
Abstract
Phytophthora root rot (PRR), caused by Phytophthora sojae Kaufmann & Gerdemann, is one of the most destructive diseases of soybean [Glycine max (L.) Merr.]. Deployment of resistance genes is the most economical and effective way of controlling the disease. The soybean cultivar ‘Yudou 29’ is resistant to many P. sojae isolates in China. The genetic basis of the resistance in ‘Yudou 29’ was elucidated through an inheritance study and molecular mapping. In response to 25 P. sojae isolates, ‘Yudou 29’ displayed a new resistance reaction pattern distinct from those of differentials carrying known Rps genes. A population of 214 F2:3 families from a cross between ‘Jikedou 2’ (PRR susceptible) and ‘Yudou 29’ was used for Rps gene mapping. The segregation fit a ratio of 1:2:1 for resistance:segregation:susceptibility within this population, indicating that resistance in ‘Yudou 29’ is controlled by a single dominant gene. This gene was temporarily named RpsYD29 and mapped on soybean chromosome 03 (molecular linkage group N; MLG N) flanked by SSR markers SattWM82-50 and Satt1k4b at a genetic distance of 0.5 and 0.2 cM, respectively. Two nucleotide binding site-leucine rich repeat (NBS-LRR) type genes were detected in the 204.8 kb region between SattWM82-50 and Satt1k4b. These two genes showed high similarity to Rps1k in amino acid sequence and could be candidate genes for PRR resistance. Based on the phenotype reactions and the physical position on soybean chromosome 03, RpsYD29 might be a novel allele at, or a novel gene tightly linked to, the Rps1 locus.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Phytophthora root rot (PRR), caused by Phytophthora sojae Kaufmann & Gerdemann, is one of the most destructive diseases of soybean [Glycine max (L.) Merr.] (Bernard et al. 1957; Kuan and Erwin 1980). This disease has been reported in most soybean-production regions of the world, since it was first noted in America in 1948 (Tyler 2007; Wrather et al. 2001). In China, PRR was first observed in 1991 in Heilongjiang Province (Shen and Su 1991). Since then, the disease has become widespread in Heilongjiang Province, and the annual infected area is estimated at over 150,000 ha (Zhang et al. 2010). It has now also spread to Fujian Province (Chen et al. 2004).
PRR can be effectively controlled through the use of complete or partial resistance genes (Sugimoto et al. 2012). Partial resistance to P. sojae can be ineffective when plants are subjected to high disease pressure (Dorrance et al. 2003), so the deployment of race-specific, complete resistance genes in soybean cultivars has been the primary method used to control PRR (Schmitthenner 1999). To date, 14 genes conferring resistance to P. sojae (Rps) have been identified at 8 loci, Rps1 to Rps8, which were mapped to molecular linkage groups (MLG) N, J, F, G, G, G, N and F, respectively (Sandhu et al. 2005). Five functional genes Rps1a, Rps1b, Rps1c, Rps1d and Rps1k have been mapped at the Rps1 locus (Bernard et al. 1957; Buzzell and Anderson 1992; Mueller et al. 1978), and three alleles Rps3a, Rps3b and Rps3c have been mapped at the Rps3 locus (Mueller et al. 1978; Ploper et al. 1985). Additionally, five Rps genes, RpsYB30, RpsYD25 (Rpsyu25), RpsZS18, RpsSN10 and the Rps gene in soybean cultivar ‘Waseshiroge’, were mapped to MLG L, N, D1b, F and N, respectively (Fan et al. 2009; Sugimoto et al. 2011; Sun et al. 2011; Yao et al. 2010; Yu et al. 2010; Zhu et al. 2007).
Among these Rps genes, Rps1k is of great interest because it confers stable, broad-spectrum resistance in the USA (Schmitthenner et al. 1994), although new Rps1k-virulent P. sojae isolates have been reported in some areas (Dorrance et al. 2003; Schmitthenner 1999). This gene was first identified in cultivar ‘Kingwa’ (Bernard and Cremeens 1981) and has been transferred to soybean cultivars in the USA (Slaminko et al. 2010). The BAC-cloned Rps1k region contains two classes of functional coiled coil-nucleotide binding-leucine rich repeat genes (Rps1k-1 and Rps1k-2) and repetitive sequences (Gao et al. 2005; Gao and Bhattacharyya 2008). In addition to Rps1k, the Rps1c and Rps1a genes are also frequently introduced into commercial cultivars in the USA (Slaminko et al. 2010). However, the continuous use of the Rps genes in soybean cultivars has created selection pressure for the evolution of new pathogenic P. sojae isolates that can overcome the resistance conferred by these genes.
