
Abstract
Rationale and objectives
Understanding the mechanism of relapse provoked by conditioned and unconditioned stimuli is critical to improving treatments for alcoholism. This study compared the reinstatement of alcohol- or sucrose-seeking by conditioned stimuli and priming injections of the neuroactive steroid, allopregnanolone (ALLO).
Methods
Rats were trained to lever-press for 0.1 ml of 10% ethanol or 5% sucrose solutions. Responding was then extinguished, and subjects were tested for reinstatement of lever-press responding. The effects of priming injections of 0, 1.0, 3.0 and 7.5 mg/kg ALLO were determined in subjects trained to self-administer ethanol, and the response-reinstating effects of priming injections of 3.0 mg/kg ALLO were compared with those of conditioned cue presentation in subjects trained to self-administer either ethanol or sucrose.
Results
Priming injections of ALLO dose-dependently reinstated previously extinguished responding for ethanol, as shown by increased responding on the active (ethanol) lever. Contingent presentation of cues previously associated with the reinforcer increased the number of active lever-presses for both ethanol- and sucrose- trained subjects. In contrast, pretreatment with 3.0 mg/kg ALLO increased the number of active lever-presses for subjects that were trained to self-administer ethanol, but not sucrose.
Conclusions
ALLO promotes responding for ethanol, but not sucrose, following a period of abstinence, suggesting that GABAA receptor modulation may contribute to processes involved in reinstatement of ethanol-seeking behavior. In contrast, conditioned stimuli reinstate previously extinguished ethanol- and sucrose-seeking behavior, indicating that the mechanisms that subserve cue-induced reinstatement do not depend upon the nature of the positive reinforcer.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Knowledge of the mechanisms whereby stimuli trigger ethanol-seeking behavior after a period of abstinence is critical for understanding and treating relapse. The ability of conditioned and unconditioned stimuli to induce relapse can be modeled using an operant conditioning procedure in which response reinstatement is provoked in extinguished subjects. Stimuli that increase, or reinstate, instrumental responding above extinction levels include non-contingent presentation of the drug reinforcer, foot shock stress, and exposure to environmental stimuli previously associated with drug-reinforced instrumental responding (for review, see Le and Shaham 2002; Shalev et al. 2002). For example, de Wit and Stewart (1981) found that intravenous cocaine reinstates responding for cocaine. Curiously, when administered via intraperitoneal injection or gavage, ethanol itself is not very effective in inducing reinstatement of responding. Although significant ethanol-induced reinstatement of responding has been reported (Le et al. 1998, 1999; Vosler et al. 2001), the effects are modest (Le and Shaham 2002), and are difficult to reproduce (Nie and Janak, unpublished results).
In addition to establishing that priming injections of drugs of abuse reinstate responding, Stewart and de Wit demonstrated that reinstatement of drug-seeking behavior is elicited by pharmacological agents that mimic the stimulus properties of the drug reinforcer (Stewart and de Wit 1987). Drug discrimination studies have identified multiple neurotransmitter receptors that mediate the stimulus properties of ethanol, including GABAA, NMDA, and 5-HT3 receptors (Grant 1999; Kostowski and Bienkowski 1999). The pharmacological enhancement of GABA-mediated Cl− entry at the GABAA receptor by ethanol is thought to form the basis for the GABAergic component of the ethanol discriminative stimulus complex (Grant 1999).
Like ethanol, the neuroactive steroid, allopregnanolone (3α-hydroxy-5α-pregnan-20-one; ALLO), is a potent modulator of the GABAA receptor complex (Harrison et al. 1987; Baulieu 1992; Paul and Purdy 1992; Lambert et al. 1995). There are many reports of interactions between ALLO and ethanol at the behavioral level that are likely mediated by actions at the GABAA receptor (Grobin et al. 1998). For example, ALLO enhances the ataxic and sedative effects of ethanol (Vanover et al. 1999), and also enhances the augmentation of aggression by low doses of ethanol (Fish et al. 2001). ALLO is reported to inhibit seizures induced during ethanol withdrawal (Devaud et al. 1996; Finn et al. 2000). ALLO also has ethanol-like discriminative stimulus effects in both rats and non-human primates (Ator et al. 1993; Grant et al. 1996).
