
Abstract
A population of 131 recombinant inbred lines from a wide cross between chickpea (Cicer arietinum L., resistant parent) and Cicer reticulatum (susceptible parent) segregating for the closely linked resistances against Fusarium oxysporum f.sp. ciceri races 4 and 5 was used to develop DNA amplification fingerprinting markers linked to both resistance loci. Bulked segregant analysis revealed 19 new markers on linkage group 2 of the genetic map on which the resistance genes are located. Closest linkage (2.0 cM) was observed between marker R-2609-1 and the race 4 resistance locus. Seven other markers flanked this locus in a range from 4.1 to 9.0 cM. These are the most closely linked markers available for this locus up to date. The sequences of the linked markers were highly similar to genes encoding proteins involved in plant pathogen response, such as a PR-5 thaumatin-like protein and an important regulator of the phytoalexin pathway, anthranilate N-hydroxycinnamoyl-benzoyltransferase. Others showed significant alignments to genes encoding housekeeping enzymes such as the MutS2 DNA-mismatch repair protein. In the Arabidopsis genome, similar genes are located on short segments of chromosome 1 and 5, respectively, suggesting synteny between the fusarium resistance gene cluster of chickpea and the corresponding regions in the Arabidopsis genome. Three marker sequences were similar to retrotransposon-derived and/or satellite DNA sequences. The markers developed here provide a starting point for physical mapping and map-based cloning of the fusarium resistance genes and exploration of synteny in this highly interesting region of the chickpea genome.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Chickpea (Cicer arietinum L.), one of the most important grain legume crops in the world, is grown extensively throughout most of the Indian subcontinent, in North Africa, West Asia and the Mediterranean region. Despite its economic importance, the productivity of the crop has not significantly been improved by conventional breeding. Additionally, the annual growth rate of chickpea production has been very slow (1.9%), and yields have risen at a rate of only 0.6% annually (FAO 1994). The main obstacle to increase yield is the crop's susceptibility to two fungal pathogens, Fusarium oxysporum Schlecht. emend Snyd. & Hans. f.sp. ciceri (Padwick) Snyd. & Hans. and Ascochyta rabiei (Pass.) Lab. Fusarium wilt is reported from many Asian, African, Mediterranean and South American countries, with yield losses ranging from 10 to 90% (Jiménez-Diaz et al. 1989; Ratnaparkhe et al. 1998). Different pathogenic races of F. oxysporum f.sp. ciceri have been identified. Races 1, 2, 3 and 4 were reported from India (Haware and Nene 1982), and races 0, 5 and 6 from Spain, respectively (Haware and Nene 1982; Jiménez-Diaz et al. 1989). Previous results for the mapping population used in the present study suggested that the resistance genes for Fusarium races 4 and 5 (foc-4 and -5) are located close to each other on the same linkage group (Simon and Muehlbauer 1997; Ratnaparkhe et al. 1998). Moreover, evidence derived from a narrow cross indicated clustering of resistance genes for Fusarium races 1 and 2 near the foc-4 locus (Simon and Muehlbauer 1997).
Chickpea breeding aims at combining resistances against different races of the fungus with tolerance against abiotic stresses like drought and cold in agronomically acceptable cultivars. Molecular markers could speed up the development of such cultivars by allowing the accumulation of resistance genes against different pathogen races by marker-assisted selection (MAS, reviewed by Winter and Kahl 1995). However, this approach requires the tagging of resistance genes by closely linked markers. Different marker types such as isozymes, RAPDs, AFLPs, SCARs, ISSRs and RGAs (Gaur and Slinkard 1990; Kazan et al. 1993; Labdi et al. 1996; Mayer et al. 1997; Ratnaparkhe et al. 1998; Hüttel et al. 1999), as well as sequence-tagged microsatellite sites (STMS, Beckmann and Soller 1990), and integrated marker maps are available for chickpea (Simon and Muehlbauer 1997; Hüttel et al. 1999; Winter et al. 1999, 2000). However, the marker density on linkage group 2, where the foc-4 and -5 resistance loci reside, is still too low for map-based cloning of the underlying resistance genes, even in the most advanced map of the chickpea genome (Winter et al. 2000). Here we report the increase of marker density around the fusarium resistance loci using bulked segregant analysis (BSA, Michelmore et al. 1991) and DNA amplification fingerprinting (DAF, Caetano-Anollés et al. 1991). DAF is comparable to the random amplified polymorphic DNA technique (RAPD, Welsh and McClelland 1990), but employs less DNA (0.1 to 1 ng/μl), and much higher primer concentrations (Caetano-Anollés et al. 1991). In our hands, DAF was superior to RAPD, since banding patterns were highly reproducible and polymorphic, despite the generally low level of polymorphism in the chickpea genome.
