
Abstract
Biodiversity–ecosystem functioning experiments (BEF) typically manipulate sown species richness and composition of experimental communities to study ecosystem functioning as a response to changes in diversity. If sown species richness is taken as a measure of diversity and aboveground biomass production as a measure of community functioning, then this relationship is usually found to be positive. The sown species richness can be considered the equivalent of a local species pool in natural communities. However, in addition to species richness, realized diversity is also an important community diversity component. Realized diversity is affected by environmental filtering and biotic interactions operating within a community. As both sown species richness and the realized diversity in BEF studies (as well as local species pool vs observed realized richness in natural communities) can differ markedly, so can their effects on the community functioning. We tested this assumption using two data sets: data from a short-term pot experiment and data from the long-term Jena biodiversity plot experiment. We considered three possible predictors of community functioning (aboveground biomass production): sown species richness, realized diversity (defined as inverse of Simpson dominance index), and survivor species richness. Sown species richness affected biomass production positively in all cases. Realized diversity as well as survivor species richness had positive effects on biomass in approximately half of cases. When realized diversity or survivor species richness was tested together with sown species richness, their partial effects were none or negative. Our results suggest that we can expect positive diversity–productivity relationship when the local species pool size is the decisive factor determining realized observed diversity; in other cases, the shape of the diversity–functioning relationship may be quite opposite.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Species diversity is often considered to be a driving factor of ecosystem functioning. This idea is mostly based on “biodiversity–ecosystem functioning experiments” (BEF), which have demonstrated that in sown mixtures of randomly selected sets of species from a common pool, the ecosystem functioning (often represented by aboveground biomass production) increases with the number of sown species (Balvanera et al. 2006; Hooper et al. 2012; Naeem et al. 2012; but see Kenkel et al. 2000). To explain the mechanisms driving this positive relationship, two mutually nonexclusive hypotheses were suggested: the complementarity and selection (sampling) effects (Loreau and Hector 2001). The complementarity effect hypothesis states that multiple functionally of different species should be able to use the available niche space better and thus increase their biomass production. The selection effect is more or less a probabilistic phenomenon produced by the BEF studies experimental setting: with increasing number of species in mixture, the chance that a species with a dominant role in community functioning will be included also increases.
It is important to note that a variety of community biodiversity measures have been proposed in addition to species richness; species evenness and functional trait composition have been recognized as important factors shaping community functioning (Diaz and Cabido 2001; Garnier et al. 2004; Sanderson 2010; Sasaki and Lauernroth 2011; Schmitz et al. 2013). In BEF studies, the sown species richness (mixture size) is in fact equivalent to a local species pool in natural communities, as defined by Butaye et al. (2001). Following their concept, the local species pool can be defined as a pool of species which have reached a given locality and are able to grow under given environmental conditions. The observed local community is then composed of the local species pool minus species filtered out by several possible factors. Among the most important factors are interspecific interactions, mainly competition, but these effects are also modified by facilitation and multi-trophic interactions. Another factor which may influence the observed diversity in natural grasslands, as well as in field experiments using permanent plots, is small scale environmental heterogeneity. This has been shown to increase the number of locally coexisting species and also to change the magnitude and relative importance of the biodiversity effects operating within a community (Wacker et al. 2008; Richardson et al. 2012). All these factors influence not only the presence of individual species but also their relative proportions of biomass, and ultimately the diversity. The observed species richness and observed species proportions characterizing realized diversity are thus important characteristics of a community.
The diversity–productivity relationship is not only dependent on the diversity measure used but also on time and spatial scale studied (Chalcraft et al. 2004; Weiss et al. 2007). Long-term BEF experiments allow for species interactions to occur and possible competitive exclusion of some species or immigration of others, which affects the observed species richness. This can then be higher/lower than the sown one. The phenomenon of possible species extinctions is of great interest because the main aim of BEF studies has always been to predict consequences of such events in nature (Chapin et al. 2000). Some multi-seasonal BEF experiments were maintained by weeding while other let colonizers invade experimental plots. It has already been demonstrated that the positive relationship between observed species richness and biomass disappears quickly if the plots are open to new invasions (Pfisterer et al. 2004; Lepš et al. 2007; Rixen et al. 2008; Roscher et al. 2009; Petermann et al. 2010).