Our previous study found that exogenous administration of ALLO dose-dependently enhances ethanol-reinforced operant responding (Janak et al. 1998). A recent study has also found that ALLO enhances ethanol consumption in a two-bottle preference test in mice (Sinnott et al. 2002b). The potential role of ALLO in the reinstatement of ethanol self-administration is not known. The first part of this study was designed to test the hypothesis that systemic administration of ALLO would induce reinstatement of responding for ethanol.
Drug-paired stimuli also are reported to induce reinstatement of responding for drugs of abuse. However, studies of cue-induced relapse to ethanol-seeking behavior have reported mixed results (Bienkowski et al. 1999; Katner et al. 1999). A number of studies of other drugs of abuse have found higher rates of responding in reinstatement tests following response-contingent, rather than response-non-contingent, presentation of the cues [Deroche-Gamonet et al. 2002 (see Fig. 2); Meil and See 1996]. Response-contingent presentation of drug-paired cues during the reinstatement test mirrors the temporal relationship between lever press and cue presentation that subjects experience during drug self-administration. However, most previous studies of the reinstatement of ethanol-seeking behavior used non-contingent delivery of conditioned stimuli during the reinstatement test. In the second part of the present study, we tested the hypothesis that reinstatement of ethanol- and sucrose-seeking would be evoked by response-contingent presentation of a tone-light cue complex that was previously paired with reinforcer delivery. We also hypothesized that ALLO would not reinstate responding in sucrose-trained subjects, because of the lack of shared pharmacological actions between sucrose and ALLO.
Materials and methods
Animals
Male Long-Evans rats (Harlan Sprague-Dawley, Indianapolis, Ind., USA), weighing 200–300 g at the beginning of operant training, were housed individually in polycarbonate cages in ventilation racks (Biozone, Fort Mill, S.C., USA). Animals received food ad libitum, and were water restricted during 3 days of forced ethanol consumption (see below) as well as a 2-day period that included lever-press shaping. The room was illuminated on at 12-h light/dark schedule with lights on at 6 a.m. All procedures were conducted during the light cycle between 3 and 6 p.m. These procedures were approved by the Institutional Animal Care and Use Committee of the Ernest Gallo Clinic and Research Center at the University of California, San Francisco, and are in accordance with "PHS Policy on Humane Care and Use of Laboratory Animals", Office of Laboratory Animal Welfare, National Institutes of Health, revised 2002.
Apparatus
The conditioning chambers were 30 cm wide and 29 cm high, and were contained within larger sound-attenuating chambers. Two levers were located against the right wall, 7 cm from the floor and 1 cm from the right or left edge of the right wall, respectively. A 2.5 cm white stimulus light was located above each lever. A rectangular recess (3 cm in diameter) was located between the two levers, 3 cm above the floor. Syringe pumps delivered fluid into a fluid receptacle within this recess. A house light, located on the right wall 14 cm from the floor, was on for the duration of each behavioral session. All behavioral equipment (Med Associates Inc., Georgia, Vt., USA) was computer-controlled via software (Med Associates); the same software recorded the responses and reinforcer deliveries that occurred during behavioral sessions.
Behavioral procedure
Ethanol- or sucrose-reinforced operant responding
Operant responding for ethanol
The specific training procedures used for subjects in experiments 1 and 2 are different and are given in detail in Table 1 and 2, respectively. In brief, at least 5 days after arrival from the vendor, subjects were given 3 days of continuous exposure in the home cage to a solution of 10% ethanol (10E; v/v) in tap water. During these 3 days, plain water was not available. After the pre-exposure phase, subjects were water-deprived for 24 h. Subjects were then placed within the chambers and left overnight (12–14 h). The chambers were equipped with one "active" and one "inactive" lever. Active lever presses were followed by a variable delay (from 0.5 to 1.5 s). The variable delay was followed by onset for 5 s of both the stimulus light above the lever and a tone (2900 Hz, 50 decibels); 2 s after onset of the light and tone, 0.1 ml of 10% sucrose (10S) was delivered from a pump. Inactive lever presses were counted but had no further consequence. At the conclusion of this segment of the training procedure, all subjects received ad libitum water in the home cage from this point forward.
