS expression, is affected by intra-amygdala injections of NPY. Twenty-seven mice with bilateral injection sites within the amygdala (vehicle: N = 12, NPY: N = 15) were tested. Figure 2a depicts the mean startle amplitudes (mean + SEM) after startle stimuli alone (tone alone), after startle stimuli preceded by the conditioned light stimuli (light-tone), as well as their difference (FPS). Statistical analysis revealed the presence of conditioned fear indicated by significant effects of the trial type (tone alone vs light-tone: F(1,25) = 8.43, P = 0.008). Injections of NPY into the amygdala attenuated FPS expression shown by a significant interaction between trial type and treatment (F(1,25) = 5.35, P = 0.03); whereas, baseline startle magnitude was not generally affected by NPY injections (factor treatment: F(1,25) = 1.63, ns).

Effects of intra-amygdala NPY injections on the expression of fear-potentiated startle (a) and conditioned freezing (b). Data are presented as means + SEM (standard errors of the mean). *P < 0.05 post hoc Dunnett’s test (comparison with 0 μg) after a significant main effects (ANOVA)
Experiment 2: effects of intra-amygdala NPY on expression of conditioned freezing
With this experiment, we wanted to explore (1) whether the anxiolytic-like effects observed in experiment 1 can also be seen in other models of conditioned fear, and (2) whether this effect is dose-dependent. On the first experimental day, fear conditioning was carried out without any treatment. In contrast to the FPS paradigm, it is possible in freezing experiments to measure fear behavior during the conditioning phase which allows one to check for group differences before the critical treatment. In the present experiment, the four different groups of animals were not different in baseline immobility (ANOVA: F < 1; data not shown), i.e., in the immobility shown before presentation of the first CS-US pairing. During the fear-conditioning training, there was an increase of freezing behavior (ANOVA, factor trial number: F(5,230) = 32.45, P < 0.001; data not shown) which was not different in the different groups (factor group: F(3,46) = 1.08, ns; interaction trial number × group: F(15,230) = 1.04, ns) indicating successful fear conditioning in all groups.
On day 2, the animals were injected (vehicle: N = 12, 0.125 μg: N = 11, 0.25 μg: N = 13, 0.5 μg: N = 14) and tested for expression of conditioned freezing. Figure 2b shows the percent time spent freezing (mean + SEM) of the different groups of animals during the presentation of the conditioned tone stimulus. There was a significant effect of the factor treatment (F(3,46) = 2.92, P = 0.04) indicating that NPY injections into the amygdala reduced expression of conditioned fear. Furthermore, an effect of trial number was detected (F(4,184) = 10.49, P < 0.001) reflecting within-session extinction of conditioned fear. This extinction was not affected by the NPY injections (interaction treatment × trial number: F < 1). A post hoc Dunnett test showed a significant reduction of conditioned freezing after intra-amygdala injections of 0.5 μg NPY (P = 0.02) and a trend to an effect after 0.25 μg NPY (P = 0.097) vs vehicle. There were no statistical effects of NPY on either baseline immobility or motor activity (measure by distance traveled) in the habituation period before CS presentation (F < 1; data not shown).
Experiment 3: effects of intra-amygdala injections of Y-28 on conditioned freezing
The aim of this experiment was to test whether intra-amygdala injections of a Y1R agonist mimic the effects of intra-amygdala NPY injections. Therefore, we injected the specific Y1R agonist Y-28 into the amygdala and tested the effects of these injections on the expression of conditioned freezing (vehicle: N = 11, 0.01 μg: N = 10, 0.1 μg: N = 15, 0.4 μg: N = 13). As in experiment 2, the behavior (baseline immobility, freezing behavior) of the different groups was not different in the conditioning session (F < 1.24, ns; data not shown). Figure 3a depicts the percent time spent freezing (mean + SEM) during the presentation of the conditioned tone stimulus after injections of different Y-28 doses (retention test). Statistical analysis revealed no effects of intra-amygdala Y-28 on conditioned freezing (ANOVA, factor treatment: F(3,45) = 1.73, ns). However, there was a trend to more rapid extinction after intra-amygdala Y-28 injections (interaction treatment × trial number: F(12,180) = 1.64, P = 0.09; factor trial number: F(4,180) = 15.67, P < 0.001; extinction time course not shown).