The shapes of diversity–productivity relationship differing from the positive one widely found in BEF experiments are often reported from observational studies located in temperate managed grasslands. These are the most similar communities to those in plant BEF studies, and the relationship between observed local species richness and biomass production is often negative in these places or not monotonous (Thompson et al. 2005; Lepš 2013; Mittelbach et al. 2001). In such communities, the species richness is limited mostly by environmental filtering and competitive exclusion of weak competitors.
In our study, we addressed the possibility of a differential relationship between sown species richness, two measures of realized diversity and biomass production. The sown species richness is the only manipulated variable in BEF studies, and therefore seems to be the natural explanatory variable. However, the species already excluded from a community can hardly participate in niche partitioning, resulting in the complementarity effect. Similarly, the potential for complementarity will decrease with decreasing evenness of species (Nijs and Roy 2000). This justifies using realized diversity as an explanatory variable in the study of the diversity–productivity relationship. Concerning the mechanisms behind the diversity–productivity relationship, there is also a question of causality. In BEF experiments, the causal direction is clear: sown richness affects biomass production. In contrast, in natural conditions, the community diversity is affected by both species pool and by processes within the community, making direction of causality unclear (Lepš 2013; Stachová and Lepš 2010). It is known that the positive biodiversity–productivity relationship is not often found in nature (Schmid 2002). In natural conditions, we are usually only able to observe the realized species richness and diversity and not the sown diversity, which corresponds to the community local species pool (i.e., to the species that were able to arrive to the site). And so, the community species pool size is generally unknown, and although the estimation methods do exist, they are far from being precise enough to enable any comparison with BEF experiments. For this reason, the only way to compare the diversity–productivity relationships found in experiments with those in nature is using the realized diversity.
Consequently, we have asked which of the three possible predictors (sown species richness, survivor species richness, and realized diversity, defined as inverse of Simpson dominance index) predicts best the community productivity as a sole predictor (what are their marginal effects). We have also asked how the use of characteristics of realized diversity changes the predictions based on the sown diversity (what are their partial effects after accounting for the sown species richness).
We based our hypotheses on the mechanisms operating within the community. It is clear that the number of sown species is a determinant of selection effect: more species available (i.e., sown) leads to a higher chance of getting highly productive species in the mixture. Following this reasoning, there should be an explicit positive relationship between sown species richness and biomass production; and as the sown species richness is a defined set of species at the beginning and does not change during an experiment, this positive relationship should be constant over time.
On the other hand, complementarity should be mainly dependent on the species actually present in the community and on their proportions. So, there should be a tight positive relationship between the observed species richness (survivor species richness and realized diversity) and biomass production. We can expect this relationship to strengthen with the length of an experiment, as it has been shown in multi-seasonal BEF experiments that the complementarity effect size increases with time. This is not surprising because in all the theories explaining maintenance of species diversity, niche differentiation (and so the resource that use complementarity) is one of the basic prerequisites of species coexistence (Wilson 2011). Therefore, we can expect that species surviving in the experimental plots are those able to use the resources in a complementary way.
In our analyses, we also focus on the effect of realized diversity and survivor species richness in a constant local species pool (fixed mixture size in BEF studies). Our model (details in Stachová and Lepš 2010) predicted a nonexistent or negative relationship between survivor species richness and biomass production in communities resulting from a species pool of constant size.
We examined two data sets: data from a seasonal greenhouse pot experiment using six plant species (Špaekova and Lepš 2001) and a publicly available data set from a 6-year period of the Jena experiment, one of the largest and longest BEF experiment ever (Weigelt et al. 2010). The short-term experiment is a typical example of a widespread BEF experiment lasting only one season. In such experiments, time for competitive exclusion to occur is relatively short, and thus, the realized and sown species richness is usually quite similar. The communities were established in pots where the potential for even minor environmental heterogeneity is limited. In the long-term experiment, the diversities, both realized and sown, had enough time to diverge allowing us to test the above hypotheses and theoretical predictions.