After shaping, subjects began daily 60-min sessions, 5–6 days a week. During this time, and as detailed in Table 1, for subjects in experiment 1 the fixed-ratio (FR) requirement was increased to FR3. These subjects were then exposed to a 10S/10E solution for 3 days; a 10E solution was the reinforcer for each session after that. As described in Table 2, for subjects in experiment 2, a longer period of gradual introduction of ethanol occurred, followed by increases in the FR requirement to FR3. For experiment 2, the number of days at each reinforcer solution was arbitrary and was based on previous experience (Janak et al. 1998). Subjects that on average responded for 15 or fewer lever presses per session (<0.5 ml 10E) were excluded from the study as they were not considered likely to attain pharmacological levels of ethanol (0 out of 8 subjects from experiment 1; 1 out of 10 subjects from experiment 2).
Operant responding for sucrose
Acquisition of lever-press responding for 10% sucrose was conducted as described above for ethanol self-administration. After shaping, subjects began daily 60-min sessions, 6 days a week. The details of the training schedule can be found in Table 2. Briefly, a 10S solution was available on a continuous reinforcement schedule overnight. Then, daily 60-min sessions began, and the reinforcer solution was changed to 5S. One week later, the response requirement was gradually increased until subjects were responding under an FR3 schedule. Subjects responding 15 or fewer times per session were excluded from the analysis (1 of 12 subjects).
Extinction of ethanol- or sucrose-seeking
After 3 weeks of lever-press responding for 10E or 5S, extinction sessions were conducted for all rats during which rats were permitted to respond but no cues or reinforcers were delivered. Reinstatement testing for an individual subject did not begin until that animal reached a criterion of <5 lever responses on either the active or the inactive lever per 60-min session.
Reinstatement induced by allopregnanolone
Subjects were habituated to the injection procedure by administering saline prior to two or three extinction sessions per week for up to 3 weeks, depending on how long an individual subject took to reach criterion. The schedule for testing ALLO's effects was different for experiment 1 and experiment 2, as detailed in Table 1 and Table 2, respectively. For experiment 1, after reaching the extinction criterion, subjects received 1.0, 3.0, or 7.5 mg/kg ALLO or vehicle. Higher doses were not given because we have noted behavioral side-effects of distress such as increased vocalization and tendency to jump from the experimenter's hand following an injection of 10 mg/kg (Janak et al. 1998). Reinstatement was measured by counting the number of lever presses emitted in a single 60-min session. Next, extinction sessions were conducted until each subject again attained criterion, and an additional injection was delivered until the dose-response determination was completed with every subject receiving each dose in a counter-balanced fashion. For subjects in experiment 2, cue-induced reinstatment was tested first; additional extinction sessions were conducted until subjects re-attained the extinction criterion, after which each subject received a vehicle injection followed the next day by an injection of 3.0 mg/kg ALLO.
Reinstatement induced by conditioned stimuli
The ability of conditioned stimuli to reinstate responding by ethanol- or sucrose-trained subjects was tested after extinction by presenting the 5-s tone-light stimulus pair contingent upon active lever pressing (Meil and See 1996), with one tone-light presentation following each active lever press with the same programmed variable delay of 0.5–1.5 s that occurred during ethanol or sucrose self-administration. Reinstatement was measured by counting the number of lever presses emitted in a single 60-min session.
Drugs
Allopregnanolone (3-α-hydroxy-5α-pregnan-20-one; Sigma, St Louis, Mo., USA) was solubilized in a 25% (w/v) solution of 2-hydroxypropyl-β-cyclodextrin (Sigma) in 0.9% saline via sonication for 4–8 h. All injections were subcutaneous (SC) and were made 20 min before the beginning of the behavioral sessions. The injection volume was 1 ml/kg.
Data analysis
The numbers of active and inactive lever-presses for each subject were analyzed using Statistica 5.1 (StatSoft, Inc., Tulsa, Okla., USA). For experiment 1, data were analyzed using ANOVA with repeated measures on Lever type (active versus inactive) and Dose (0, 1.0, 3.0, or 7.5 mg/kg ALLO). For experiment 2, the ANOVA included factors of Reinstatement test (Control session versus Test session) and Lever, as well as the type of Stimulus used to induce reinstatement (ALLO versus Cue). Significant main effects or interactions were followed by paired t-tests. Statistical significance for all tests was set at the P<0.05 level.
