
Abstract
Arboreal ants form patchy spatial patterns in tropical agroforest canopies. Such patchy distributions more likely occur in disturbed habitats associated with lower ant diversity and resource availability than in forests. Yet, few studies have quantitatively examined these patchy patterns to statistically test if ants are non-randomly distributed or at what scale. Coffee agroecosystems form a gradient of management intensification along which vegetative complexity and ant diversity decline. Using field studies and a spatially explicit randomization model, I investigated ant patchiness in coffee agroecosystems in Chiapas, Mexico varying in management intensity to examine if: (1) coffee intensification affects occurrence of numerically dominant ants, (2) numerical dominants form statistically distinguishable single-species patches in coffee plants, (3) shade trees play a role in patch location, and (4) patch formation or size varies with management intensity. Coffee intensification correlated with lower occurrence frequency of numerically dominant species generally and of one of four taxa examined. All dominant ant species formed patches but only Azteca instabilis was patchy around shade trees. Ant patchiness did vary somewhat with spatial scale and with strata (within the coffee layer vs around shade trees). Patchiness, however, did not vary with management intensity. These results provide quantitative evidence that numerically dominant ants are patchy within the coffee layer at different scales and that shade tree location, but not coffee management intensity, may play a role in the formation of patchy distributions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In tropical agroforests, numerically dominant ants, those found in higher numbers, biomass, or frequency of occurrence, are often spatially arranged such that different species may form mutually exclusive patches where submissive ants coexist (Leston 1973; Room 1975; Majer 1978; Majer and Queiroz 1993; Davidson 1998). Descriptions of these patchy patterns, although abundant in the literature, are mostly qualitative; researchers generally use maps to show patchy distributions without quantifying spatial patterns (e.g., Leston 1973; Majer and Queiroz 1993). Many non-exclusive hypotheses explain formation of patchy distribution patterns including (1) interspecific competition (Leston 1973; Room 1975; Majer 1978; Adams 1994), (2) stochastic processes (Ribas and Schoereder 2002), (3) decreasing species richness (Hölldobler and Wilson 1990), (4) physical or temporal habitat specialization (Floren and Linsenmair 2000; Soares and Schoereder 2001), and (5) anthropogenic disturbances (Floren et al. 2001). Ants are more likely patchily distributed in disturbed habitats where they suffer more intense resource competition for food or nest sites (Floren and Linsenmair 2000; Floren et al. 2001). In this paper, I quantitatively test for patchy distributions of ants in the coffee plant layer of coffee agroecosystems to examine if these patterns are statistically distinguishable from random and if patchiness is related to shade tree locations or influenced by coffee management intensity.
Coffee agroecosystems fall along an intensification gradient from those cultivated under a native forest shade canopy at the least intensive end, towards intensive production characterized by lower density, diversity, and height of shade trees, lower canopy cover, higher coffee density, and agrochemical use (Moguel and Toledo 1999). Generally speaking, coffee intensification results in losses of ant biodiversity (Philpott and Armbrecht 2006), but this is not always the case (Lachaud and García-Ballinas 2001). Diversity losses are likely a result of microclimatic differences (Perfecto and Vandermeer 1996), reductions in number or diversity of nesting sites (Armbrecht et al. 2004), or increases in pesticide use. In shaded coffee agroecosystems, numerically dominant arboreal ants most often nest in shade trees and forage in coffee plants nearby (Vandermeer et al. 2002), only sometimes nesting in coffee plants. Although most studies examining the patchy distributions of ants refer to ant communities in canopy trees, it is likely that, where fewer trees are available as nest sites, patchy distributions may form in what is usually the understory—i.e., the coffee plant layer. Furthermore, given that patchy patterns are influenced by habitat disturbance and that trees play an important role in arboreal ant communities, coffee management intensification may affect distributions of patch-forming arboreal ants within the coffee agroecosystems.
In this study, I quantitatively tested for single-species patches of numerically dominant arboreal ants in the coffee plant layer (excluding the shade canopy) and examined whether shade tree locations or management intensification may affect either location of size of patches in coffee farms. I specifically asked (1) whether coffee intensification affects occurrence frequency of numerical dominants, (2) if numerical dominants form statistically distinguishable single-species patches in the coffee layer, (3) if shade tree location plays a role in the placement of patches, and (4) if the number of species forming patches or if size of patches varies with management intensity. The approach here is relatively new in that it uses a quantitative model to test for ant patchiness, thereby allowing us to statistically distinguish single-species patches in a landscape and also allowing quantitative testing of alternative hypotheses for formation of patchy distributions (but see Theunis et al. 2005, for a review).