Materials and methods
Plant material and DNA extraction
The mapping population of 131 F7–F8 recombinant inbred lines (RILs) from a wide cross between chickpea accession ICC-4958 (resistant parent) and Cicer reticulatum accession PI 489777 (susceptible parent) has been described (Hüttel et al. 1999; Winter et al. 1999, 2000). Scoring for resistance to fusarium wilt in the parents and the RILs was extensively explained by Ratnaparkhe et al. (1998). Plants were propagated in the greenhouse. DNA was isolated from young leaflets using a modified cetyl-trimethyl-amoniumbromide (CTAB) protocol (Weising et al. 1995). Contaminating polysaccharides were selectively precipitated (Michaels et al. 1994). DNA concentrations were determined electrophoretically using known amounts of phage λ-DNA as a reference.
Bulked segregant analysis, BSA
For BSA, 1 ng of each of the DNA of 12 individuals that were either resistant (R14, R18, R22, R29, R53, R56, R72, R74, R87, R88, R94, R96) or susceptible (S11, S25, S32, S37, S40, S49, S55, S61, S63, S64, S65, S77) to Fusarium races 4 and 5 were mixed and used as a template.
DNA amplification fingerprinting and electrophoresis
DAF followed the procedure of Caetano-Anollés et al. (1991) with the following modifications: PCR was carried out on a Perkin Elmer Geneamp 9700 thermal cycler using random 10-mer primers procured from Eurogentec (Cologne, Germany), Operon Technologies (Alameda, USA) or Roth (Karlsruhe, Germany), respectively. Each 15 μl PCR reaction contained 1.5 μl of 10 × PCR buffer, 2.5 mM of MgCl2; 10 mM of dNTP-mix; 0.4 U of "Silverstar" DNA polymerase (Eurogentec), 40 pmol of oligonucleotide primer and 1 ng/μl of template DNA. The DNA was first denatured for 2 min at 95 °C, followed by 40 cycles of 15 s denaturation at 95 °C, 1 min annealing at 35 °C and 2 min elongation at 72 °C, with a final elongation at the same temperature for 2 min. The reaction products were separated on 1.8% agarose gels stained with ethidium bromide and visualized by UV light.
Linkage analysis
Marker order was determined using MAPMAKER V 3.0, kindly provided by Prof. Lander (Lander et al. 1987). Markers were placed on the map relative to the already mapped STMS and other markers (Hüttel et al. 1999; Winter et al. 1999, 2000) employing the "try" routine of the program. Final marker order was confirmed by multipoint analysis provided by MAPMAKER's "ripple" function using default parameters. Map distances were calculated using MAPMAKER's "Kosambi" option (Kosambi 1944).
Isolation, cloning and sequencing of linked DNA markers
Polymorphic marker fragments were isolated directly from agarose gels using the Qiaquick Kit (Qiagen, Cologne, Germany). The isolated fragments were ligated into the pGEM-T vector (Promega, Mannheim, Germany). Ligation products were transformed into electrocompetent Escherichia coli DH10-B, DH5-α (Gibco-BRL) or SURE (Stratagene) cells by electroporation (Gene-Pulser, BioRad, Munich, Germany). White colonies were transferred to liquid culture medium. The most frequent insert types were determined by restriction with endonucleases HinfI and AluI and their restriction fragments compared to those from the isolated DNA. Plasmids with restriction patterns similar to the original fragments were isolated using a modified alkaline-lysis procedure (Sambrook et al. 1989), sequenced in both directions using a dye terminator cycle sequencing kit (Perkin Elmer) and separated on an Applied Biosystems 373A. Sequences were translated into protein and screened for homology to entries in GenBank (National Center of Biotechnology Information, http://www.ncbi.nlm.nih.gov ) using BLASTN or BLASTX (Altschul et al. 1990, 1997).
Design of primers for sequence-specific amplified region (SCAR) markers
SCAR primer design was assisted by program Primer 3 (Rozen and Skaletsky 1998). Primers were 18–23 nt long (see Table 2), with annealing temperatures ranging from 62 to 65 °C, and expected product lengths from 314 to 1,103 bp. PCR was performed in 50-μl volumes in an Eppendorf Gradient Mastercycler. Each reaction contained 12 ng of DNA, 5 pMol/μl of each primer, 1.5 mM of MgCl2, 200 μM of each dNTP, 5.0 μl of 10 × PCR buffer and 0.5 U of "Silverstar" DNA polymerase (Eurogentec). The PCR reaction involved an initial denaturation step (94 °C for 2 min), followed by 40 cycles of denaturation at 94 °C for 15 s, annealing at 62, 64 or 65 °C for 30 s and elongation at 72 °C for 30 s, followed by a final elongation of 2 min at the same temperature. The SCAR PCR products from parental lines (accession ICC-4959, C. arietinum, and PI 489777, C. reticulatum), and three resistant (R-18, R-22, R-53) and susceptible (S-37, S-40, S-49) RILs, respectively, were separated on 1.8% agarose gels in 0.5 × TBE buffer and stained with ethidium bromide.