Effects of intra-amygdala injections of 0.5 μg/side NPY (a, b), of the Y1R agonists Y-28 (0.01–0.4 μg/side; a) and Y-36 (0.62 μg/side; b), and of the Y1R antagonist BIBO 3304 (130 ng/side; b) on the expression of conditioned freezing (mean of the first five CS presentation). Data are presented as means + SEM. **P < 0.01 post hoc Dunnett’s test (comparison with 0 μg) after a significant main effects (ANOVA)
As in experiment 2, intra-amygdala NPY injections (N = 14) had anxiolytic-like effects on conditioned freezing (factor treatment: F(1,23) = 12.68, P = 0.002, interaction treatment × trial number: F(4,92) = 1.58, ns; factor trial number: F(4,92) = 6.75, P < 0.001).
Experiment 4: effects of intra-amygdala injections of Y-36 and BIBO 3304 on conditioned freezing
The aim of our fourth experiment was to test (1) whether another Y1R agonist, Y-36, mimics the effects of intra-amygdala NPY; (2) whether a Y1R antagonist, BIBO 3304, has opposite effects to those of intra-amygdala NPY, i.e., anxiogenic-like effects; (3) whether the Y1R antagonist, BIBO 3304, attenuates or even blocks the effects of intra-amygdala NPY injections; and (4) whether any of these treatments affect within- or between-session extinction of conditioned fear. As in the previous experiments, the behavior (baseline immobility, freezing behavior) of the different treatment groups was not different in the conditioning session (F < 1; Fig. 4, day 1). After intra-amygdala injections of vehicle (N = 12), Y-36 (N = 14), NPY (N = 12), BIBO 3304 (N = 13), and NPY + BIBO 3304 (N = 12), expression of conditioned fear to the first five CS presentations (see Figs. 3b and 4, day 2) was reduced in the two groups which received either NPY alone or NPY in combination with BIBO 3304 (separated ANOVAs vs vehicle injection: NPY: factor treatment: F(1,22) = 8.13, P = 0.009; interaction treatment × trial number: F < 1; factor trial number: F(4,88) = 3.38, P = 0.01; NPY + BIBO 3304: factor treatment: F(1,22) = 7.73, P = 0.01; interaction treatment × trial number: F < 1; factor trial number: F(4,88) = 5.93, P < 0.001). Neither intra-amygdala injection of Y-36 nor of BIBO 3304 affected the expression of conditioned freezing (Y-36: factor treatment: F < 1; interaction treatment × trial number: F < 1; factor trial number: F(4,96) = 4.63, P = 0.002; BIBO 3304: factor treatment: F < 1; interaction treatment × trial number: F < 1; factor trial number: F(4,92) = 6.01, P < 0.001).

Time course of experiment 4 (intra-amygdala injections of 0.5 μg/side NPY, 0.62 μg/side Y-36, and 130 ng/side BIBO 3304). Data are presented as means + SEM. Day 1, fear conditioning with six pairings of the tone CS with the foot shock US. Day 2, retention test with 15 presentations of the CS. Day 3, extinction test with five CS presentations
If not only the first five CS presentation but all 15 CS presentations were analyzed (Fig. 4, day 2), the statistical results were qualitatively identical (data not shown) with one exception: Intra-amygdala BIBO 3304 injections had again no general effect on fear expression (factor treatment: F < 1) but within-session extinction was facilitated (interaction treatment × trial number: F(14,322) = 1.91, P = 0.03; factor trial number: F(14,322) = 7.44, P < 0.001). However, freezing behavior on the next day (second retention test without treatment; Fig. 4, day 3) was neither changed for the group that received BIBO on the first retention test (factor treatment: F < 1; interaction trial number × treatment: F(4,92) = 1.06, ns) nor for the other treatment groups (factor treatment: F < 1; interaction trial number × treatment: F < 1.55, ns), indicating that between-session extinction was not affected. In both retention tests, baseline immobility before CS presentation was not affected by the treatments (F < 1.74, ns). Therefore, conditioned freezing was not affected by intra-amygdala Y1R agonist or antagonists, and the effects of intra-amygdala NPY were not blocked by co-administration of a Y1R antagonist.