Materials and methods
Site description
I carried out field studies in four farms in the Soconusco region of SW Chiapas, Mexico: (1) Finca Belen (15°15′ N, 92°22′ W), (2) Finca Irlanda (15°11′ N, 92°20′ W), (3) Finca Hamburgo (15°10′ N, 92°19′ W), and (4) Guadalupe Zajú (15°9′ N, 92°17′ W). All farms cover more than 200 ha and receive ca. 4,500 mm of rain per year, and all study plots within farms are located between 950 and 1,150 m. According to the coffee management scheme of Moguel and Toledo (1999) and in order of increasing intensification, Irlanda and Belen both contain areas of traditional polyculture and commercial polyculture, Hamburgo is a shaded monoculture, and Guadalupe Zajú is an unshaded sun plantation. Irlanda and Belen are both certified organic farms and no pesticides have been used in Hamburgo for at least 5 years. Guadalupe Zajú was also the only site where pesticides may have been used but this information was not made available to me. Coffee densities varied between sites but, in general, plants are separated by 0.5–3 m and sometimes are directly touching other coffee plants or shade trees. I haphazardly established a total of nine 25×50-m survey plots in the four farms to capture the range of management intensification. I set up three plots in traditional polycultures (TPB1, TPB2, and TPI), three in commercial polycultures (CPB1, CPB2, and CPI), two in shade monocultures (SM1 and SM2), and one in unshaded sun production (SUN). I established six of them in June 2000 (TPB1, TPI, CPB1, CPI, SM1, and SM2), two of them in January 2001 (TPB2 and CPB2), and the final one in June 2002 (SUN).
Vegetation and ant sampling
To verify the qualitative assessments of management intensity in each plot, I surveyed multiple vegetation characteristics and summarized these using a management index. I collected data on percent shade cover, shade tree density, tree species richness, number of trees >100 cm circumference at breast height, and coffee density. I estimated percent cover for all sites in January 2001, June 2001, January 2002, and June 2002 using a vertical tube densitometer. Every 5 m throughout plots, for a total of 50 points, I recorded 0, 50, or 100% cover and averaged all points for a per-plot value. I summarized all vegetation variables using a management index (modified from Mas and Dietsch 2003). Raw data for each variable (per plot and per date) were converted to a scale from 0 (least intensive) to 1 (most intensive). To convert all variables except coffee density, I divided each measurement by the highest possible value and subtracted by one. Increases in coffee density, unlike increases of the remaining vegetation variables, correspond to increasing management intensity. Thus, for coffee density, I divided by the lowest measurement and subtracted by one. I summed values for each variable for a possible total of five per plot and divided by the number of variables measured on each date.
I surveyed coffee plants and shade trees for foraging ants in most plots three times in the wet season (June 2000, 2001, and 2002) and twice in the dry season (January 2001 and 2002), depending on the date in which plots were established. All ants were sampled during the day, thus no nocturnally foraging ants were included in the study. In each plot, I first mapped x- and y-coordinates of coffee and shade trees. I sampled ants in each coffee plant by kicking or shaking plants and watching branches, leaves, and trunks for 2 min after disturbances. I sampled shade trees using tuna baits (5–10 g) placed on trunks 1 m aboveground and checked after 30–45 min. Coffee plants and shade trees were assigned to the following categories based on the presence of foraging workers: (1) Azteca instabilis (F. Smith), (2) Camponotus senex textor Forel, (3) Camponotus novogranadensis Mayr, (4) Crematogaster spp., (5) all other ants, and (6) no ants. Several species of Crematogaster are found within the study sites including C. carinata Mayr, C. crinosa Mayr, C. formosa Mayr, C. nigropilosa Mayr, and C. sumichrasti Mayr. Unfortunately, these were not distinguished at the time field surveys were conducted and thus are treated as a genus here. Other common arboreal ants found in the study sites include Pheidole spp., Nesomyrmex spp., Brachymyrmex spp., and Pseudomyrmex spp. A total of 81 species of ants have been sampled from coffee plants and shade trees in the study sites from 1999–2003. Extensive information about ant diversity and abundance in the same sites is summarized in Philpott et al. (2006). I included only A. instabilis, C. senex textor, C. novogranadensis, and Crematogaster spp. as numerical dominants based on the fact that they were frequently encountered in the shade trees in the study sites.