Results
Experiment 1
The mean number of active lever presses for eight subjects the last three days prior to extinction was 89±11.6/h, range 32–125. Mean responding at the inactive lever was 2±0.52/h, range 0–4. These values correspond to an estimated ethanol intake of 0.44±0.06, range 0.15–0.66 g/kg. Extinction sessions, during which time ethanol was unavailable, were conducted to a criterion of ≤5 lever responses on either lever per session (mean sessions to criterion=29±4.6).
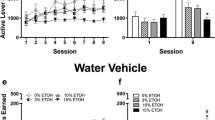
After extinction, injection of ALLO increased lever-press responding. Figure 1 depicts the mean lever presses on the active and inactive levers following ALLO treatment. A within-subject repeated measures ANOVA found main effects of Dose [F(3,21)=3.24, P<0.05] and Lever [active versus inactive; F(1,7)=7.94, P<0.03], and a significant Dose by Lever interaction [F(3,21)=3.77, P<0.03]. Paired t-tests revealed that 3.0 mg/kg ALLO treatment increased responding on the active lever (P<0.05). The increase in responding induced by administration of 7.5 mg/kg ALLO did not reach significance (P=0.06). There were no significant effects of ALLO administration on inactive lever responding (all P>0.05).

Mean number of responses on the active and inactive levers in 1-h sessions following SC administration of vehicle, 1.0, 3.0 or 7.5 mg/kg ALLO by subjects previously trained to lever press for oral ethanol (10E). Error bars represent the standard error of the mean. n=8. *P<0.05, compared to vehicle-active lever presses
Experiment 2
Experiment 2 compared the reinstating effects of priming doses of ALLO and response-contingent cue presentation in extinguished subjects previously trained to self-administer either 10E (n=9) or 5S (n=11). In the ethanol-trained group, the baseline number of active lever presses the last 3 days before extinction was 52±6.7/h, range 31–91. Mean responding at the inactive lever was 2±1.4/h, range 1–4). These values correspond to an estimated g/kg of 0.28±0.04, range 0.17–0.50 g/kg. Baseline active lever responding for the sucrose-reinforced group was 165±42/h, range 24–447. Mean responding at the inactive lever was 3±0.8/h, range 0–8. Mean number of sessions to extinction criterion was 17±3.3 for the ethanol-trained group, and 22±3.0 for the sucrose-trained group.
Both ALLO and conditioned stimuli induced reinstatement of previously extinguished responding in ethanol-trained subjects (Fig. 2A). A three-factor ANOVA found significant main effects of Reinstatement test [Vehicle versus ALLO and Extinction baseline versus Cue; F(1,8)=19.56, P<0.003] and Lever [Active versus Inactive; F(1,8)=20.17, P<0.003], and a significant interaction between the Reinstatement test and the Lever [F(1,8)=9.89, P<0.015]. There was no overall difference in the Stimulus (ALLO versus Cue) used to induce reinstatement and no significant interactions between the Stimulus, Reinstatement test and Lever (all P>0.05), indicating that both response-contingent cue presentation and ALLO injection induced reinstatement of responding for ethanol to a similar degree. Further analysis confirmed that the main effect of Reinstatement test was due to an increase in responding on the active lever [F(1,8)=15.93, P<0.004] but not the inactive lever at test (P>0.05).