Experiment 5: effects of Y1R deficiency on conditioned freezing
To further investigate this apparent lack of Y1R involvement in the intra-amygdala anxiolytic effect of NPY, the effects of Y1R deficiency were tested in our next experiment. Therefore, heterozygote (N = 12) and homozygote (N = 11) Y1R deficient mice as well as their wild type littermates (N = 12) were tested for acquisition, expression, and extinction of conditioned fear.
There was no effect of genotype on baseline inactivity in the phase before the first presentation of the tone CS (ANOVA: F(2,32) = 1.21, ns). In Fig. 5a, freezing behavior during fear conditioning is depicted. The response to the first CS presentation did not differ between the genotypes (F(2,32) = 1.41, ns). There was a significant increase in the freezing response during the CS presentations (F(5,160) = 15.84, P < 0.001) indicating successful fear conditioning. In addition, there was a main effect of genotype (F(2,32) = 3.28, P = 0.05; no significant effects detected in post hoc pairwise comparisons), i.e., generally higher freezing levels in Y1R deficient animals during fear conditioning but no interaction genotype × trial number (F < 1).

Conditioned freezing in Y1R deficient mice, heterozygous, and wild type littermates. a Fear conditioning with six pairings of the tone CS with the foot shock US. b First retention test on contextual fear and on cue-induced fear (mean of the first five CS presentation). c Time course of first retention test (all ten CS presentations). d Mean freezing response of day 2 to 5 (between-session extinction). Data are presented as means + SEM
On the following day, expression of conditioned fear was tested. There was no effect of genotype on contextual fear (F < 1; Fig. 5b), on freezing during the first five CS presentation (F < 1; Fig. 5b), or on freezing during all ten CS presentation (F(2,32) = 1.08, ns; Fig. 5c). After ten CS presentations, within-session extinction was observed (F(9,288) = 5.52, P < 0.001; Fig. 5c), and there was an interaction between genotype and trial number (F(18,288) = 1.68, P = 0.04) indicating less within-session extinction in the Y1R deficient mice. On the following test days (Fig. 5d), between-session extinction was observed (F(3,96) = 78.11, P < 0.001). There was no main effect of genotype (F < 1) but a significant interaction of genotype × test day (F(6,96) = 2.29, P = 0.04) indicating stronger between-session extinction in Y1R-deficient mice.
Experiment 6: effects of intra-amygdala NPY injections in Y1R deficient mice on conditioned freezing
The aim of the last experiment was to investigate whether the anxiolytic-like effects of intra-amygdala NPY injections can also be observed in Y1R deficient mice.
As in experiment 5, freezing behavior during fear conditioning was not affected by genotype (data not shown; factor gene: F < 1; factor trial number: F(5,200) = 35.97, P < 0.001; interaction gene × trial number: F < 1).
In Fig. 6, the freezing response during the presentation of five CS after injection of either vehicle (wild type: N = 11, deficient: N = 12) or 0.5 μg NPY (wild type: N = 10; deficient: N = 11) is depicted. Again, the genotype of the animal did not affect expression of conditioned freezing (F < 1). As in the previous experiments, intra-amygdala NPY injections significantly decreased conditioned freezing (F(1,40) = 9.70, P = 0.003). However, this NPY effect was quite similar in the different genotypes (interaction gene × treatment: F < 1). Furthermore, within-session extinction (factor trial number: F(4,140) = 14.96, P < 0.001) was neither affected by genotype (F(4,160) = 2.13, P = 0.08) nor by NPY injections (F(4,160) = 1.40, ns; interaction gene × treatment × trial number: F(4,160) = 2.08, P = 0.09).