Material and methods
Glasshouse experiment
For this experiment, six naturally co-occurring meadow species from two functional groups were planted in pots (16 cm in diameter, 14 cm high). All possible species combinations within all species-richness level were used. After 5 months, aboveground biomass was assessed and sorted into individual species. For details on this experiment, see Špaekova and Lepš (2001).
Jena experiment
The data set retrieved from Weigelt et al. (2010) was based on sampling of permanent plots 20 × 20 m. The species pool of this experiment consists of sixty species and the following species richness levels: monocultures, two, four, eight, 16, and 60 species mixtures. The plots were harvested twice a year before mowing (usually in May and August). We used data from six consecutive years (2003–2008), from the May harvest as well as the August harvest. Every plot consisted of three or four subplots 0.2 × 0.5 m, and the exact coordinates of each subplot were randomly generated within the main plot before every harvest. This is exactly the spatial scale, at which the individuals are expected to interact with each other; thus, we analyzed the subplots as separate observations (for details and publications based on previous analyses of this data set, see Weigelt et al. (2010)).
Data analysis
The diversity of plant communities of each pot/subplot was described by reciprocal of Simpson’s index of dominance (Lepš 2013): \( D={\displaystyle {\sum}_i\;{\left(\frac{N_i}{N}\right)}^2} \), where N i is the biomass of ith species and N is total biomass of a community. Note that in this form, 1/D is scaled as the number of species and sometimes is called the equivalent number of species, because it is the number of species reaching the same diversity being equally represented in the community. We refer to the sown number of species “sown species richness” and 1/D as “realized diversity.” The “survivor species richness” was calculated as the number of species yielding more than 5 % of the total pot/subsample biomass. This approach was used because in the pot experiment, there was no sufficient time for a species to become extinct, and we wanted to keep a unified methodology for both compared data sets. For the Jena data set, we have also used as survivor species richness the number of all species present in a subsample (total survivor species richness) (results are presented in the Supplementary material). In our linear regressions, aboveground biomass was considered as a response, with realized diversity, sown species richness and survivor species richness used as predictors. Sown species richness and the survivor species richness were log-transformed in all analyses of the Jena experiment data set. First, the marginal effects of all predictors were evaluated (all predictors were used in three separate linear regressions for all years in the Jena experiment), and in the second step, we used sown species richness as a first predictor and realized diversity or survivor species richness as a second predictor to get their partial effect on biomass production, again for all years in/of the Jena experiment. The partial effects show the effect of each of the predictors in addition to the first predictor, i.e., to the sown species richness. It can be also understood as the effects of the predictor if the sown species richness is kept constant. In the Jena experiment (which has enough various combinations of species composition at the same sown richness), we visualized this effect by inspecting the relationship between realized diversity and biomass for subsets of the same sown richness.
Results
In the glasshouse experiment, marginal effects of all predictors were significantly positive, and partial effects of realized diversity or survivor species richness were always nonsignificant (Tables 1 and 2).
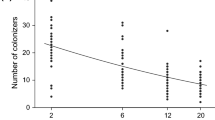
In the Jena experiment, marginal effects of sown species richness were significantly positive in all harvests and all seasons while marginal effects of both realized diversity and survivor species richness were significantly positive in around half of cases (six and seven out of 11 for realized diversity and survivor species richness, respectively) (Table 1). In a linear model using the predictors sown species richness and realized diversity, the partial effect of realized diversity was significantly negative in eight cases. Similar results (seven significantly negative cases) were detected for the partial effects of survivor species richness, again with sown species richness as the first predictor (for summarized statistical results for partial effects, see Table 2). The same analyses (marginal and partial effects) performed using total survivor species richness from the Jena experiment again yielded rather similar results (see Supplementary material). The negative effects of realized species richness and survivor species richness at a fixed sown species richness level are displayed in Fig. 1.