Results
Screening for polymorphic DAF markers using BSA and selective genotyping
In the first screening cycle 432 different 10-mers were tested on the bulks. Of these, 174 detected at least one amplification product that was polymorphic between the bulks, 242 detected no polymorphism, and 16 did not give rise to any amplification product. The 174 informative primers were then tested by selective genotyping of the two parental lines and seven resistant and seven susceptible RILs, respectively. Of these, 24 primers detected clear-cut polymorphic bands, that were either present or absent in all susceptible or resistant individuals, respectively (Fig. 1). Only these were selected for segregation analysis of the whole population. Nineteen of them were located on the same linkage group as the resistance genes (Table 1), and eight were tightly linked to foc-4 (Fig. 2).

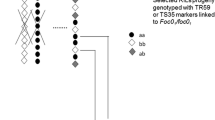
Representative picture of DAF products amplified with primer OP-M20 (upper part) apparently displaying two different alleles (black and white arrow) in both resistant (OP-M20-2) and susceptible (OP-M20-3) lines and a third faint band of the same size as band OP-M20-2 in the susceptible lines (see text for more detail). Lower part: marker OP-Q11 with a polymorphic band only in the resistant lines (arrow). ICC 4958: accession number of resistant parent; Resistant: resistant RILs; Susceptible: susceptible RILs; PI 489777: accession number of susceptible parent. First lane: 100-bp ladder

High-density map surrounding Fusarium resistance loci foc-4 and foc-5: comparison of marker order in chickpea LG 2 to the distribution of similar sequences on a short segment of Arabidopsis chromosome 5. Genetic distances between markers on LG 2 are indicated in cM, physical distances on the Arabidopsis chromosome are indicated in mega-base-pairs (Mbp). Bacterial artificial chromosome (BAC) clones from Arabidopsis containing the respective sequences are described by Sato et al. (1997) and Kaneko et al. (1999). The arrangement of clones is published in Arabidopsis Genome Initiative (2000). For more detail see text and Table 2
Marker order in relation to the foc-4 and -5 resistance genes
The 19 markers located on the same linkage group as resistance loci foc-4 and -5 are listed in Table 1. Fourteen markers displayed a polymorphic band in only the resistant (seven markers) or the susceptible (other seven marker) lines. Five primers revealed two different bands co-segregating in both resistant and susceptible lines. Figure 1 shows the banding patterns generated by primer OP-M20, with two apparently co-segregating alleles (Fig. 1, OP-M20-2 and -3), and OP-Q11-1, with a single polymorphic band in the resistant lines (Fig. 1B). Note the faint band of the same size as band OP-M20-2 in the susceptible RILs. The distribution of the markers in relation to resistance loci foc-4 and -5 on linkage group 2 is presented in Fig. 2. The markers most closely linked to foc-4 are R-2609-1 and OP-U17-1, flanking the locus on either side at map distances of 2.0 and 4.1 cM, respectively. Chi-square analysis revealed that 10 out of 19 loci deviated significantly (P = 0.05) from the expected Mendelian segregation ratio of 1:1. Clustering of distorted loci was most pronounced in the neighborhood of foc-4, which also showed distorted segregation (Table 1, Fig. 2).
Sequences of DAF bands closely linked to the foc-4 locus
To provide single-copy markers for marker-assisted selection and map-based cloning of the foc-4 gene, the amplification products were separated on agarose gels. The PCR products differentiating resistant and susceptible phenotypes were excised from complex DAF banding patterns, cloned and sequenced. Two bands amplified with primers OP-M20 and OP-P15 from both parental lines segregated like alleles of the same locus. In this case all four bands (OP-M20-2, OP-M20-3, OP-P15-3 and OP-P15-4) were cloned and sequenced. Therefore, the eight most tightly linked markers (see Fig. 2 and Table 1) were sequenced.
After a screen for homologies to entries in GenBank using BLASTN and BLASTX (Altschul et al. 1990, 1997), five out of eight cloned sequences were significantly similar to previously registered coding sequences (Table 2). Only markers OP-P08-1 (314 bp) and OP-P15-4 (492 bp) showed no significant similarity to GenBank entries. Interestingly, neither were homologies detected between the sequences of OP-M20-2 and OP-M20-3 (Fig. 1A), nor between OP-P15-3 and OP-P15-4, which initially were suspected to be alleles of one locus as judged from their segregation behavior.