Fear expression after intra-amygdala NPY injections in Y1R deficient mice and wild type littermates. Data are presented as means + SEM. **P < 0.01 post hoc Tukey test after significant main effects (ANOVA)
Discussion
The aims of our study were to test whether intra-amygdala NPY injections have fear-reducing effects in animal models of conditioned fear and whether such effects are mediated by Y1R. In summary, our experiments showed that intra-amygdala NPY significantly reduces fear expression in FPS and conditioned freezing. The latter model was further used to clarify whether the fear-reducing effects of intra-amygdala NPY are mediated by Y1R. First, we tried to mimic the NPY effects by intra-amygdala injections of specific Y1R agonists. Two different agonists, Y-28 and Y-36, did not affect fear expression. Furthermore, a specific Y1R antagonist, BIBO 3304, was neither able to block the NPY effects after co-administration nor had any effects when injected alone. Finally, we evaluated Y1R-deficient mice in conditioned freezing. Fear expression was not affected in Y1R-deficient mice, and the fear-reducing effects of intra-amygdala NPY were not affected in Y1R-deficient mice.
There are numerous studies supporting an important role of NPY in human anxiety disorders (Rasmusson et al. 2000; Heilig et al. 2004), as well as in traditional animal models of anxiety (e.g., Heilig et al. 1993; Sajdyk et al. 1999; Krysiak et al. 2000; Sorensen et al. 2004; Karlsson et al. 2005). However, there are only few studies in animal models of conditioned fear which are believed to be more translational than animal models of anxiety (e.g., Grillon and Baas 2003; Garakani et al. 2006). A very recent study showed that NPY injections into the basolateral amygdala reduce the expression of FPS in rats (Gutman et al. 2008).
In the present study, NPY was injected into the amygdala before the retention test for cue-induced conditioned fear. In two different models, conditioned freezing and FPS, we observed robust fear-reducing effects of intra-amygdala NPY injections. These results provide the first demonstration of the efficacy of intra-amygdala NPY injections in mice and confirm the recently published data in rats showing fear-reducing effects after such injections of NPY in FPS (Gutman et al. 2008). Similar intra-amygdala NPY effects were observed in animal models of anxiety (Heilig 1995; Sajdyk et al. 1999). In all of these studies, rats were used as experimental subjects, and the injections were made into the basolateral region of the amygdala. However, in mice (present study), due to their much smaller size, it is not really possible to precisely hit a subnucleus of the amygdala using normal injection cannulae. Most of our injections sites were located in the basolateral part of the amygdala (which also has the highest Y1R density within the amygdala; Wolak et al. 2003; Kishi et al. 2005). However, since the injected volume (0.3 μl) probably floods the whole mouse amygdala complex, our experiments do not allow to conclude which amygdaloid subnucleus mediates the observed NPY effects.
Beside from using mice, our study has another critical difference to literature studies investigating intra-ventricular or intra-amygdala NPY effects on anxiety: We used models of conditioned fear. These models were chosen since the amygdala plays a very crucial role in conditioned fear (e.g., Davis 1992; LeDoux 2000; Antoniadis and McDonald 2001), and pathological processing of conditioned fear by the amygdala is believed to be involved in the etiology of human anxiety disorders (e.g., Etkin and Wager 2007; Mineka and Oehlberg 2008). As in the anxiety models in rats (Heilig 1995; Sajdyk et al. 1999), we observed robust fear-reducing effects of intra-amygdala NPY injections in our mouse experiments. It is important to mention that other potentially interfering behavioral effects such as changes in motor activity and baseline immobility or startle magnitude were not observed (see results).
After demonstrating the fear-reducing effects of intra-amygdala NPY in these two models of conditioned fear, our next step was to test whether these effects can be mimicked by Y1R agonists. We used two different specific Y1R agonists, Y-28 and Y-36. These agonists both have high affinities and potencies for Y1R (Mullins et al. 2001; data confirmed by in-house tests), and we used concentrations which are clearly over the EC50s for Y1R. In fact, the highest concentrations used in the present study were more than 100-folds above the EC50 of these agonists, and higher doses would be unspecific due to some potency of the Y-28 and Y-36 on NPY Y2 and Y4 receptors (Mullins et al. 2001). Usually, concentrations of ca. five- to tenfold EC50 are enough to get behavioral effects with intracranial injections (e.g., Fendt et al. 2008; Siegl et al. 2008). However, both agonists were not able to mimic the fear-reducing effects of intra-amygdala NPY injections. In addition, co-administration of BIBO 3304, a specific Y1R antagonist (Wieland et al. 1998), was not able to block the fear-reducing effects of NPY injections into the amygdala, and this, despite the evidence from other studies that the dose of BIBO 3304 used, had behavioral effects per se and blocked NPY effects (Sajdyk et al. 1999; Wieronska et al. 2004, 2008; Smialowska et al. 2007). Our data are in line with those of Gutman et al. (2008) which found no effects of intra-amygdala BIBO 3304 injections on expression of FPS in rats. However, it cannot totally be excluded that intra-amygdala injections of Y1R agonists or antagonists could be effective if other conditioning protocols are used (e.g., less pairings during conditioning).