Jena experiment: relationships between biomass production in 2007 and realized diversity or survivor species richness for mixtures. Regression lines in bold denote significant relationships, and dashed lines denote nonsignificant ones
Discussion
The diversity of a natural community is determined by the available species pool, abiotic filtering, and biotic interactions within the community (Reitalu et al. 2008; Myers and Harms 2009; Götzenberger et al. 2012; Lepš 2013). Our data support the frequently found positive dependence of productivity on the sown species richness (Cardinale et al. 2011; Naeem et al. 2012), which can be translated into a positive relationship between the local species pool and productivity in nature. In our view, all the species used in a BEF experiment can be understood as a regional (“total” sensu Butaye et al. 2001) species pool, i.e., all the species available in the geographical area. The set of sown species in individual plots corresponds to the local species pool, i.e., set of the species able to reach the site, and all the other species are artificially prevented from entering the community (either by not sowing or by weeding if they were able to reach the site naturally). This in fact corresponds to the dispersal limitation of these species in nature. It is obvious that our analogy is somewhat limited. In the Jena experiment, the size of the plots is 20 × 20 m. At this spatial scale, species are able to recolonize subplots from which they had disappeared much faster than they are able to recolonize localities at the landscape level. Also, species present in a given plot but absent from a subplot may affect biomass in a subplot through indirect effects on other trophic levels or through belowground processes. These indirect effects of extinct species are negligible at the landscape level, although even here the species once present can affect the present community (e.g., through soil herbivores and pathogens; Kostenko et al. 2012).
Sown species richness in our analyses showed positive marginal effects on biomass production in all seasons and harvests studied in the Jena experiment and also in the glasshouse experiment. Realized diversity and survivor species richness are derived from and highly correlated with (ranging between 0.45 and 0.7 in the Jena experiment) sown species richness; so, their marginal effects were positive in half of cases in the Jena experiment and in the glasshouse experiment. Sown species richness had the strongest explanatory power of all the predictors. Nevertheless, the R 2 of all the positive relationships were rather low (see Table 1). Observation of a temporally stable positive relationship between sown species richness and biomass production is in-line with our hypothesis concerning the strong mechanism of selection effect. The actual values of selection effect for the Jena experiment have been assessed by Marquard et al. (2009) and reported as decreasing over time. However, it has been debated whether or not the phenomenon of increasing importance of (statistically determined) complementarity effect and the corresponding decreasing importance of selection effect over time might be caused solely by deteriorating performance of monocultures. The monoculture values enter the formulas for biodiversity effects as a “reference,” and it is very possible that the monoculture production diminished due to accumulation of pathogens and imbalanced depletion of resources (e.g., Marquard et al. 2013). Taking a different point of view, accumulation of pathogens can be seen as a valid proximate mechanism behind the complementarity effect. In our analyses of the Jena experiment, we used data on number and biomass proportions of species present in a subplot (as these data were available). However, it is important to note that the species missing in subplots might have not been missing from the whole plot. Thus, these rare species were probably available for recolonization and also might have provided potential for some mechanisms considered as complementarity effect (e.g., via support of populations of herbivores, mutualists, pathogens, see Fornara and Tilman 2009; de Kroon et al. 2012; Reich et al. 2012). For example, Lepš et al. (1998) have demonstrated that even a species with as low biomass as 0.27 g. m−2 in Alder Carr was able to support rather large population of monophagous moth. Considering these facts, even species absent from the basic sampling unit can, to some limited extent, contribute to the complementarity effect.
The weak marginal and sometimes negative partial effects of realized diversity and survivor species richness on biomass production which we observed in the Jena experiment are very probably caused by the fact that our measure of community functioning is aboveground biomass. When sown species diversity turns into realized diversity, species which are likely to be lost from the realized community are those that are rare, i.e., have low biomass. Such species by definition do not contribute much to the biomass production, and their loss might be followed by biomass increase as the dominant species take over. However, these rare species might be important for different measures of community functioning (e.g., Jain et al. 2014) than biomass production.