Mean number of responses on the active and inactive levers in 1-h sessions following response-contingent presentation of the tone-light stimulus or SC priming injections of 3.0 mg/kg ALLO in subjects previously trained to respond for A ethanol (10E), n=9 or B sucrose (5S), n=11. *P<0.004, compared to extinction- or vehicle-active lever presses. # P<0.02 compared to number of presses under extinction conditions
In contrast to the results for the ethanol-trained subjects, response-contingent cue presentation, but not priming injections of ALLO, induced reinstatement in sucrose-trained subjects [main effect of Stimulus (ALLO versus Cue), F(1,10)=7.31, P<0.025]. Main effects of Reinstatement test [F(1,10)=9.05, P<0.015] and Lever [F(1,10)=14.82, P<0.0035] were also significant, as were all possible two-way interactions (P<0.04), and the three-way interaction of Stimulus by Reinstatement test by Lever [F(1,10)=5.88, P<0.036]. The basis for the significant three-way interaction was examined using two Simple Interactions analyzing the factors of Reinstatement test and Lever within the two Stimulus conditions separately. The interaction between Reinstatement test and Lever was not significant when data from the ALLO-induced reinstatement test were examined (P>0.05), and there was no effect of Reinstatement test (P>0.05). However, analysis of responding following response-contingent presentation of the conditioned tone-light cue revealed a significant interaction between Reinstatement test and Lever [F(1,10)=6.97, P<0.025] that is accounted for by a significant increase in responding at the active (P<0.02) but not the inactive (P=0.21) lever (Fig. 2B).
Discussion
We found that the endogenous neuroactive steroid, ALLO, reinstates responding in an operant conditioning model of ethanol self-administration in which the effects of ALLO were determined after a period of abstinence that corresponded to extinction training. We also found that presentation of conditioned stimuli can induce reinstatement to ethanol in this model as has been reported for other drugs of abuse (Meil and See 1996; Fuchs et al. 1998; Grimm and See 2000; Caggiula et al. 2001; De Vries et al. 2001; Fuchs and See 2002). The reinstatement-inducing effects of ALLO were specific for ethanol, as ALLO had no effect in animals trained to lever-press for sucrose reinforcement. In contrast, the effect of conditioned stimuli was similar in subjects trained under conditions of either ethanol-reinforcement or sucrose-reinforcement.
In both experiments 1 and 2, we found that 3 mg/kg ALLO increased responding on the active (ethanol-reinforced), but not inactive, lever after extinction. Therefore, although ALLO enhances ethanol's actions at the GABAA receptor (Majewska 1988), ALLO alteration of ethanol-seeking behavior does not always require the simultaneous presence of ethanol. Previously, we found significant increases in responding following administration of the same dose of ALLO, but not 1 or 10 mg/kg, on operant ethanol self-adminstration (Janak et al. 1998). Therefore it is possible that ALLO generally enhances appetitive behavior as measured by lever presses for ethanol whether ethanol is present in the bloodstream or not. ALLO also has direct effects on consumption of ethanol, as recently demonstrated by Sinnott et al. (2002b), who found that ALLO enhances ethanol intake in mice when ethanol was available in the home cage. These findings indicate that ALLO's effects on operant ethanol self-administration may also reflect direct effects on consumption. Whether increases in ethanol consumption reflect enhancement or attenuation of ethanol's reinforcing effects is not yet known (Sinnott et al. 2002b).
Although ethanol need not be present for ALLO to enhance ethanol-seeking behavior in the reinstatement model, the lack of effect in the sucrose group suggests that reinstatement of responding induced by ALLO depends upon previous experience with the pharmacological actions of ethanol. The specific reinstatement by ethanol-trained subjects seen here is interesting in light of the enhancement of non-alcohol consumption that has been demonstrated following administration of ALLO (Sinnott et al. 2002b) and other GABAA modulators (Cooper and Yerbury 1988; Shelton and Balster 1997). It may be that ALLO has effects upon consumption of reinforcing substances that are distinct from the effect on reinstatement.
Ethanol intakes across individual subjects varied greatly, as is typical in an outbred rat population. Although the intakes reported here do not produce blood levels that lead to overt physical dependence, we (unpublished results) and others find that similar intakes have been found repeatedly to produce measurable blood alcohol levels (cf. Koob 2000), and probably correspond to levels attained by moderate drinkers. Importantly, our results support the notion that subjects in both studies reported here achieved familiarity with the pharmacological effects of ethanol. If this were not the case, then it would be difficult to explain the differential effect of ALLO on reinstatement for ethanol and sucrose.