Since 1955, at least 55 P. sojae races with virulence pathotype varying from 7 to 1a, 1b, 1c, 1d, 1k, 3a, 6 and 7 were identified in the USA (Grau et al. 2004). P. sojae also presents similarly abundant virulence diversity in China (Zhang et al. 2010; Zhu et al. 2003). So far, 12 P. sojae races (1, 3, 4, 5, 9, 11, 13, 17, 21, 24, 44 and 54) were found in Heilongjiang Province, China (Zhang et al. 2010). In addition, other virulence pathotypes that differ from the 55 P. sojae races in the USA were also found in China (Zhu et al. 2003). Most of the soybean cultivars or germplasms containing the known Rps genes, except Rps1a, Rps1c and Rps1k, are not effective at controlling PRR in the major soybean-production regions (Cui et al. 2010; Tang et al. 2010; Zhang et al. 2010; Zhu et al. 2003). Although new races of P. sojae appear with the release of resistant cultivars, the use of genetic resistance still remains the most effective strategy to reduce losses caused by the pathogen. Thus, it is necessary to identify new Rps genes and develop molecular markers to genes of interest for marker-assisted selection (MAS).
In our previous study, we found that the cultivar ‘Yudou 29’ had a broad-spectrum resistance (Chen et al. 2008). The objectives of this study were to further characterize the inheritance of the Rps gene(s) in the cultivar ‘Yudou 29’, identify SSR markers for fine mapping and predict the candidate gene(s).
Materials and methods
Plant materials
‘Yudou 29’ is a PRR-resistant cultivar, and ‘Jikedou 2’ is a PRR susceptible cultivar. A population of 214 families derived from a cross between ‘Yudou 29’ and ‘Jikedou 2’ was used for gene mapping. The F1 plants from the cross were self-pollinated to produce a population of F2 plants. Each F2 plant was self-pollinated and threshed individually to yield seeds of the F2:3 families for both genotype and phenotype evaluations.
To determine which Rps gene or Rps gene combination was present in ‘Yudou 29’, phenotype analysis was performed using 15 differentials carrying a single Rps gene: Harlon (Rps1a), Harosoy13XX (Rps1b), Williams79 (Rps1c), PI103091 (Rps1d), Williams82 (Rps1k), L76-988 (Rps2), L83-570 (Rps3a), PRX146-36 (Rps3b), PRX145-48 (Rps3c), L85-2352 (Rps4), L85-3059 (Rps5), Harosoy62XX (Rps6), Harosoy (Rps7), PI399073 (Rps8) and Yudou25 (RpsYD25). Williams (rps) was used as the susceptible check to indicate successful inoculation.
Phytophthora sojae inoculation and PRR evaluation
A total of 25 P. sojae isolates with different virulence were used in the phenotype test (Table 1). For each isolate inoculation, 12 seeds of each differential were planted per replication in a pot with a diameter of 10 cm, and all differentials were tested in 3 replications using the hypocotyl-inoculation technique (Haas and Buzzell 1976). To test the phenotypes of the population, the isolate PsMC1 was used to inoculate ‘Yudou 29’, ‘Jikedou 2’, and 30 individual seedlings of each F2:3 family using the same technique as above.
After inoculation, the plants were placed in a mist room with relative humidity 100 % and an average temperature of 25 °C for 2 days. They were then placed in a greenhouse with an average temperature of 25 °C. Reactions were recorded as the percentage of dead seedlings in each F2:3 family at 6 days post-inoculation (DPI). In accordance with Gordon et al. (2006), families with 0–20 % dead seedlings were scored as homozygous resistant (R), families with 80–100 % dead seedlings were considered homozygous susceptible (S), and families with 21–79 % of dead seedlings was scored as segregating (Rs).
DNA preparation and pooling for bulk segregation analysis
DNA was extracted from soybean leaf tissues collected from 15–30 seedlings of each family using the CTAB method with minor modifications (Allen et al. 2006). Resistant and susceptible bulks for the bulk segregation analysis (BSA) were prepared from DNA samples of either ten homozygous-resistant or ten homozygous-susceptible F2:3 families (Michelmore et al. 1991). DNA bulks were prepared by pooling 1 μg DNA of each selected family. The final concentration of the DNA bulks was adjusted to 50 ng/μl.