There seemed to be no directed temporal pattern for the relationship between biomass production and realized diversity measures in the Jena experiment. In the first year, there was a positive relationship between both measures of realized diversity and biomass production, which was not present in two subsequent years. However, between the fourth and sixth years, these relationships were positive again. The experiments which have found no relationship between realized diversity and community functioning usually allowed for immigration of new species resulting in a survivor species richness higher than the sown one (at least in smaller mixture sizes). Plots in the Jena experiment were regularly weeded, and so, the survivor species richness was always lower than the sown one in the data set we used. In our model (Stachová and Lepš 2010) which used different sown species richness and allowed for competitive exclusion only, the relationship between survivor species richness and biomass production was positively saturated since the positive slope was mainly caused by the lowest richness level. Our results on realized diversity and survivor species richness affecting biomass production in either positive or no way partially contradicts our hypothesis of increasing positive relationship with time due to increasing importance of complementarity effect. The analysis of the Jena experiment by Marquard et al. (2009) showed that the net and complementarity effects slightly decreased in 5 years and after reaching a minimum in the third year began increasing in subsequent years. Correspondingly, a similar temporal trend of the average survivor species richness and average realized diversity was pronounced in our analyses (Fig. S1). This means that the average complementarity and net effects were higher when more species survived and/or average species evenness in mixtures was higher. The fact that the relationship between measures of realized diversity and biomass production was not always positive and was generally rather weak is caused by a mechanism counteracting the positive complementarity effect. This mechanism is linked with the nature of the commonly used measure of community functioning and biomass production. If a community contains highly productive species, their biomass production will suppress the other species, which can manifest in decreased evenness or even survival and thus shape the realized diversity of the community. As we can see, the causality here is quite opposite: The functioning affects realized diversity. The two mechanisms (the complementarity and a negative effect of dominant species on community diversity) are operating simultaneously and can either balance out each other or one of them prevails. If we screen out the positive effects of species pool, the effects of realized diversity/survivor species richness is negative or none (partial effects). This was also apparent in constant sown species richness (mixture levels) conditions (Fig. 1), very similarly to the predictions of Stachová and Lepš (2010). Marquard et al. (2009) found that increased biomass production between diversity levels in Jena was caused by increased plant density, but within a mixture level, it was caused by increased plant size. This is in concordance with the negative effect of realized diversity at the mixture (sown diversity) level as plots consisting larger individuals would presumably have lower realized diversity.
We may also generalize the mechanism responsible for no or negative effects of realized community diversity metrics for the whole data set: Plants compete mainly for light, and as this competition is asymmetric, the presence of a highly productive species in a mixture not only increases its productivity but also increases the suppression (or even extinction) of other species. Absence of such a species not only leads to lower productivity but also increases the chance that none of the species will be outcompeted, particularly when the species use the resources in a complementary way.
Another possible control over the expression of sown richness and realized diversity would have been environmental filtering affecting species sorting. However, it is obvious that the species for the Jena experiment were selected on the basis of a deep field experience to fit the habitat conditions; the fact that the monocultures perform fairly well over long-term in the site (Marquard et al. 2013) confirms this view.
To conclude, if the diversity in diversity–productivity relationship in natural conditions is mostly determined by variation in species pool, then, this relationship will be positive and the causality is clear: The more species present, the more biomass they produce. However, we should be aware that in nature, the species pool seldom differs among geographically close communities, as it is varied in BEF studies. If the size of species pool does not differ among communities, both the realized diversity and the productivity depend mostly on the traits of constituent species (and environmental characteristics which are usually kept constant in BEF). The realized diversity is then often negatively correlated with (or independent of) biomass production. This hypothesis was supported by a model (Stachová and Lepš 2010) which simulated composition of communities from a common pool: The more species a simulated community in equilibrium had, the more they were complementary (had lower competition coefficients). On the other hand, species in poor communities consisted of highly productive species gaining dominance and increasing total community productivity. As a result, the model predicted no or negative diversity—productivity relationships depending on the species pool size.
There is a direct causal relationship between the size of available species pool and community productivity. Both the realized diversity and the productivity are affected by a series of factors (including species pool size and composition, environmental factors, and also determining biotic interactions). If the size of species pool is the decisive factor determining realized diversity, then, we can expect positive diversity–productivity relationship. In other cases, the shape of the relationship will be highly variable.