It is likely that the specificity of the effects of ALLO for ethanol-training depends upon their overlapping pharmacological mechanisms of action. Both ethanol and ALLO enhance GABAA-receptor mediated Cl− flux. Ethanol's reinforcing effects appear to be mediated at least in part by its effects upon GABAergic systems (for review, see Koob et al. 1998; McBride et al. 1999), and the oral self-administration of ALLO itself has recently been demonstrated (Sinnott et al. 2002a). In addition, ALLO is reported to induce a place preference (Finn et al. 1997; but see Beauchamp et al. 2000). Therefore, it is possible that ALLO itself can serve as a reinforcer, and that some GABAergic neural mechanisms that underlie ethanol reinforcement and ALLO reinforcement overlap. In addition, their shared GABAergic pharmacological effects may form the basis for the findings that ALLO substitutes for ethanol in drug discrimination paradigms in both rats and primates (Ator et al. 1993; Grant et al. 1996). Previous studies have suggested that drugs that share aspects of their stimulus properties with a given drug of abuse may induce relapse in a manner similar to non-contingent administration of that drug of abuse itself (Stewart and de Wit 1987). Therefore, it may be that ALLO reinstates responding for ethanol because ALLO shares some of the stimulus and/or reinforcing properties of ethanol. However, not all drugs that substitute for ethanol in drug discrimination studies also induce reinstatement of responding for ethanol; Vosler et al. (2001) found that the NMDA-antagonist dizocilpine did not reinstate responding for ethanol. Therefore sharing stimulus properties with ethanol is not a sufficient condition for a compound to induce ethanol reinstatement.
The effects of ALLO on ethanol-seeking behavior are of particular interest because ALLO is one of a number of endogenous neuroactive steroids synthesized from circulating and/or local progesterone in the CNS of male and female mammals. Our findings suggest the possibility that fluctuations in endogenous CNS levels of ALLO could contribute to relapse to ethanol, although further work is necessary to determine if systemic injection of the effective dose of 3 mg/kg results in physiological or supra-physiological CNS levels of ALLO.
Previous studies of cocaine (Meil and See 1996; Fuchs et al. 1998; Tran-Nguyen et al. 1998; Grimm and See 2000; De Vries et al. 2001; Alleweireldt et al. 2002), heroin (Fuchs and See 2002), nicotine (Caggiula et al. 2001) and "speedball" (cocaine and heroin together; Highfield et al. 2001) self-administration by rats have found that response-contingent presentation of drug-paired stimuli reliably induce reinstatement after extinction. The current results extend these findings to include ethanol. In contrast, we found no significant change in active lever responding after non-contingent presentation of an ethanol-paired tone to extinguished rats (Nie and Janak, unpublished observations), in agreement with findings from cocaine self-administration studies (Meil and See 1996; Fuchs et al. 1998).
In conclusion, the neuroactive steroid, ALLO, reinstates previously-extinguished responding in ethanol-trained rats, but has no effect in sucrose-trained rats, suggesting that ALLO may contribute to biological processes underlying relapse to ethanol. Cue-induced reinstatement of responding is observed in subjects trained to self-administer either ethanol or sucrose. Future studies designed to elucidate the neural circuits that underlie conditioned cue- and neuroactive steroid-induced reinstatement of responding for ethanol will contribute to our understanding of the biological processes that control ethanol-seeking behavior.
References
Alleweireldt AT, Weber SM, Kirschner KF, Bullock BL, Neisewander JL (2002) Blockade or stimulation of D1 dopamine receptors attenuates cue reinstatement of extinguished cocaine-seeking behavior in rats. Psychopharmacology 159:284–293
Ator NA, Grant KA, Purdy RH, Paul SM, Griffiths RR (1993) Drug discrimination analysis of endogenous neuroactive steroids in rats. Eur J Pharmacol 241:237–243