Finally, we conducted a further and perhaps optimal approach to probe the role of Y1R in the NPY effects we observed: NPY was injected into the amygdala of Y1R-deficient mice. Such an experiment is only conclusive if Y1R-deficient mice have no particular phenotype in conditioned fear. In literature, there are quite different phenotypes of Y1R-deficient mice described: In one study, they express an anxiolytic-like phenotype in the elevated plus maze but an anxiogenic-like phenotype in the light–dark box (Karl et al. 2006). In another study (Karlsson et al. 2008), they showed no phenotype in these anxiety models, as well as in Pavlovian fear conditioning. In our experiment, Y1R-deficient mice expressed slightly increased freezing levels during fear conditioning but showed no phenotype in the expression of conditioned fear. Furthermore, the fear-reducing effects of intra-amygdala NPY injections were still present in Y1R-deficient mice. It should be noted that Y1R-deficient mice have normal mRNA levels of NPY Y2 and Y5 receptors, as well as normal NPY plasma concentrations (Pedrazzini et al. 1998) which argue against possible compensatory changes in Y1R-deficient mice.
This result, together with our other experiments, clearly indicates that Y1R within the amygdala is not involved in the expression of conditioned fear. This apparently stands in contrast to other studies showing that amygdaloid Y1R are crucial for mediating the anxiolytic-like effects of intracranial or intra-amygdaloid NPY (Heilig et al. 1993; Heilig 1995; Sajdyk et al. 1999). However, in all the latter studies, anxiety or conflict models were used (social interaction test, Geller-Seifter test), and the idea that the neural circuitries underlying anxiety and conditioned fear are to some extend different (e.g., Davis 1998; McNaughton and Corr 2004; Rosen 2004) could explain these apparently contradictory findings. In this respect, it should be noted that Y1R-deficient mice expressed increased levels of fear during fear conditioning (here, both the context and the cue is present). Furthermore, freezing to the conditioning context was non-significantly increased. Based on the hypothesis that contextual fear is more related to anxiety (Grillon et al. 2006), these observations of the present study well fit to the published role of Y1R in anxiety models.
In our experiments, we also observed a potential role of Y1R in fear extinction. It should be noted that our experimental protocols were not optimized to test for extinction effects, and we also used different extinction protocols in the different experiments. Nevertheless, in each experiment, significant extinction was observed in the control groups. In general, our data do provide support for the idea that NPY is involved in extinction (Gutman et al. 2008). Gutman et al. demonstrated a facilitation of fear extinction after NPY injections into the ventricle, but not after injections into the amygdala. In line with these data, we observed no effect of intra-amygdala NPY injections on extinction. However, we found slight effects on fear extinction after intra-amygdala injections of the Y1R antagonist BIBO 3304 and in Y1R-deficient mice (experiment 4). Surprisingly, the effects of intra-amygdala injections of the Y1R antagonist were not in line with the changes observed in Y1R-deficient mice. In addition, Gutman et al. reported a blockade of between-session extinction after BIBO 3304 injections into the basolateral (but not into the medial) amygdala. This was clearly not the case in our experiments (experiment 4): We even observed a weak facilitation of between-session extinction in Y1R-deficient mice. In summary, the here-observed effects on extinction are very weak, incongruous, and not completely in line with literature data. Future studies have to clarify the exact role of Y1R in the different brain areas in fear extinction.