Generation of SCARs and amplification of genomic DNA using SCAR primers
For the eight cloned DAF products, SCAR primer pairs were designed (Table 3). PCR with six primer pairs generated unique amplification products. However, in three cases non-reproducible, faint bands of lower molecular weight were also observed. SCAR primer pair OP-P06-4 produced a strong band of the expected size and a nearly similar band of higher molecular weight. From primer pair SCAR R-2609-1 only a stronger band superimposed on a smeary background could be obtained even under highly stringent annealing temperatures (64 °C). As expected, the strongest amplification products were polymorphic between the parental lines. Two primer pairs, SCAR OP-M20-3 and SCAR OP-P15-4, generated clear-cut, polymorphic bands between the parents and also between the two different classes of RILs (data not shown), whereas the others produced monomorphic bands only.
Discussion
Fine map around foc-4 and foc-5 loci
Recently, mapping of the chickpea genome advanced considerably due to the availability of STMS markers (Hüttel et al. 1999; Winter et al. 1999), and the integration of different marker types into a single map (Winter et al. 2000). Nevertheless, the tagging of agronomically important genes (as for example resistance genes for A. rabiei and F. oxysporum) is lagging far behind the progress made in other crops. None of the presently available markers were located close enough to either locus to allow map-based cloning of the respective genes. In fact, when the present work started, only 31 markers were available for linkage group 2 (LG-2), covering approximately 298 cM. The addition of DAF markers reported here did not only increase marker density specifically around the foc-4 locus, but expanded the size of LG-2 to nearly 380 cM by connecting hitherto unlinked markers to the LG-2 main body.
The use of BSA (Michelmore et al. 1991) for the targeted mapping of foc-4 and foc-5 on LG-2 was very efficient, since in a very short time and only with the DAF technique, the number of markers increased by 63% in this linkage group, and by 30% in the region harboring resistance genes. However, the distribution of markers on LG-2 is not uniform. A cluster of eight markers are closely linked to the foc-4 resistance locus, whereas only two markers segregated with the foc-5 locus, though at considerable distance. This is surprising, because the bulks contained individuals that were either resistant or susceptible, respectively, to both races of the pathogen. Therefore, both loci should have been hit with similar frequencies. It is possible that linkage of markers to the foc-5 locus reflects the much higher recombination frequencies around this locus as compared to the foc-4 locus. Markers therefore are apparently located further away. This assumption is corroborated by the observation that segregation distortion around the foc-4 locus not only affected the locus itself, but also the surrounding markers. In contrast, the foc-5 locus segregates normally. This would indeed explain why the apparent density of all markers is much higher around the foc-4 locus, as in many cases the whole region is transmitted to the offspring from the wild parent's genome. Segregation distortion is quite common in wide crosses and reflects inhomogeneities of parental genomes (Paran et al. 1995; Winter et al. 1999, 2000). Therefore, the map distance of 3.5 cM between the foc-5 locus and STMS marker TA27 (Winter et al. 2000) might indicate a smaller physical distance to the respective resistance gene than the distance of 2.0 cM between R-2609-1 and the foc-4 locus (Fig. 2). Another explanation for the discrepancies between marker densities around the two resistance loci could be that the region surrounding the foc-4 locus is more polymorphic than the one around the foc-5 locus, and therefore more markers are found there.
Comparison of cloned marker sequences to GenBank entries
Five out of eight sequences showed significant similarities to sequences coding for proteins (summarized in Table 2). This is astonishing, because the chickpea genome contains large amounts of repetitive DNA (Staginnus et al. 1999). Using random primers one would expect to preferentially hit these repeats. Indeed, three out of eight marker sequences contained repetitive elements. For example, the most tightly linked marker R-2609-1 was similar to a Ty3-gypsy-like retrotransposon (e-value 1e–124) and to members of the CaSat2 satellite DNA family (e-value 1e–66), both recently characterized in chickpea (Staginnus et al. 1999). The sequence of the marker OP-P06-4, 5.6 cM apart from foc-4 at the other flank of the locus, was similar to a retrotransposon-related gag-protease polyprotein gene and the retrovirus-like element SIRE-1 from soybean. The descent of these markers from repetitive elements is also mirrored by problems to define robust SCAR markers from them. There is no evidence yet that retrotransposons are involved in stress response in chickpea. However, several marker sequences (for accession numbers see Table 2) displayed striking similarities to factors associated with plant response to pathogen attack. For example, OP-U17-1, located between foc-4 (4.1 cM) and OP-P06-4 (1.5 cM), showed significant similarity (BLAST-X, 5e–10) to a thaumatin-like protein precursor (TLP) of Prunus avium and other plants, including Arabidopsis. This sweet-tasting protein belongs to the PR-5 family of antifungal proteins (Malehorn et al. 1994; Sassa and Hirano 1998). Several members of the PR-5-protein group of tobacco and other plants significantly inhibited spore germination or hyphal growth of various fungi, probably by permeabilizing fungal membranes (Stintzi et al. 1993; Sassa and Hirano 1998; Shih et al. 2001). PR-5-proteins also act as antifreeze-glycoproteins inhibiting growth under extreme cold. Southern hybridization data indicated that these proteins are encoded by single-copy genes (Shih et al. 2001).