The loss of species in Central European grasslands is most often connected with increased dominance of highly productive species (Lepš 2013), meaning, it does not lead to decreased productivity. However, our results suggest that multiple local extinctions of various species will lead to a considerable decrease of regional and local species pool size, which might result in negative effects on ecosystem functioning in future.
References
Balvanera P, Pfisterer AB, Buchmann N, He JS, Nakashizuka T, Raffaelli D, Schmid B (2006) Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecol Lett 9:1146–1156
Butaye J, Jacquemyn H, Honnay O, Hermy M (2001) The species pool concept applied to forests in a fragmented landscape: dispersal limitation versus habitat limitation. J Veg Sci 13:27–34
Cardinale BJ, Matulich KL, Hooper DU, Byrnes JE, Duffy E, Gamfeldt L, Balvanera P, O’Connor MI, Gonzales A (2011) The functional role of producer diversity in ecosystems. Am J Bot 98:572–592
Chalcraft DR, Williams JW, Smith MD, Willig MR (2004) Scale dependence in the species richness-productivity relationship: the role of species turnover. Ecology 85:2701–2708
Chapin FS, Zavaleta ES, Eviner VT et al (2000) Consequences of changing biodiversity. Nature 405:234–242
Diaz S, Cabido M (2001) Vive la difference: plant functional diversity matters to ecosystem processes. Trends Ecol Evol 16:646–655
Fornara DA, Tilman D (2009) Ecological mechanisms associated with the positive diversity-productivity relationship in an N-limited grassland. Ecology 90:408–418
Garnier E, Cortez J, Billes G, Navas M et al (2004) Plant functional markers capture ecosystem properties during secondary succession. Ecology 85:2630–2637
Götzenberger L, de Bello F, Bråthen KA et al (2012) Ecological assembly rules in plant communities—approaches, patterns and prospects. Biol Rev Camb Philos 87:111–127
Hooper DU, Adair EC, Cardinale BJ et al (2012) A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 486:105–109
Jain M, Flynn DFB, Prager CM, Hart GM, DeVan CM, Ahrestani FS et al (2014) The importance of rare species: a trait-based assessment of rare species contributions to functional diversity and possible ecosystem function in tall-grass prairies. Ecol Evol 4:104–112
Kenkel NC, Peltzer DA, Baluta D, Pirie D (2000) Increasing plant diversity does not influence productivity: empirical evidence and potential mechanisms. Commun Ecol 1:165–170
Kostenko O, van de Voorde TFJ, Mulder PPJ, van der Putten WH, Bezemer TM (2012) Legacy effects of aboveground–belowground interactions. Ecol Lett 15:813–821
de Kroon H, Hendriks M, van Ruijven J, Ravenek J, Padilla FM, Jongejans E et al (2012) Root responses to nutrients and soil biota: drivers of species coexistence and ecosystem productivity. J Ecol 100:6–15
Lepš J (2013) Diversity and ecosystem function. In: van der Maarel E, Franklin J (eds) Vegetation ecology, 2nd edn. John Wiley, Oxford, pp 308–346
Lepš J, Spitzer K, Jaroš J (1998) Food plants, species composition and variability of the moth community in undisturbed forest. Oikos 81:538–548
Lepš J, Doležal J, Bezemer TM et al (2007) Long-term effectiveness of sowing high and low diversity seed mixtures to enhance plant community development on ex-arable fields in five European countries. Appl Veg Sci 10:97–110
Loreau M, Hector A (2001) Partitioning selection and complementarity in biodiversity experiments. Nature 412:72–76
Marquard E, Weigelt A, Temperton VM et al (2009) Plant species richness and functional composition drive over yielding in a six-year grassland experiment. Ecology 90:3290–3302
Marquard E, Schmid B, Roscher C et al (2013) Changes in the abundance of grassland species in monocultures versus mixtures and their relation to biodiversity effects. PLoS One 8(9):e75599
Mittelbach GG, Steiner CF, Scheiner SM et al (2001) What is the observed relationship between species richness and productivity? Ecology 82:2381–2396