SSR marker development and analysis
The Rps gene in ‘Yudou 29’ was first mapped with the SSR markers described in Soybase (http://soybase.org). Since the G. max sequences (http://www.phytozome.net/soybean) have been released (Schmutz et al. 2010), new markers could be developed to finely map the gene. Thus, new SSR markers between Sat_186 and Satt530 were designed with Primer Premier 5.0 (Premier Biosoft International, Palo Alto, CA, USA) with default parameters according to the sequence downloaded from Phytozome (http://www.phytozome.net/soybean), in which the simple repeat sequences were identified by SSR Hunter 1.3 (http://en.bio-soft.net/dna/SSRHunter). Because Rps1k is located at the Rps1 locus on MLG N (chromosome 03) and has been sequenced, new SSR markers were also designed from the contig containing the Rps1k-1, Rps1k-2 and Rps1k-3 (GenBank accession no. EU450800).
PCR reactions were performed on a thermal cycler (Biometra, USA) in 20 μl reaction volumes containing 50 ng genomic DNA, 2.5 μl 10× PCR reaction buffer (2.0 mM MgCl2), 0.2 mM each dNTP, 1.0 U of Taq DNA polymerase and 0.2 μM of each primer. The PCR amplification consisted of an initial denaturation at 94 °C for 5 min, followed by 35 cycles of denaturation at 94 °C for 45 s, annealing at 49–52 °C for 45 s and extension at 72 °C for 45 s, and final extension at 72 °C for 10 min. PCR products were mixed with 4 μl of 6× loading buffer (0.25 % bromophenol blue, 0.25 % xylene cyanol FF and 40 % sugar) and were separated by 6 % polyacrylamide sequencing gel. The BSA polymorphic SSR markers were further tested for the entire F2:3 mapping population.
Data analysis and linkage map construction
Resistant (R) and intermediate (I) phenotypes to the isolates (resistant reactions) were recorded as “1” for presence, and susceptible phenotypes were recorded as “0”. Cluster analysis was performed with different modules of the NTSYS-PC software, version 2.11L (Rohlf 2000).
The segregation patterns of phenotypes and selected SSR markers in the mapping population were tested for goodness-of-fit to Mendelian segregation ratio with Chi-square (χ 2) analysis. A genetic linkage map of RpsYD29 was constructed with the Joinmap 4.0 linkage analysis software (Van Ooijen 2006). Linkage groups were determined using a log-likelihood (LOD) threshold of 3.0.
Results
Phenotype reaction of ‘Yudou 29’ to P. sojae isolates
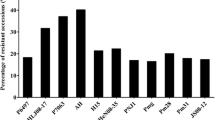
‘Yudou 29’ was resistant to 19 of the 25 isolates tested, while the 15 differentials carrying a single known Rps gene were resistant to 8–18 isolates. The reactions of ‘Yudou 29’ differed from those of Harlon, Harosoy13XX, Williams79, PI103091 and Williams82 to 17, 10, 10, 11 and 13 isolates, respectively. The reaction pattern of ‘Yudou 29’ to the 25 P. sojae isolates did not resemble any of the 15 differentials. The genetic diversity and phenotypic relationships between ‘Yudou 29’ and the 15 differentials in this study were depicted by cluster analysis (Fig. 1). Cutting at the coefficient 0.726, ‘Yudou 29’ comprised a single subgroup, which suggested that it might contain a novel Rps gene.

Dendrogram revealed by UPGMA cluster analysis of the Phytophthora resistance among ‘Yudou 29’, 15 differentials carrying a single Rps gene: Harlon (Rps1a), Harosoy13XX (Rps1b), Williams79 (Rps1c), PI103091 (Rps1d), Williams82 (Rps1k), L76-988 (Rps2), L83-570 (Rps3a), PRX146-36 (Rps3b), PRX145-48 (Rps3c), L85-2352 (Rps4), L85-3059 (Rps5), Harosoy62XX (Rps6), Harosoy (Rps7), PI399073 (Rps8) and Yudou25 (RpsYD25) and the susceptible cultivar Williams
Phenotype analysis for the mapping population
‘Jikedou 2’ plants inoculated with PsMC1 displayed severe rot at the inoculation site and all plants were dead at 6 DPI. ‘Yudou 29’ plants showed no symptoms at 6 DPI, and thus were resistant to the isolate PsMC1.