The two adjacent markers OP-P15-3 and OP-P15-4 (co-segregating in the analyzed population and appearing as two alleles of the same locus) were of very different sequence composition. While OP-P15-4 possessed an anonymous AT-rich sequence, OP-P15-3, after translation to a peptide sequence, was in part significantly similar to the anthranilate N-hydroxycinnamoyl/benzoyltransferase protein from Arabidopsis thaliana chromosome 5 (e-value 6e–41). A shorter stretch of the chickpea sequence was nearly identical to the LRR-containing, N-like putative resistance protein from Arabidopsis and tobacco.
In tobacco and other solanaceous plants the synthesis of hydroxycinnamic acid amides of tyramide is induced after pathogen attack, wounding and ozone exposure. These amides are present in high concentrations in tobacco root tissues. They are found associated with cell-wall fractions and probably form a phenolic barrier protecting the cell walls against enzymatic degradation (Clarke 1982; Negrel and Javelle 1997; Farmer et al. 1999). In carnation (Dianthus caryophyllus L.), anthranilate N-hydroxy-cinnamoyl/benzoyltransferase catalyzes the first reaction of phytoalexin biosynthesis, and the product N-benzoylanthranilate is the precursor of several dianthramide phytoalexins, accumulating after infection with F. oxysporum f.sp. dianthi or Phytophtora parasitica (Yang et al. 1997). Phytoalexins dramatically increase in concentration after fungal infection (Grayer and Kokubun 2001). Moreover, a positive correlation between phytoalexin accumulation and fusarium wilt resistance exists in chickpea. Stevenson et al. (1997) infected four chickpea cultivars with F. oxysporum f.sp. ciceri races 1 and 2. The roots of susceptible and resistant cultivars were penetrated, but hyphal growth in resistant accessions was very slow. The infection was accompanied by a remarkable accumulation of phytoalexins in all cultivars. Altogether, these evidences suggest a correlation between marker OP-P15-3 and the resistance to fusarium wilt race 4 in chickpea.
Whereas the AT-rich sequence of locus OP-P08-1, following markers OP-P15-3/OP-P15-4 at a distance of 1 cM had no homology to known sequences, the translated sequences of markers OP-M20-2 and OP-M20-3 displayed considerable similarity to proteins involved in housekeeping functions such as DNA repair or chromatin condensation. Similar to the marker pair OP-P15-3/OP-P15-4, the OP-M20-derived markers also segregated as alleles of the same locus. However, whereas the sequence of marker OP-M20-2 was similar to a MutS2-like mismatch repair protein gene from A. thaliana located on chromosome 5 (e-value 3e–13), marker OP-M20-3 was highly similar (2e–167) to a putative NBS-LRR-encoding gene and the predicted chromosome condensation regulator protein of A. thaliana, also on chromosome 5 (Kaneko et al. 1999), and thus both sequences are not allelic.
The translated sequence of marker OP-M20-3 is significantly similar to the putative chromosome condensation factor from Arabidopsis and Triticum monococcum (1e–140), and to the group of HERC2 genes from Drosophila and human. These highly conserved proteins are involved in chromosome condensation and regulation of chromosome rearrangements (Ji et al. 2000). Like the MutS-gene, the gene underlying OP-M20-3 may also be associated with cell division and growth.