In summary, the present experiments show strong and robust effects of intra-amygdala injections on conditioned fear in mice. However, our different experimental approaches did not support an involvement of Y1R in these effects. Therefore, we conclude that the intra-amygdala NPY effects on conditioned fear are mediated by other NPY receptor subtypes. A good candidate therefore is the NPY Y5 receptor (Y5R). Several studies already demonstrated an important role of Y5R (Sajdyk et al. 2002; Sorensen et al. 2004). Furthermore, there are indications that Y1R and Y5R co-expression results in heterodimerization and affect agonist and antagonist responses as well as receptor internalization which may account for a very complex NPY pharmacology (Gehlert et al. 2007). Clearly, more research is necessary to unravel the mechanisms underlying NPY-mediated anxiolysis.
Abbreviations
- ANOVA:
-
Analysis of variance
- CS:
-
Conditioned stimulus
- FPS:
-
Fear-potentiated startle
- NPY:
-
Neuropeptide Y
- Y1R:
-
NPY Y1 receptor
- SPL:
-
Sound pressure level
References
Antoniadis EA, McDonald RJ (2001) Amygdala, hippocampus, and unconditioned fear. Exp Brain Res 138:200–209
Bacchi F, Mathe AA, Jimenez P, Stasi L, Arban R, Gerrard P, Caberlotto L (2006) Anxiolytic-like effect of the selective Neuropeptide Y Y2 receptor antagonist BIIE0246 in the elevated plus-maze. Peptides 27:3202–3207
Broqua P, Wettstein JG, Rocher MN, Gauthier-Martin B, Junien JL (1995) Behavioral effects of neuropeptide Y receptor agonists in the elevated plus-maze and fear-potentiated startle procedure. Behav Pharmacol 6:215–222
Davis M (1992) The role of the amygdala in fear-potentiated startle: implications for animal models of anxiety. Trends Pharmacol Sci 13:35–41
Davis M (1994) The role of the amygdala in emotional learning. Int Rev Neurobiol 36:225–266
Davis M (1998) Are different parts of the extended amygdala involved in fear versus anxiety? Biol Psychiatry 44:1239–1247
Davis M, Shi C (1999) The extended amygdala: are the central nucleus of the amygdala and the bed nucleus of the stria terminalis differentially involved in fear versus anxiety. Ann NY Acad Sci 877:281–291
Etkin A, Wager TD (2007) Functional neuroimaging of anxiety: a meta-analysis of emotional processing in PTSD, social anxiety disorder, and specific phobia. Am J Psychiatry 164:1476–1488
Fendt M (2006) Animal models of fear and anxiety. In: Koch M (ed) Animal models of neuropsychiatric diseases. Imperial College Press, London, pp 293–336
Fendt M, Fanselow MS (1999) The neuroanatomical and neurochemical basis of conditioned fear. Neurosci Biobehav Rev 23:743–760
Fendt M, Schmid S, Thakker DR, Jacobson LH, Yamamoto R, Mitsukawa K, Maier R, Natt F, Hüsken D, Kelly PH, McAllister KH, Hoyer D, van der Putten H, Cryan JF, Flor PJ (2008) mGluR7 facilitates extinction of aversive memories and controls amygdala plasticity. Mol Psychiatry 13:970–979
Garakani A, Mathew SJ, Charney DS (2006) Neurobiology of anxiety disorders and implications for treatment. Mt Sinai J Med 73:941–949
Gehlert DR, Schober DA, Morin M, Berglund MM (2007) Co-expression of neuropeptide YY1 and Y5 receptors results in heterodimerization and altered functional properties. Biochem Pharmacol 74:1652–1664
Grillon C, Baas JM (2003) A review of the modulation of the startle reflex by affective states and its application in psychiatry. Clin Neurophysiol 114:1557–1579
Grillon C, Baas JM, Pine DS, Lissek S, Lawley M, Ellis V, Levine J (2006) The benzodiazepine alprazolam dissociates contextual fear from cued fear in humans as assessed by fear-potentiated startle. Biol Psychiatry 60:760–766