Among the 214 F2:3 families of the mapping population, 51 were homozygous resistant (R), 60 were homozygous susceptible (S) and 103 were segregating (Rs) to the isolate PsMC1 (Table 2). A segregation ratio of 51:103:60 in the F2:3 population fits well with the genetic model ratio 1:2:1 (χ 2 = 1.06, p = 0.62). This result suggested that Phytophthora resistance in ‘Yudou 29’ was controlled by a single dominant gene, which was temporarily named RpsYD29.
Molecular mapping of the RpsYD29 gene
Using the BSA method, five SSR markers, Satt152, Satt159, Sat_186, Satt631 and Satt530 on chromosome 03 (MLG N), showed polymorphisms between ‘Yudou 29’ and ‘Jikedou 2’, as well as between the resistant bulk and the susceptible bulk. Linkage analysis further revealed that RpsYD29 was linked to these five SSR markers and located between Sat_186 and Satt530 (Fig. 2).

Simple sequence repeat (SSR)-based genetic linkage map of the RpsYD29 region. The map was deduced from segregation analysis of 214 F2:3 families from a cross between Glycine max cultivars ‘Yudou 29’ and ‘Jikedou 2’. The map was generated in Joinmap using Kosambi’s mapping function. Marker names are on the right and distances (cM) on the left
A 2.34 Mb DNA segment of soybean chromosome 03 between Sat_186 and Satt530 was identified using a “BLAST genome” search in the Phytozome soybean genome database. Within this region, 218 SSR loci were identified by SSR hunter. A total of 156 SSR loci with 15 bp or greater repeat motifs were selected to design primers for fine mapping of RpsYD29. Three SSR markers, SattWM82-32, SattWM82-39 and SattWM82-50, showed polymorphisms between ‘Yudou 29’ and ‘Jikedou 2’ (Supplementary Table 1).
In addition, 11 SSR markers were designed from the 184 kb contig containing the Rps1k region (GenBank accession no. EU450800), three of which (Satt1k2a, Satt1k3 and Satt1k4b) were polymorphic between ‘Yudou 29’ and ‘Jikedou 2’ (Supplementary Table 1).
Molecular analysis of the 214 F2:3 families using these 11 polymorphic markers revealed that their segregation pattern fits the 1:2:1 ratio (Supplementary Table 2). RpsYD29 was closely linked to the SSR markers SattWM82-50 and Satt1k4b at a distance of 0.5 and 0.2 cM, respectively, and a genetic map consisting of the 11 SSR markers and RpsYD29 was constructed (Fig. 2).
Candidate gene prediction
A BLAST search showed that the physical distance of the region between markers SattWM82-50 and Satt1k4b, which are at nucleotide positions 3,857,715 and 4,062,474, respectively, is approximately 204.8 kb. According to the Glyma1.0 annotations, a total of 11 genes have been identified in this region, of which 9 are supported by ESTs and two are putative (Supplementary Table 3).
Nucleotide binding site-leucine rich repeat (NBS-LRR) type resistance genes have been implicated in the resistance of Rps1k (Gao and Bhattacharyya 2008). Therefore, the identified 204.8 kb region containing RpsYD29 was strictly evaluated for this motif, and two full-length NBS-LRR type genes, Glyma03g04030.1 and Glyma03g04080.1, were found to share amino acid sequence identities of 62–75 % with Rps1k-1 and Rps1k-2.
Discussion
The soybean originated in China and large quantities of Chinese germplasms have been screened for PRR resistance (Tang et al. 2010; Xia et al. 2011; Zhu et al. 2006). All identified Rps genes (except Rps8) and Rps gene combinations are present in germplasms from China (Kyle et al. 1998; Lohnes et al. 1994, 1996) and many germplasms are postulated to carry novel Rps genes (Xia et al. 2011; Zhu et al. 2006).
In this study, we investigated the resistance phenotype and genomic location of the RpsYD29 gene in ‘Yudou 29’. The results confirm that RpsYD29 is a strong, broad-spectrum resistance gene that is likely a novel allele at the Rps1 locus, or a novel gene at another locus tightly linked to Rps1 locus.