Baulieu EE (1992) Neurosteroids: an overview. Adv Biochem Psychopharmacol 47:1–16
Beaman CM, Hunter GA, Dunn LL, Reid LD (1984) Opioids, benzodiazepines and intake of ethanol. Alcohol 1:39–42
Beauchamp MH, Ormerod BK, Jhamandas K, Boegman RJ, Beninger RJ (2000) Neurosteroids and reward: allopregnanolone produces a conditioned place aversion in rats. Pharmacol Biochem Behav 67:29–35
Bienkowski P, Kostowski W, Koros E (1999) The role of drug-paired stimuli in extinction and reinstatement of ethanol-seeking behaviour in the rat. Eur J Pharmacol 374:315–319
Caggiula AR, Donny EC, White AR, Chaudhri N, Booth S, Gharib MA, Hoffman A, Perkins KA, Sved AF (2001) Cue dependency of nicotine self-administration and smoking. Pharmacol Biochem Behav 70:515–530
Cooper SJ, Yerbury RE (1988) Clonazepam selectively increases saccharin ingestion in a two-choice test. Brain Res 456:173–176
De Vries TJ, Shaham Y, Homberg JR, Crombag H, Schuurman K, Dieben J, Vanderschuren LJ, Schoffelmeer AN (2001) A cannabinoid mechanism in relapse to cocaine seeking. Nat Med 7:1151–1154
de Wit H, Stewart J (1981) Reinstatement of cocaine-reinforced responding in the rat. Psychopharmacology 75:134–143
Deroche-Gamonet V, Piat F, Le Moal M, Piazza PV (2002) Influence of cue-conditioning on acquisition, maintenance and relapse of cocaine intravenous self-administration. Eur J Neurosci 15:1363–1370
Devaud LL, Purdy RH, Finn DA, Morrow AL (1996) Sensitization of gamma-aminobutyric acidA receptors to neuroactive steroids in rats during ethanol withdrawal. J Pharmacol Exp Ther 278:510–517
Finn DA, Phillips TJ, Okorn DM, Chester JA, Cunningham CL (1997) Rewarding effect of the neuroactive steroid 3 alpha-hydroxy-5 alpha-pregnan-20-one in mice. Pharmacol Biochem Behav 56:261–264
Finn DA, Gallaher EJ, Crabbe JC (2000) Differential change in neuroactive steroid sensitivity during ethanol withdrawal. J Pharmacol Exp Ther 292:394–405
Fish EW, Faccidomo S, DeBold JF, Miczek KA (2001) Alcohol, allopregnanolone and aggression in mice. Psychopharmacology 153:473–483
Fuchs RA, See RE (2002) Basolateral amygdala inactivation abolishes conditioned stimulus- and heroin-induced reinstatement of extinguished heroin-seeking behavior in rats. Psychopharmacology 160:425–433
Fuchs RA, Tran-Nguyen LT, Specio SE, Groff RS, Neisewander JL (1998) Predictive validity of the extinction/reinstatement model of drug craving. Psychopharmacology 135:151–160
Grant KA (1999) Strategies for understanding the pharmacological effects of ethanol with drug discrimination procedures. Pharmacol Biochem Behav 64:261–267
Grant KA, Azarov A, Bowen CA, Mirkis S, Purdy RH (1996) Ethanol-like discriminative stimulus effects of the neurosteroid 3 alpha-hydroxy-5 alpha-pregnan-20-one in female Macaca fascicularis monkeys. Psychopharmacology 124:340–346
Grimm JW, See RE (2000) Dissociation of primary and secondary reward-relevant limbic nuclei in an animal model of relapse. Neuropsychopharmacology 22:473–479
Grobin AC, Matthews DB, Devaud LL, Morrow AL (1998) The role of GABAA receptors in the acute and chronic effects of ethanol. Psychopharmacology 139:2–19
Harrison NL, Majewska MD, Harrington JW, Barker JL (1987) Structure-activity relationships for steroid interaction with the gamma-aminobutyric acidA receptor complex. J Pharmacol Exp Ther 241:346–353
Highfield D, Yap J, Grimm JW, Shalev U, Shaham Y (2001) Repeated lofexidine treatment attenuates stress-induced, but not drug cues-induced reinstatement of a heroin-cocaine mixture (speedball) seeking in rats. Neuropsychopharmacology 25:320–331
Janak PH, Redfern JE, Samson HH (1998) The reinforcing effects of ethanol are altered by the endogenous neurosteroid, allopregnanolone. Alcohol Clin Exp Res 22:1106–1112
Katner SN, Magalong JG, Weiss F (1999) Reinstatement of alcohol-seeking behavior by drug-associated discriminative stimuli after prolonged extinction in the rat. Neuropsychopharmacology 20:471–479
Koob GF (2000) Animal models of craving for ethanol. Addiction 95: S73–S81