Gutman AR, Yang Y, Ressler KJ, Davis M (2008) The role of neuropeptide Y in the expression and extinction of fear-potentiated startle. J Neurosci 28:12682–12690
Heilig M (1995) Antisense inhibition of neuropeptide Y (NPY)-Y1 receptor expression blocks the anxiolytic-like action of NPY in amygdala and paradoxically increases feeding. Regul Pept 59:201–205
Heilig M, Söderpalm B, Engel JA, Widerlöv E (1989) Centrally administered neuropeptide Y (NPY) produces anxiolytic-like effects in animal anxiety models. Psychopharmacology 98:524–529
Heilig M, McLeod S, Brot M, Heinrichs SC, Menzaghi F, Koob GF, Britton KT (1993) Anxiolytic-like action of neuropeptide Y: mediation by Y1 receptors in amygdala, and dissociation from food intake effects. Neuropsychopharmacology 8:357–363
Heilig M, Zachrisson O, Thorsell A, Ehnvall A, Mottaggui-Tabar S, Sjogren M, Asberg M, Ekman R, Wahlestedt C, Agren H (2004) Decreased cerebrospinal fluid neuropeptide Y (NPY) in patients with treatment refractory unipolar major depression: preliminary evidence for association with preproNPY gene polymorphism. J Psychiatr Res 38:113–121
Holmes A, Heilig M, Rupniak NMJ, Steckler T, Griebel G (2003) Neuropeptide systems as novel therapeutic targets for depression and anxiety disorders. Trends Pharmacol Sci 24:580–588
Karl T, Burne THJ, Herzog H (2006) Effect of Y1 receptor deficiency on motor activity, exploration, and anxiety. Behav Brain Res 167:87–93
Karlsson RM, Holmes A, Heilig M, Crawley JN (2005) Anxiolytic-like actions of centrally-administered neuropeptide Y, but not galanin, in C57BL/6J mice. Pharmacol Biochem Behav 80:427–436
Karlsson RM, Choe J, Cameron H, Thorsell A, Crawley J, Holmes A, Heilig M (2008) The neuropeptide Y Y1 receptor subtype is necessary for the anxiolytic-like effects of neuropeptide Y, but not the antidepressant-like effects of fluoxetine, in mice. Psychopharmacology 195:547–557
Kask A, Harro J, von Horsten S, Redrobe JP, Dumont Y, Quirion R (2002) The neurocircuitry and receptor subtypes mediating anxiolytic-like effects of neuropeptide Y. Neurosci Biobehav Rev 26:259–283
Kim JJ, Jung MW (2006) Neural circuits and mechanisms involved in Pavlovian fear conditioning: a critical review. Neurosci Biobehav Rev 30:188–202
Kishi T, Aschkenasi CJ, Choi BJ, Lopez ME, Lee CE, Liu HY, Hollenberg AN, Friedman JM, Elmquist JK (2005) Neuropeptide Y Y1 receptor mRNA in rodent brain: distribution and colocalization with melanocortin-4 receptor. J Comp Neurol 482:217–243
Krysiak R, Obuchowicz E, Herman ZS (2000) Conditioned fear-induced changes in neuropeptide Y-like immunoreactivity in rats: the effect of diazepam and buspirone. Neuropeptides 34:148–157
LeDoux JE (2000) Emotion circuits in the brain. Annu Rev Neurosci 23:155–184
McNaughton N, Corr PJ (2004) A two-dimensional neuropsychology of defense: fear/anxiety and defensive distance. Neurosci Biobehav Rev 28:285–305
Mineka S, Oehlberg K (2008) The relevance of recent developments in classical conditioning to understanding the etiology and maintenance of anxiety disorders. Acta Psychol 127:567–580
Mullins D, Kirby D, Hwa J, Guzzi M, Rivier J, Parker E (2001) Identification of potent and selective neuropeptide Y Y(1) receptor agonists with orexigenic activity in vivo. Mol Pharmacol 60:534–540
Paxinos G, Franklin KBJ (2001) The mouse brain in stereotaxic coordinates, 2nd edn. Academic, San Diego
Pedrazzini T, Seydoux J, Künstner P, Aubert JF, Grouzmann E, Beermann F, Brunner HR (1998) Cardiovascular response, feeding behavior and locomotor activity in mice lacking the NPY Y1 receptor. Nat Med 4:722–726