In addition to RpsYD29, eight known Rps genes were previously mapped to MLG N, including five alleles at the Rps1 locus (Bernard et al. 1957; Buzzell and Anderson 1992; Mueller et al. 1978), Rps7 (Weng et al. 2001), RpsYD25 (Rpsyu25) (Fan et al. 2009; Sun et al. 2011) and an Rps gene in cv. Waseshiroge (Sugimoto et al. 2011). In our cluster analysis based on the Phytophthora resistance to 25 P. sojae isolates, ‘Yudou 29’ and the differentials carrying Rps1 alleles (1a, 1b, 1c, 1d, 1k), Rps7 and RpsYD25 fell into different subgroups cutting at the coefficient 0.726, and ‘Yudou 29’ was clustered in a subgroup different from all these genes (except Rps1d) on MLG N even cutting at 0.518. Sugimoto et al. (2008) stated that Rps1d was located “above” Sat_186, and was flanked by Satt152 (11.5 cM) and Sat_186 (5.7 cM). In the present study, RpsYD29 was located “below” Sat_186, and was flanked by Sat_186 (2.9 cM) and Satt530 (2.5 cM). These indicated that RpsYD29 is a distinct gene from the Rps1 alleles, Rps7 and RpsYD25. In addition, the Rps gene in Waseshiroge was located between Satt009 and T003044871 and may reside in the nucleotide region between 3,919,203 and 4,486,048 of the chromosome 03 (MLG N) (Sugimoto et al. 2011). RpsYD29 is not at the same location as the Rps gene in Waseshiroge and thus is distinct from this gene as well.
Carrying the novel resistance gene, RpsYD29, conveying broad-spectrum resistance to P. sojae isolates, ‘Yudou 29’ could be used as a new source of resistance for breeding Phytophthora-resistant cultivars. The markers identified in this study will facilitate the tracking of RpsYD29 in progenies or cultivars for reliable MAS in breeding programs. The SSR markers SattWM82-50 and Satt1k4b positioned RpsYD29 within very tight genetic distances of 0.5 and 0.2 cM, respectively. The accuracy of genotyping was theoretically estimated to be 99.60 and 99.80 % for MAS with SattWM82-50 and Satt1k4b, respectively. The selection accuracy would be as high as 99.99 % for MAS using both markers.
References
Allen GC, Flores-Vergara MA, Krasynanski S, Kumar S, Thompson WF (2006) A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethylammonium bromide. Nat Protoc 1:2320–2325
Bernard RL, Cremeens CR (1981) An allele at the Rps1 locus from the variety Kingwa. Soybean Genet Newsl 8:40–42
Bernard RL, Smith PE, Kaufmann MJ, Schmitthenner AF (1957) Inheritance of resistance to Phytophthora root rot and stem rot in the soybean. Agron J 49:391
Buzzell RI, Anderson TR (1992) Inheritance and race reaction of a new soybean Rps1 allele. Plant Dis 76:600–601
Chen QH, Wang QY, Wang YC, Zheng XB (2004) Identification and sequencing of ribosomal DNA-ITS of Phytophthora sojae in Fujian. Acta Phytopathol Sinica 34:112–116
Chen XL, Zhu ZD, Wang XM, Xiao YN, Wu XF (2008) Postulation of Phytophthora resistance genes in soybean cultivars or lines. Sci Agric Sinica 41:1227–1234
Cui KL, Yin WX, Tang QH, Dong SM, Zheng XB, Zhang ZG, Wang YC (2010) Distribution, pathotypes, and metalaxyl sensitivity of Phytophthora sojae from Heilongjiang and Fujian provinces in China. Plant Dis 94:881–884
Dorrance AE, McClure SA, St Martin SK (2003) Effect of partial resistance on Phytophthora stem rot incidence and yield of soybean in Ohio. Plant Dis 87:308–312
Fan AY, Wang XM, Fang XP, Wu XF, Zhu ZD (2009) Molecular identification of Phytophthora resistance gene in soybean cultivar Yudou 25. Acta Agron Sinica 35:1844–1850
Gao H, Bhattacharyya MK (2008) The soybean-Phytophthora resistance locus Rps1-k encompasses coiled coil-nucleotide binding-leucine rich repeat-like genes and repetitive sequences. BMC Plant Biol 8:1–14