I used ANOVA and Tukey’s tests to verify differences in management intensity between all sites. For individual vegetation characteristics, I used a Bonferonni correction for multiple tests (α=0.0125); multivariate statistics were unpractical because not all variables were measured on all dates. Tree circumference and SUN were only sampled once and were not included in statistical comparisons. To test for relationships between the management index and ant occurrence on coffee plants or shade trees, I used multiple regressions. Because plots sampled on different dates do not constitute independent data points, I averaged management index values and ant occurrence frequency (proportion of total coffee plants or shade trees with ants) for use in regressions. I first tested for relationships between occurrence frequency on coffee plants and shade trees and the management index for all species, numerical dominants, and other ants combined. I then tested data for each species separately.
Quantifying ant patchiness in the coffee layer and around shade trees
To quantitatively test for ant patchiness within species, I developed two models, conceptually equivalent to two-tailed t tests, using Pascal (Turbo 5). With each model, I compared data for observed ant spatial distribution patterns (field data) to computer-generated, randomized spatial distribution patterns. Each model thus simultaneously evaluated if ants formed single-species patches or were distributed at random or in uniform patterns. The two models differed in that model I examined patchiness in the coffee layer (excluding the shade trees) and model II examined patchiness relative to the shade trees. Model II specifically tested the prediction that shade tree location determines patch placement within coffee agroecosystems. Models I and II separately analyzed data from each date and plot and for each numerically dominant species. For observed (i.e., field collected) data, the models counted the number of coffee neighbors that contained conspecifics relative to other coffee plants (model I) or shade trees (model II). The numbers of coffee neighbors with conspecifics were summed across all coffee plants or shade trees in a given plot for each species. The models repeated this process 1,000 times for computer-generated data sets where the model randomly assigned coffee plants to an ant category with the same frequency in which they were observed in the field. The model then compared observed data to the frequency distributions created to determine if each species was more likely than random to occur on neighbors (patchy), less likely to occur (uniform), or equally as likely to occur (randomly distributed). The outcome from the two models indicated for each plot and sampling date if each species was distributed in patches, at random, or uniformly in the coffee plant layer (model I) or in coffee plants around shade trees with the same ant species (model II). During each run of the model, every coffee plant (I) or shade tree (II) in the plot was used as a focal point around which neighbors were assessed. For each plot and date, I ran models four times using distinct neighbor distances (3, 5, and 10 m or the ten closest plants) to determine which scale, if any, best captured ant patchiness for each species. At times during field surveys, I found more than one numerical dominant species on a coffee plant or shade tree. However, because running models was highly time intensive (20 min per simulation for >300 simulations), I assigned plants to one species giving priority to A. instabilis, then C. senex textor, then Crematogaster spp., ultimately limiting the number of additional simulations needed. In some plots, one or more species were absent or infrequently encountered, thus models were programmed to run if an ant species was found on more than two plants per plot.
To determine overall patterns of species-specific patchiness and changes due to distance, strata, or management intensity, I compiled data for each plot across all dates. It was not surprising that ant distribution patterns did differ somewhat among dates for particular plots, but I was more interested in comparing plots under different management systems. For each plot, I counted on how many dates each species was distributed in patches, uniformly, or at random and converted these frequencies into percent of total observations for a total of nine data points per species. Overall patterns of patchiness for all plots, ants, distances, and strata (coffee and shade trees) were assessed qualitatively. To test if patchiness varied with distance or depending on whether the focal tree was a coffee plant or shade tree, I used MANOVA using the percent of trials that were patchy for each species as the dependent variables and distance (3, 5, and 10 m or the nearest ten plants) and strata (coffee or shade tree) as main factors. I also tested if patchiness varied with coffee management intensity. I used multiple regressions to examine if the percent of trials in which ants were patchy correlated with the management index. I used one test for each numerical dominant where percent of trials that were patchy at each distance and strata were treated as the dependent variables (eight in each regression) and management intensity as the independent variable.
Results
Effects of management intensity on ant occurrence
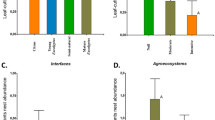
Management intensity differed significantly between sites as measured in terms of individual vegetation variables and the management index (Table 1). For all species, numerical dominants, and other ants combined, occurrence frequency was significantly correlated with increasing management intensity (R 2=0.715; F=17.587; df=1,7; P=0.004). With increasing intensity, occurrence frequency of one numerical dominant, but not of others, significantly declined (Fig. 1). On both coffee plants and shade trees, occurrence of A. instabilis declined with increasing intensity (R 2=0.485; F=6.59; df=1,7; P=0.037). The occurrence of other (not numerically dominant) ants increased with management intensification (R 2=0.448; F=5.68; df=1,7; P=0.048). Individual occurrence of C. senex textor, C. novogranadensis and Crematogaster spp. did not vary with management intensity.