It is noteworthy, that most marker sequences were significantly similar to sequences located on chromosome 5 (sometimes also chromosome 1) of A. thaliana. The presence of similar sequences on both chromosomes is not surprising, since the Arabidopsis genome harbors blocks of duplicated sequences on different or even the same chromosome (The Arabidopsis Genome Initiative 2000). At least three of these sequences (similar to markers OP-M20-3, OP-P15-3 and OP-P06-4) are located in close vicinity in a segment spanning 5 Mbp on Arabidopsis chromosome 5 (Kaneko et al. 1999). Adjacent to this region, sequences similar to OP-M20-2 and OP-U17-1 are located (Sato et al. 1997; Kaneko et al. 1999). If the similarities are not by chance, the order of these sequences in chickpea and Arabidopsis is virtually the same. Such colinearity or conserved synteny in chromosomes of plants from distantly related taxa is evident for the entire length of soybean linkage group 2 and Arabidopsis chromosome 1 (Grant et al. 2000), for a 105-kbp segment of tomato chromosome 2 and Arabidopsis chromosomes 2 and 5 (Ku et al. 2000), and also for the sequence of a 57-kbp fragment of tomato chromosome 7 and Arabidopsis chromosome 1 (Rossberg et al. 2001). Lee et al. (2001) compared genomic segments in soybean to the homologous regions in the related Phaseolus vulgaris and Vigna radiata. Not only were linkage blocks between the legume genomes conserved, but also between the legumes and Arabidopsis, which diverged from each other some 90 million years ago. Thus, our results fit into the emerging picture of genome evolution in plants of the Arabidopsis – tomato clade including legumes as a result of repeated rounds of genome duplication followed by gene loss in one or the other species (Ku et al. 2000; Eckardt 2001). The potential synteny between segments of chickpea LG-2 and chromosomes 1 and 5 of Arabidopsis requires further research.
Our results allow the assumption that, additionally to the cluster of fusarium resistance genes (foc-1, foc-2, foc-4, foc-5; Mayer et al. 1997; Ratnaparkhe et al. 1998; Tullu et al. 1998; Winter et al. 2000; review in Winter et al. 2002), pathogenesis-related genes are also clustered in the same region of linkage group 2 of the genetic map of chickpea. Co-localization of defense-response genes with disease resistance loci has also been observed in pepper (Pflieger et al. 2001). In A. thaliana, genes recognizing pathogens and responding to pathogen attack are unevenly distributed among the chromosomes. Most of these genes reside on chromosome 1 (49) and chromosome 5 (55 genes), respectively, and a few only on other chromosomes (The Arabidopsis Genome Initiative 2000). Co-localization of genes involved in different aspects of defense against pathogens possibly is a more general phenomenon than expected.
To gain deeper insight into chickpea's defense against pathogens it is, therefore, justified to focus on the region surrounding the fusarium resistance gene cluster and eventually sequence it. A BAC library for chickpea has been generated already (Hüttel et al., in preparation), and a physical map and BAC-contigs starting from the SCARs reported here are in preparation. This will not only allow to clone the fusarium resistance genes, but also to clarify whether or not this region is indeed syntenic to Arabidopsis chromosome 1 and 5, and to orthologous genomic regions in other plants.
References
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215:403–410
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25:3389–3402
Beckmann JS, Soller M (1990) Towards a unified approach to genetic mapping of eucaryotes based on sequence tagged microsatellite sites. Bio/Technology 8:930–932
Caetano-Anollés G, Bassam BJ, Gresshoff PM (1991) DNA amplification fingerprinting: a strategy for genome analysis. Plant Mol Biol Rep 9:292–305
Clarke DD (1982) The accumulation of cinnamic acid amides in the cell walls of potato tissues as an early response to fungal attack. In: Wood KRS (ed) Active defense mechanisms in plants. Plenum Press, New York, pp 321–322
Eckardt NA (2001) Everything in its place: conservation of gene order among distantly related plant species. Plant Cell 13:723–725
Farmer MJ, Czernic P, Michael A, Negrel J (1999) Identification and characterization of cDNA clones encoding hydroxycinnamoyl-CoA:tyramine N-hydroxycinnamoyltransferase from tobacco. Eur J Biochem 263:686–694