Gao H, Narayanan NN, Ellison L, Bhattacharyya MK (2005) Two classes of highly similar coiled coil-nucleotide binding-leucine rich repeat genes isolated from the Rps1-k locus encode Phytophthora resistance in soybean. Mol Plant Microbe Interact 18:1035–1045
Gordon SG, St Martin SK, Dorrance AE (2006) Rps8 maps to a resistance gene rich region on soybean molecular linkage group F. Crop Sci 46:168–173
Grau CR, Dorrance AE, Bond J, Russin J (2004) Fungal diseases. In: Boerma HR, Specht JE (eds) Soybeans: Improvement, production and uses. Agronomy Monograph, 3rd edn. American Society of Agronomy, Madison, pp 679–763
Haas JH, Buzzell RI (1976) New races 5 and 6 of Phytophthora megasperma var. sojae and differential reactions of soybean cultivars for races 1 and 6. Phytopathology 66:1361–1362
Kuan TL, Erwin DC (1980) Formae speciales differentiation of Phytophthora megasperma isolates from soybeans and alfalfa. Phytopathology 70:333–338
Kyle DE, Nickell CD, Nelson RL, Pedersen WL (1998) Response of soybean accessions from provinces in southern China to Phytophthora sojae. Plant Dis 82:555–559
Lohnes DG, Schmitthenner AF, Nickell CD (1994) Incidence and distribution of Phytophthora resistance in new soybean germplasm from China. Agronomy Abstracts 217
Lohnes DG, Nickell CD, Schmitthenner AF (1996) Origin of soybean alleles for Phytophthora resistance in China. Crop Sci 36:1689–1692
Michelmore RW, Paran I, Kesseli RV (1991) Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci USA 88:9828–9832
Mueller EH, Athow KL, Laviolette FA (1978) Inheritance of resistance to four physiologic races of Phytophthora megasperma var. sojae. Phytopathology 68:1318–1322
Ploper LD, Athow KL, Laviolette FA (1985) A new allele at Rps3 locus for resistance to Phytophthora megasperma f. sp. glycinea in soybean. Phytopathology 75:690–694
Rohlf F (2000) NTSYS-PC numerical taxonomy and multivariate analysis system ver 2.11L. Appl Biostat, New York
Sandhu D, Schallock KG, Rivera-Velez N, Lundeen P, Cianzio S, Bhattacharyya MK (2005) Soybean Phytophthora resistance gene Rps8 maps closely to the Rps3 region. J Hered 96:536–541
Schmitthenner AF (1999) Phytophthora rot of soybean. In: Hartman GL, Sinclair JB, Rupe JC (eds) Compendium of soybean diseases, 4th edn. The American Phytopathological Society Press, St Paul, pp 39–42
Schmitthenner AF, Hobe M, Bhat RG (1994) Phytophthora sojae races in Ohio over a 10-year interval. Plant Dis 78:269–276
Schmutz J, Cannon SB, Schlueter J, Ma J, Mitros T, Nelson W, Hyten DL, Song Q, Thelen JJ, Cheng J, Xu D, Hellsten U, May GD, Yu Y, Sakurai T, Umezawa T, Bhattacharyya MK, Sandhu D, Valliyodan B, Lindquist E, Peto M, Grant D, Shu S, Goodstein D, Barry K, Futrell-Griggs M, Ab-ernathy B, Du J, Tian Z, Zhu L, Gill N, Joshi T, Libault M, Sethuraman A, Zhang XC, Shinozaki K, Nguyen HT, Wing RA, Cregan PB, Specht J, Grimwood J, Rokhsar D, Stacey G, Shoemaker RC, Jackson SA (2010) Genome sequence of the palaeopolyploid soybean. Nature 463:178–183
Shen CY, Su YC (1991) Discovery and preliminary studies of Phytophthora megasperma on soybean in China. Acta Phytopathol Sinica 21:298
Slaminko TL, Bowen CR, Hartman GL (2010) Multi-Year evolution of commercial soybean cultivars for resistance to Phytophthora sojae. Plant Dis 94:368–371
Sugimoto T, Yoshida S, Watanabe K, Aino M, Kanto T, Maekawa K, Irie K (2008) Identification of SSR markers linked to the Phytophthora resistance gene Rps1-d in soybean. Plant Breed 127:154–159