Occurrence frequency of numerical dominants and other ants as a function of management intensity. Symbols show proportion of coffee plants (closed diamonds) and shade trees (open squares) with a A. instabilis, b C. senex textor, c C. novogranadensis, d Crematogaster spp., e all numerical dominant ants, and f other non-numerical dominants. The management index measured coffee management intensity (Table 1), and high values correspond to more intensive management. Dagger (†) indicates that occurrence decreased significantly with management index and double dagger (‡) indicates that occurrence increased significantly with management index (see text for details)
Effects of distance and strata on ant patchiness
All numerically dominant ant species sampled formed patches in the coffee layer and shade in at least some trials, but the proportion of trials in which ants were patchy differed by species identity, distance, and strata (Fig. 2). For all distances, A. instabilis was patchy in both the coffee layer (65–96% of trials) and around the shade trees (73–83% of trials). In roughly half of trials in the coffee layer, C. senex textor formed patches (48–59%), and this ant was patchy less often in the shade trees (21–47%). The proportion of trials in which C. novogranadensis was patchy ranged greatly with distance in the coffee layer (30–84%) as well as around shade trees (40–88%). Finally, Crematogaster spp. were patchy in 37–76% of trials in the coffee layer and in 25–40% of trials with shade trees. Overall, ant patchiness, however, did not differ consistently with distance from focal trees or with strata (Table 2). Nonetheless, between-subjects tests on a per-species basis showed that the percent of trials in which Crematogaster spp. were patchy was significantly higher for the coffee layer. Furthermore, C. novogranadensis was more likely to be patchy at the distance of 3 m than that at 10 m (Tukey’s test, P=0.024), but the percent of trials where this ant was patchy did not differ for other distances.

Numerical dominant ant patchiness in coffee agroecosystems as a factor of strata, distance, and species. Bars show mean percents of model trials and for which distances [3, 5, and 10 m, or the ten closest trees (10t)] for ant taxa (see Fig. 1) were distributed in patches, at random, or uniformly in the coffee plant layer (a) or around shade trees (b) in 25×50-m-plots species
Ant patchiness and management intensity
In general, the percent of trials in which ants were patchy did not correlate with changes in coffee management intensity. For none of the distances or strata included did any numerically dominant ant patchiness correlate with changes in management intensity: A. instabilis (R 2=0.018, F=0.0768, df=1, 4, P=0.7953), C. senex textor (R 2=0.2518, F=1.346, df=1, 4, P=0.3104), C. novogranadensis (R 2=0.006, F=0.027, df=1, 4, P=0.8779), and Crematogaster spp. (R 2=0.039, F=0.163, df=1, 4, P=0.706). Because multivariate tests did not show significant overall patterns, no tests for individual strata or distances were conducted.
Discussion
The proportion of coffee and shade trees with numerically dominant ants overall declined with coffee management intensification, but this pattern was entirely driven by declines of A. instabilis; the other numerically dominant species were not affected by intensification. All ant species formed statistically distinguishable single-taxon patches in at least some plots on some sample dates. Although the analyses carried out here do not allow detailed quantitative comparisons between species, the patterns observed suggest that A. instabilis was patchy in a higher proportion of trials both in the coffee layer and around shade trees than the other species were. Only A. instabilis appeared to consistently form patches around shade trees. C. senex textor was patchy in approximately half or a little fewer than half of trials. Patterns for C. novogranadensis showed much wider variation in the percent of trials where this ant was patchy than did other species. Statistical tests indicated that C. novogranadensis were significantly more patchy at shorter distances, such that they formed patches in a lower percentage of trials at 10 m than at 3 m. Finally, Crematogaster spp. were relatively rarely patchy compared to other species, especially around shade trees. Crematogaster spp. formed patches less frequently around shade trees than in the coffee layer. Patchiness of none of the ants, however, was affected by coffee management intensity, indicating that this anthropogenic disturbance may not influence formation of single-species patches.