Pich EM, Agnati LF, Zini I, Marrama P, Carani C (1993) Neuropeptide Y produces anxiolytic effects in spontaneously hypertensive rats. Peptides 14:909–912
Primeaux SD, Wilson SP, Cusick MC, York DA, Wilson MA (2005) Effects of altered amygdalar neuropeptide Y expression on anxiety-related behaviors. Neuropsychopharmacology 30:1589–1597
Rasmusson AM, Hauger RL, Morgan CA III, Bremner JD, Charney DS, Southwick SM (2000) Low baseline and yohimbine-stimulated plasma neuropeptide Y (NPY) levels in combat-related PTSD. Biol Psychiatry 47:526–539
Rosen JB (2004) The neurobiology of conditioned and unconditioned fear: a neurobehavioral system analysis of the amygdala. Behav Cogn Neurosci Rev 3:23–41
Sajdyk TJ, Vandergriff MG, Gehlert DR (1999) Amygdalar neuropeptide Y Y1 receptors mediate the anxiolytic-like actions of neuropeptide Y in the social interaction test. Eur J Pharmacol 368:143–147
Sajdyk TJ, Schober DA, Gehlert DR (2002) Neuropeptide Y receptor subtypes in the basolateral nucleus of the amygdala modulate anxiogenic responses in rats. Neuropharmacology 43:1165–1172
Siegl S, Flor PJ, Fendt M (2008) Amygdaloid metabotropic glutamate receptor subtype 7 is involved in the acquisition of conditioned fear. NeuroReport 19:1147–1150
Smialowska M, Wieronska JM, Domin H, Zieba B (2007) The effect of intrahippocampal injection of group II and III metobotropic glutamate receptor agonists on anxiety; the role of neuropeptide Y. Neuropsychopharmacology 32:1242–1250
Sorensen G, Lindberg C, Wörtwein G, Bolwig TG, Woldbye DPD (2004) Differential roles for neuropeptide Y Y1 and Y5 receptors in anxiety and sedation. J Neurosci Res 77:723–729
Tatemoto K, Carlquist M, Mutt V (1982) Neuropeptide Y—a novel brain peptide with structural similarities to peptide YY and pancreatic polypeptide. Nature 296:659–660
Wieland HA, Engel W, Eberlein W, Rudolf K, Doods HN (1998) Subtype selectivity of the novel nonpeptide neuropeptide Y Y1 receptor antagonist BIBO 3304 and its effect on feeding in rodents. Br J Pharmacol 125:549–555
Wieronska JM, Stachowicz K, Klodzinska A, Smialowska M, Pilc A (2004) Intraamygdaloid administration of BIBO 3304 increases water intake and extends anxiolytic effects. Pol J Pharmacol 56:867–870
Wieronska JM, Smialowska M, Branski P, Gasparini F, Klodzinska A, Szewczyk B, Palucha A, Chojnacka-Wojcik E, Pilc A (2008) In the amygdala anxiolytic action of mGlu5 receptors antagonist MPEP involves neuropeptide Y but not GABAA signaling. Neuropsychopharmacology 29:514–521
Wilensky AE, Schafe GE, Kristensen MP, LeDoux J (2006) Rethinking the fear circuit: the central nucleus of the amygdala is required for the acquisition, consolidation, and expression of pavlovian fear conditioning. J Neurosci 26:12387–12396
Wolak ML, deJoseph MR, Cator AD, Mokashi AS, Brownfield MS, Urban JH (2003) Comparative distribution of neuropeptide YY1 and Y5 receptors in the rat brain by using immunohistochemistry. J Comp Neurol 464:285–311
Acknowledgements
The authors thank Agnes Jeker, Charlotte Huber, and Chantal Portet for their excellent technical help and Dr. Christopher Pryce for his helpful comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Fendt, M., Bürki, H., Imobersteg, S. et al. Fear-reducing effects of intra-amygdala neuropeptide Y infusion in animal models of conditioned fear: an NPY Y1 receptor independent effect. Psychopharmacology 206, 291–301 (2009). https://doi.org/10.1007/s00213-009-1610-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-009-1610-8