Sugimoto T, Yoshida S, Kaga A, Hajika M, Watanabe K, Aino M, Tatsuda K, Yamamoto R, Matoh T, Walker DR, Biggs AR, Ishimoto M (2011) Genetic analysis and Identification of DNA markers linked to a novel Phytophthora sojae resistance gene in the Japanese soybean cultivar Waseshiroge. Euphytica 182:133–145
Sugimoto T, Kato M, Yoshida S, Matsumoto I, Kobayashi T, Kaga A, Hajika M, Yamamoto R, Watanabe K, Aino M, Matoh T, David R, Walker DR, Biggs AR, Ishimoto M (2012) Pathogenic diversity of Phytophthora sojae and breeding strategies to develop Phytophthora-resistant soybeans. Breed Sci 61:511–522
Sun S, Wu XL, Zhao JM, Wang YC, Tang QH, Yu DY, Gai JY, Xing H (2011) Characterization and mapping of RpsYu25, a novel resistance gene to Phytophthora sojae. Plant Breed 130:139–143
Tang QH, Cui LK, Li DL, Dai TT, Yin WX, Dong SM, Xing H, Zheng XB, Wang YC (2010) Resistance evaluation of soybean germplasm from Huanghuai Valley to Phytophthora Root Rot. Sci Agric Sinica 43:2246–2252
Tyler BM (2007) Phytophthora sojae: root rot pathogen of soybean and model oomycete. Mol Plant Pathol 8:1–8
Van Ooijen JW (2006) JoinMap 4 Software for the calculation of genetic linkage maps in experimental populations. Kyazma B.V., Wageningen
Weng C, Yu K, Anderson TR, Poysa V (2001) Mapping genes conferring resistance to Phytophthora root rot of soybean, Rps1a and Rps7. J Hered 92:442–446
Wrather JA, Stienstra WC, Koenning SR (2001) Soybean disease loss estimates for the United States from 1996 to 1998. Can J Plant Pathol 23:122–131
Xia CJ, Zhang JQ, Wang XM, Wu XF, Liu ZX, Zhu ZD (2011) Analyses of resistance genes to Phytophthora root rot in soybean germplasm. Chin J Oil Crop Sci 33:396–401
Yao HY, Wang XM, Wu XF, Xiao YN, Zhu ZD (2010) Molecular mapping of Phytophthora resistance gene in soybean cultivar Zaoshu18. J Plant Genet Resour 11:213–217
Yu AL, Xu PF, Wang JS, Zhang SZ, Wu JJ, Li WB, Chen WY, Li NH, Fan SJ, Wang X, Jiang LY (2010) Genetic analysis and SSR mapping of gene resistance to Phytophthora sojae race 1 in soybean cv Suinong 10. Chin J Oil Crop Sci 32:462–466
Zhang SZ, Xu PF, Wu JJ, Xue AG, Zhang JX, Li WB, Chen C, Chen WY, Lv HY (2010) Races of Phytophthora sojae and their virulences on soybean cultivars in Heilongjiang, China. Plant Dis 94:87–91
Zhu ZD, Wang HB, Wang XM, Chang RZ, Wu XF (2003) Distribution and virulence diversity of Phytophthora sojae in China. Sci Agric Sinica 36:793–799
Zhu ZD, Huo YL, Wang XM, Huang JB, Wu XF (2006) Screening for resistance sources to Phytophthora root rot in soybean. J Plant Genet Resour 7:24–30
Zhu ZD, Huo YL, Wang XM, Huang JB, Wu XF (2007) Molecular identification of a novel Phytophthora resistance gene in soybean. Acta Agron Sinica 33:154–157
Acknowledgments
The work was supported by the Special Fund for Agro-scientific Research in the Public Interest (3-20, 201303018) and the Program of Protection of Crop Germplasm Resources (NB2010-2130135-25-14) from the Ministry of Agriculture of the People’s Republic of China. We thank Dr. Schmitthenner at the Ohio State University, USA, for providing the cultivars/lines carrying a single Rps gene at the Rps1 to Rps8 loci and the susceptible control cultivar Williams.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by D. Lightfoot.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Zhang, J., Xia, C., Wang, X. et al. Genetic characterization and fine mapping of the novel Phytophthora resistance gene in a Chinese soybean cultivar. Theor Appl Genet 126, 1555–1561 (2013). https://doi.org/10.1007/s00122-013-2073-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00122-013-2073-1