Food and Agriculture Organization of the United Nations (1994) FAO production year book. FAO. Rome
Gaur PM, Slinkard AE (1990) Inheritance and linkage of isozyme-coding genes in chickpea. J Hered 81:455–461
Grant D, Cregan P, Shoemaker RC (2000) Genome organization in dicots: genome duplication in Arabidopsis and synteny between soybean and Arabidopsis. Proc Natl Acad Sci USA 97:4168–4173
Grayer RJ, Kokubun T (2001) Plant-fungal interactions: the search for phytoalexins and other antifungal compounds from higher plants. Phytochemistry 56:253–261
Haware MP, Nene YL (1982) Races of Fusarium oxysporum f. sp. ciceri. Plant Dis 66:809–810
Hüttel B, Winter P, Weising K, Choumane W, Weigand F, Kahl G (1999) Sequence-tagged microsatellite site markers for chickpea (Cicer arietinum L.). Genome 42:210–217
Ji Y, Rebert NA, Joslin JM, Higgins MJ, Schultz RA, Nicholls RD (2000) Structure of the highly conserved HERC2 gene and of multiple partially duplicated paralogs in human. Genetic Res 10:319–329
Jiménez-Diaz RM, Trapero-Casas AT, Cabrera de la Colina J (1989) Races of Fusarium oxysporum f.sp. ciceri infecting chickpea in southern Spain. In: Tjamos EC, Beckman EH (eds) Vascular wilt diseases of plants. Springer, Berlin Heidelberg New York, vol H28
Kaneko T, Katoh T, Sato S, Nakamura Y, Asamizu E, Kotani H, Miyajima N, Tabata S (1999) Structural analysis of Arabidopsis thaliana chromosome 5. IX. Sequence of the regions of 1,011,550 bp covered by 17 TAC clones. DNA Res 6:183–195
Kazan K, Muehlbauer FJ, Weeden NF, Ladizinsky G (1993) Inheritance and linkage relationships of morphological and isozyme loci in chickpea (Cicer arietinum L.). Theor Appl Genet 86:417–426
Kosambi DD (1944) The estimation of map distances from recombination values. Ann Eugen 12:172–175
Ku H-M, Vision T, Liu J, Tanksley SD (2000) Comparing sequenced segments of the tomato and Arabidopsis genomes: large-scale duplication following selective gene loss creates a network of synteny. Proc Natl Acad Sci USA 97:9121–9126
Labdi M, Robertson LD, Singh KB, Charrier A (1996) Genetic diversity and phylogenetic relationships among the annual Cicer species as revealed by isozyme polymorphisms. Euphytica 88:181–188
Lander E, Green P, Abrahamson J, Barlow A, Daly M, Lincoln S, Newburg L (1987) MAPMAKER: an interactive computer package for constructing primary linkage maps of experimental and natural populations. Genomics 1:174–181
Lee JM, Grant D, Vallejos CE, Shoemaker RC (2001) Genome organization in dicots. II. Arabidopsis as a 'bridging species' to resolve genome evolution events among legumes. Theor Appl Genet 103:765–773
Malehorn DE, Borgmeyer JR, Smith CE, Shah DM (1994) Characterization and expression of an antifungal zeamatin-like protein (Zlp) gene from Zea mays. Plant Physiol 106:1471–1481
Mayer MS, Tullu A, Simon CJ, Kumar J, Kraft JM, Muehlbauer FJ (1997) Development of a DNA marker for fusarium wilt resistance in chickpea. Crop Sci 37:1625–1629
Michaels SD, John MC, Amasino RM (1994) Removal of polysaccharides from plant DNA by ethanol precipitation. Biotechniques 17:274–276
Michelmore RW, Paran I, Kesseli RV (1991) Identification of markers linked to disease resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions using segregating populations. Proc Natl Acad Sci USA 88:9828–9832
Negrel J, Javelle F (1997) Purification, characterization and partial amino-acid sequencing of hydroxycinnamoyl-CoA:tyramine N-(hydroxycinnamoyl)transferase from tobacco cell suspension cultures. Eur J Biochem 247:1127–1135
Paran I, Goldmann I, Tanksley SD, Zamir D (1995) Recombinant inbred lines for genetic mapping in tomato. Theor Appl Genet 90:542–548
Pflieger S, Palloix A, Caranta C, Blattes A, Lefebvre V (2001) Defense response genes co-localize with quantitative disease resistance loci in pepper. Theor Appl Genet 103:920–929
Ratnaparkhe MP, Santra DK, Tullu A, Muehlbauer FJ (1998) Inheritance of inter-simple-sequence-repeat polymorphisms and linkage with a Fusarium wilt resistance gene in chickpea. Theor Appl Genet 96:348–353
Rossberg M, Theres K, Acarkan A, Herrero R, Schmitt, T, Schumacher K, Schmitz G, Schmidt R (2001) Comparative sequence analysis reveals extensive microcolinearity in the lateral suppressor regions of the tomato, Arabidopsis and Capsella genomes. Plant Cell 13:979–988
Rozen S, Skaletsky HJ (1998) Primer 3. http://www-genome.wi.mit.edu/genome_software/ other/ primer3.html
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning: a laboratory manual, 2nd edn. Cold Spring Harbor Laboratory Press, New York