Koob GF, Sanna PP, Bloom FE (1998) Neuroscience of addiction. Neuron 21:467–476
Kostowski W, Bienkowski P (1999) Discriminative stimulus effects of ethanol: neuropharmacological characterization. Alcohol 17:63–80
Lambert JJ, Belelli D, Hill-Venning C, Peters JA (1995) Neurosteroids and GABAA receptor function. Trends Pharmacol Sci 16:295–303
Le A, Shaham Y (2002) Neurobiology of relapse to alcohol in rats. Pharmacol Ther 94:137–156
Le AD, Quan B, Juzytch W, Fletcher PJ, Joharchi N, Shaham Y (1998) Reinstatement of alcohol-seeking by priming injections of alcohol and exposure to stress in rats. Psychopharmacology 135:169–174
Le AD, Poulos CX, Harding S, Watchus J, Juzytsch W, Shaham Y (1999) Effects of naltrexone and fluoxetine on alcohol self-administration and reinstatement of alcohol seeking induced by priming injections of alcohol and exposure to stress. Neuropsychopharmacology 21:435–444
Majewska MD (1988) Interaction of ethanol with the GABAA receptor in the rat brain: possible involvement of endogenous steroids. Alcohol 5:269–273
McBride WJ, Murphy JM, Ikemoto S (1999) Localization of brain reinforcement mechanisms: intracranial self- administration and intracranial place-conditioning studies. Behav Brain Res 101:129–152
Meil WM, See RE (1996) Conditioned cued recovery of responding following prolonged withdrawal from self-administered cocaine in rats: an animal model of relapse. Behav Pharmacol 7:754–763
Paul SM, Purdy RH (1992) Neuroactive steroids. FASEB J 6:2311–2322
Shalev U, Grimm JW, Shaham Y (2002) Neurobiology of relapse to heroin and cocaine seeking: a review. Pharmacol Rev 54:1–42
Shelton KL, Balster RL (1997) Effects of gamma-aminobutyric acid agonists and N-methyl-d-aspartate antagonists on a multiple schedule of ethanol and saccharin self-administration in rats. J Pharmacol Exp Ther 280:1250–1260
Sinnott RS, Mark GP, Finn DA (2002a) Reinforcing effects of the neurosteroid allopregnanolone in rats. Pharmacol Biochem Behav 72:923–929
Sinnott RS, Phillips TJ, Finn DA (2002b) Alteration of voluntary ethanol and saccharin consumption by the neurosteroid allopregnanolone in mice. Psychopharmacology 162:438–447
Stewart J, de Wit H (1987) Reinstatement of drug-taking behavior as a method of assessing incentive motivational properties of drug. In: Bozarth MA (ed) Methods of assessing the reinforcing properties of abused drugs. Springer, New York, pp 211–227
Tran-Nguyen LT, Fuchs RA, Coffey GP, Baker DA, O'Dell LE, Neisewander JL (1998) Time-dependent changes in cocaine-seeking behavior and extracellular dopamine levels in the amygdala during cocaine withdrawal. Neuropsychopharmacology 19:48–59
Vanover KE, Suruki M, Robledo S, Huber M, Wieland S, Lan NC, Gee KW, Wood PL, Carter RB (1999) Positive allosteric modulators of the GABA(A) receptor: differential interaction of benzodiazepines and neuroactive steroids with ethanol. Psychopharmacology 141:77–82
Vosler PS, Bombace JC, Kosten TA (2001) A discriminative two-lever test of dizocilpine's ability to reinstate ethanol-seeking behavior. Life Sci 69:591–598
Acknowledgements
This study was supported by award no. DAMD17-01-1-0739 to P.H.J. awarded and administered by the US Army Medical Research Acquisition Activity, 820 Chandler Street, Fort Detrick, MD 21702, USA. The content of information herein does not reflect the position or policy of the Government and no official endorsement should be inferred. The study was also supported by funds from the State of California for Medical Research on Alcohol and Substance Abuse through the University of California at San Francisco. We are indebted to Dr. Yavin Shaham for expert advice on reinstatement paradigms.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Nie, H., Janak, P.H. Comparison of reinstatement of ethanol- and sucrose-seeking by conditioned stimuli and priming injections of allopregnanolone after extinction in rats. Psychopharmacology 168, 222–228 (2003). https://doi.org/10.1007/s00213-003-1468-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-003-1468-0