Patchiness greatly varied between species, as A. instabilis was always patchy whereas other ant species were only patchy at some distances or only in the coffee plants and not in coffee plants around the shade trees. A. instabilis formed tight patches up to 10 m from coffee and shade trees in all management systems, even though A. instabilis abundance declined with intensification. The two patterns showing that A. instabilis occurrence declined with management intensification and that this species very frequently formed patches around shade trees may indicate that trees are a very important resource for this species and may in part determine its spatial distribution (i.e., patch location) within coffee agroecosystems but not patch formation. In particular, A. instabilis tend to nest in large trees within coffee farms (personal observation). Thus, intensifying production by removing shade trees would limit the number of plants per plot with this ant. C. senex textor is a nest-weaver (Forel 1899) and is more often seen nesting in the coffee plants (personal observation). Crematogaster spp. and C. novogranadensis nest opportunistically in cavities and dead and live branches (Longino 2002) and, thus, may be ubiquitous in all parts of the coffee farms and not restricted to shade trees. Because ants were sampled differently in the coffee plants and shade trees, caution must be used in interpreting the results, as methodological differences may have influenced the apparent results. Due also to the fact that Crematogaster spp. were treated here as a genus and not as single species as is the characteristic of studies of ant spatial distributions, all results involving Crematogaster must be treated with caution. Different species of Crematogaster may have different nesting habits or food requirements, which may make examining their spatial distributions as a genus flawed.
Understanding and quantifying which factors determine ant spatial structure is critical to understand ant impacts on arthropod communities. According to this study, patchy patterns in ant communities are not influenced by changes in management intensity or, aside from A. instabilis, by shade tree locations. This is true despite the fact that fewer food and nesting resources are available in intensively managed coffee plantations (Moguel and Toledo 1999; Armbrecht et al. 2004). In coffee plantations, most dominant ant species nest in the shade trees, although occasionally A. instabilis, C. senex textor, or Crematogaster spp. nests are found in the coffee bushes (personal observation). Thus, removal of shade trees would be expected to limit one important resource for dominant ants—nest sites. Furthermore, resources such as extrafloral nectar found on shade trees (such as Inga spp.) are important diet components of these ants (Davidson et al. 2003) as are homopterans and other arboreal arthropods. In the study sites, arboreal ant diversity declines with intensification (Philpott et al. 2006) and some have suggested that, where diversity is higher, it is unnecessary or difficult for one ant species to dominate a particular area or patch (Hölldobler and Wilson 1990). Furthermore, patchy distributions of ants are not found in many natural forests because of lower resource availability (e.g., Floren and Linsenmair 2000). My results, however, provide little evidence that management intensification within this particular gradient, lower ant diversity, or a lower supply of available resources relate to changes in ant spatial patterns.
Many alternative hypotheses – including many that have already been examined qualitatively – could be tested with the same types of spatially explicit quantitative techniques presented here. Nest site specialization or niche partitioning could be examined to test if ant distributions follow closely particular tree species (Djieto-Lordon and Dejean 1999). Ant distribution patterns have elucidated that particular dominant ants associate with homopterans or other arthropods (Leston 1973; Majer 1978; Majer et al. 1994; Dejean et al. 1997). Different resource preferences among ant species may also structure ant communities in patches. This same model could be used to examine relationships between ant patterns and such resources. Furthermore, such a model could be used to examine other types of physical or temporal habitat specialization among ant species or examine aspects of interspecific competition such as negative relationships among ant species.
In summary, most ant species were patchily distributed in at least some trials at some scales, upholding a quantitative test of single-species ant patchiness across a coffee landscape. Of all species examined, A. instabilis appeared to be the most often patchy and this patchiness was associated with shade trees. The model used here was useful to determine if ant patchiness differs with scale and management intensity and could also shed light on alternative hypotheses to explain patchy distributions of ants.