Sassa H, Hirano H (1998) Style-specific developmentally regulated accumulation of a glycosylated thaumatin/PR-5-like protein in Japanese pear (Pyrus serotina Rehd.). Planta 205:514–521
Sato S, Kotani H, Nakamura Y, Kaneko T, Asamizu E, Fukami M, Miyayima N, Tabata S (1997) Analysis of Arabidopsis thaliana chromosome 5. I. Sequence features of the 1.6-Mb regions covered by 20 physically assigned P1 clones. DNA Res 4:215–230
Shih C-YT, Wu J, Jia S, Khan AA, Ting K-LH, Shih DS (2001) Purification of an osmotin-like protein from the seeds of Menincasa hispida and cloning of the gene encoding this protein. Plant Sci 160:817–826
Simon CJ, Muehlbauer FJ (1997) Construction of a chickpea linkage map and its comparison with maps of pea and lentil. J Hered 88:115–119
Staginnus C, Winter P, Desel C, Schmidt T, Kahl G (1999) Molecular structure and chromosomal localization of major repetitive DNA families in the chickpea (Cicer arietinum L.) genome. Plant Mol Biol 39:1037–1050
Stevenson PC, Turner, HC, Haware MP (1997) Phytoalexin accumulation in the roots of chickpea (Cicer arietinum L.) seedlings associated with resistance to fusarium wilt (Fusarium oxysporum f. sp. ciceri). Physiol Mol Plant Pathol 50:167–178
Stintzi A, Heitz T, Prasad V, Wiedemann-Merdinoglu S, Kaufmann S, Geoffroy P, Legrand M, Fritig B (1993) Plant 'pathogenesis-related' proteins and their role in defense against pathogens. Biochimie 75:687–706
The Arabidopsis Genome Initiative (2000) Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408:796–815
Tullu A, Muehlbauer FJ, Simon CJ, Mayer MS, Kumar J, Kaiser WJ, Kraft JM (1998) Inheritance and linkage of a gene for resistance to race 4 of fusarium wilt and RAPD markers in chickpea. Euphytica 102:227–232
Weising K, Nybom H, Wolff K, Meyer W (1995) DNA fingerprinting in plants and fungi. CRC Press, Boca Raton, Florida
Welsh J, McClelland M (1990) Fingerprinting genomes using PCR with arbitrary primers. Nucleic Acids Res 18:7213–7218
Winter P, Kahl G (1995) Molecular marker technologies for crop improvement. World J Microbiol Biotechnol 11:449–460
Winter P, Pfaff T, Udupa SM, Hüttel B, Sharma PC, Sahi S, Arreguin-Espinoza R, Weigand F, Muehlbauer FJ, Kahl G (1999) Characterization and mapping of sequence-tagged microsatellite sites in the chickpea (Cicer arietinum L.) genome. Mol Gen Genet 262:90–101
Winter P, Benko-Iseppon AM, Hüttel B, Ratnaparkhe M, Tullu A, Sonnante G, Pfaff T, Tekeoglu M, Santra D, Sant VJ, Rajesh PN, Kahl G, Muehlbauer FJ (2000) A linkage map of the chickpea (Cicer arietinum L.) genome based on recombinant inbred lines from a C. arietinum × C. reticulatum cross: localization of resistance genes for fusarium wilt races 4 and 5. Theor Appl Genet 101:1155–1163
Winter P, Staginnus C, Sharma PC, Kahl G (2002) Organization and genetic mapping of the chickpea genome. In: Jaiwal PK, Singh RP (eds) Focus on biotechnology vol 10A, improvement strategies of Leguminosae biotechnology. Kluwer Academic Publishers, The Netherlands, pp 327–380
Yang Q, Reinhard K, Schiltz E, Matern U (1997) Characterization and heterologous expression of hydroxycinnamoyl/benzoyl-CoA:anthranilate N-hydroxycinnamoyl-benzoyl-transferase from elicited cell cultures of carnation, Dianthus caryophyllus L. Plant Mol Biol 35:777–789
Acknowledgements
The authors thank T. Pfaff, J. Geistlinger, S. Rakshit and S. Kost, Plant Molecular Biology, Biocentre, for valuable suggestions and technical support. The present research was supported by Bundesministerium für Technische Zusammenarbeit (BMZ, Germany, grant no. 2001.7860.8-001.00), Deutsche Forschungsgemeinschaft (DFG grant Ka332–17/2-3), Fundação Coordenação de Aperfeiçoamento de Pessoal de Nível superior (CAPES, Brazil, grant no. BEX-0505/98-6) and FACEPE (Fundação de Amparo à Pesquisa do Estado de Pernambuco). Molecular marker sequences were registered in GenBank: accession nos. AF457587, AF457588, AF457593, AF457589, AF457590, AF457591, AF457585 and AF457586.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by C. Möllers
Rights and permissions
About this article
Cite this article
Benko-Iseppon, AM., Winter, P., Huettel, B. et al. Molecular markers closely linked to fusarium resistance genes in chickpea show significant alignments to pathogenesis-related genes located on Arabidopsis chromosomes 1 and 5. Theor Appl Genet 107, 379–386 (2003). https://doi.org/10.1007/s00122-003-1260-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00122-003-1260-x