Myers JA, Harms KE (2009) Seed arrival, ecological filters, and plant species richness: a meta-analysis. Ecol Lett 12:1250–1260
Naeem S, Duffy JE, Zavaleta E (2012) The functions of biological diversity in an age of extinctions. Science 336:1401–1406
Nijs I, Roy J (2000) How important are species richness, species evenness and interspecific differences to productivity? Oikos 88:57–66
Petermann J, Fergus A, Roscher C et al (2010) Biology, chance or history? The predictable re-assembly of temperate grassland communities. Ecology 91:408–421
Pfisterer AB, Joshi J, Schmid B et al (2004) Rapid decay of diversity-productivity relationships after invasion of experimental plant communities. Basic Appl Ecol 5:5–14
Reich PB, Tilman D, Isbell F, Mueller K, Hobbie SE, Flynn DFB, Eisenhauer N (2012) Impacts of biodiversity loss escalate through time as redundancy fades. Science 336:589–592
Reitalu T, Prentice HC, Sykes MT et al (2008) Plant species segregation on different spatial scales in semi-natural grasslands. J Veg Sci 19:407–416
Richardson PJ, MacDougall AS, Larson DW (2012) Fine-scale spatial heterogeneity and incoming seed diversity additively determine plant establishment. J Ecol 100:939–949
Rixen C, Huovinen C, Huovinen K et al (2008) A plant diversity×water chemistry experiment in subalpine grassland. Perspect Plant Ecol 10:51–61
Roscher C, Temperton VM, Buchmann N, Schulze ED (2009) Community assembly and biomass production in regularly and never weeded experimental grasslands. Acta Oecol 35:206–217
Sanderson MA (2010) Stability of production and plant species diversity in managed grasslands: a retrospective study. Basic Appl Ecol 11:216–224
Sasaki T, Lauernroth WK (2011) Dominant species, rather than diversity, regulates temporal stability of plant communities. Oecologia 166:761–768
Schmid B (2002) The species richness-productivity controversy. Trends Ecol Evol 17:113–114
Schmitz M, Flynn DFB, Mwangi PN et al (2013) Consistent effects of biodiversity on ecosystem functioning under varying density and evenness. Folia Geobot 4:335–353
Špaekova I, Lepš J (2001) Procedure for separating the selection effect from other effects in diversity-productivity relationship. Ecol Lett 4:585–594
Stachová T, Lepš J (2010) Species pool size and realized species richness affect productivity differently: a modelling study. Acta Oecol 36:578–586
Thompson K, Askew AP, Grime JP et al (2005) Biodiversity, ecosystem function and plant traits in mature and immature plant communities. Funct Ecol 19:355–358
Wacker L, Baudois O, Eichenberger-Glinz S, Schmid B (2008) Environmental heterogeneity increases complementarity in experimental grassland communities. Basic Appl Ecol 9:467–474
Weigelt A, Marquard E, Temperton VM et al (2010) The Jena experiment: six years of data from a grassland biodiversity experiment. Ecology 91:929
Weiss JJ, Cardinale BJ, Forshay KJ, Ives AR (2007) Effects of species diversity on community biomass production change over the course of succession. Ecology 88:929–939
Wilson JB (2011) The twelve theories of co-existence in plant communities: the doubtful, the important and the unexplored. J Veg Sci 22:184–195
Acknowledgments
We would like to thank Paul J. Richardson and two anonymous referees for constructive comments on our manuscript. We also thank Sirja Seppa and Genevieve Finerty for the language revision. The research was supported by GAČR-13-17118S.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by: Sven Thatje
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 120 kb)
Rights and permissions
About this article

Cite this article
Rychtecká, T., Lanta, V., Weiterová, I. et al. Sown species richness and realized diversity can influence functioning of plant communities differently. Naturwissenschaften 101, 637–644 (2014). https://doi.org/10.1007/s00114-014-1198-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00114-014-1198-7