References
Adams ES (1994) Territory defense by the ant Azteca trigona—maintenance of an arboreal ant mosaic. Oecologia 97:202–208
Armbrecht I, Perfecto I, Vandermeer J (2004) Enigmatic biodiversity correlations: ant diversity responds to diverse resources. Science 304:284–286
Davidson D (1998) Resource discovery versus resource domination in ants: a functional mechanism for breaking the trade-off. Ecol Entomol 23:484–490
Davidson DW, Cook SC, Snelling R, Chua TH (2003) Explaining the abundance of ants in lowland tropical rainforest canopies. Science 300:969–972
Dejean A, Djieto-Lordon C, Durand JL (1997) Ant mosaic in oil palm plantations of the southwest province of Cameroon: impact on leaf miner beetle (Coleoptera: Chrysomelidae). J Econ Entomol 90:1092–1096
Djieto-Lordon C, Dejean A (1999) Tropical arboreal ant mosaics: innate attraction and imprinting determine nest site selection in dominant ants. Behav Ecol Sociobiol 45:219–225
Floren A, Linsenmair KE (2000) Do ant mosaics exist in pristine lowland rain forests? Oecologia 123:129–137
Floren A, Freking A, Biehl M, Linsenmair KE (2001) Anthropogenic disturbance changes the structure of arboreal tropical ant communities. Ecography 24:547–554
Forel A (1899) Biologia Centrali-Americana; or, contributions to the knowledge of the fauna and flora of Mexico and Central America. Insecta. Hymenoptera. 3 (Formicidae). London
Hölldobler B, Wilson EO (1990) The ants. Harvard University Press, Cambridge, Massachusetts
Lachaud JP, García-Ballinas JA (2001) Diversité de la myrmécofaune (Ponerinae et Cerapachyinae) dans les agrosystèmes de café et cacao au Mexique. Insectes Soc 14:95–99
Leston D (1973) The ant mosaic—tropical tree crops and the limiting of pests and diseases. PANS 19:311–341
Longino JT (2002) Ants of Costa Rica homepage (http://www.evergreen.edu/ants/genera/camponotus/species/novogranadensis/novogranadensis.html)
Majer JD (1978) The maintenance of the ant mosaic in Ghana cocoa farms. J Appl Ecol 13:123–144
Majer JD, Queiroz MVB (1993) Distribution and abundance of ants in a Brazilian subtropical coffee plantation. P N G J Agric For Fish 36:29–35
Majer JD, Delabie JHC, Smith MRB (1994) Arboreal ant community patterns in Brazilian cocoa farms. Biotropica 26:73–83
Mas AH, Dietsch TV (2003) An index of management intensity for coffee agroecosystems to evaluate butterfly species richness. Ecol Appl 13:1491–1501
Moguel P, Toledo VM (1999) Biodiversity conservation in traditional coffee systems of Mexico. Conserv Biol 13:11–21
Perfecto I, Vandermeer J (1996) Microclimatic changes and the indirect loss of ant diversity in a tropical agroecosystem. Oecologia 108:577–582
Philpott SM, Armbrecht I (2006) Tropical agroforests and the ecological role of ants. Ecol Entomol (In press)
Philpott SM, Perfecto I, Vandermeer J (2006) Effects of management intensity and season on arboreal ant diversity and abundance in coffee agroecosystems. Biodivers Conserv 15:139–155
Ribas C, Schoereder J (2002) Are all ant mosaics caused by competition? Oecologia 131:606–611
Room P (1975) Relative distributions of ant species in cocoa plantations in Papua New Guinea. J Appl Ecol 12:47–62
Soares SM, Schoereder JH (2001) Ant-nest distribution in a remnant of tropical rainforest in southeastern Brazil. Insectes Soc 48:280–286
Theunis L, Gilbert M, Roisin Y, Leponce M (2005) Spatial structure of litter-dwelling ant distribution in a subtropical dry forest. Insectes Soc 52:366–377
Vandermeer J, Perfecto I, Ibarra-Nunez G, Philpott S, Garcia-Ballinas JA (2002) Ants (Azteca sp.) as potential biological control agents in organic shade coffee production in Southern Chiapas, Mexico: complication of indirect effects. Agrofor Syst 56:271–276
Acknowledgements
J. Maldonado, J.A. García-Ballinas, G. López, B.E. Chilel, J.C. Méndez López, A. Méndez Mendizábal, A. Hammond, F.B. Camposeco Silvestre, J.L. Cabrera Santos, F. Hernández Gómez, S. Uno, and P. Bichier helped with data collection. G. Estabrook and D. Schoolmaster helped with development of the models. J. Vandermeer, I. Perfecto, R. Burnham, B. Rathcke, and P. Bichier commented on the manuscript. The Peters and Edelman families, the owners of Guadalupe Zajú, and the ISMAM Cooperative allowed us to work on their farms. G. Ibarra-Núñez, J.A. García-Ballinas, and G. López from El Colegio de la Frontera Sur in Tapachula provided logistical support. This research was supported by NSF grant #DEB-9981526, Helen Olsen Brower Fellowship in Environmental Science of the University of Michigan, and NSF Graduate Research Fellowship to SP. The experiments comply with the current laws of the country in which they were performed.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Philpott, S.M. Ant patchiness: a spatially quantitative test in coffee agroecosystems. Naturwissenschaften 93, 386–392 (2006). https://doi.org/10.1007/s00114-006-0123-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00114-006-0123-0