
Abstract
The functional relationship between apoptosis and autophagy in anticancer drug treatment is extremely complex, and the molecular machinery is obscure. This study aims to investigate the efficacy of CYT997, a novel microtubule-disrupting agent, in head and neck squamous cell carcinomas (HNSCCs) and complete the autophagy-apoptosis puzzle involved in drug action. We report here that CYT997 exhibits anticancer activity by triggering oxidative stress-associated apoptosis in HNSCC cells. Interestingly, upregulation of autophagy by mTOR-dependent pathways appears to have a cytoprotective role in preventing apoptosis by inhibiting CYT997-induced excessively high levels of reactive oxygen species (ROS). Blockade of autophagy by ATG7 depletion or addition of autophagy inhibitor hydroxychloroquine (HCQ) sensitizes HNSCC cells to CYT997 as evidenced by enhanced ROS-associated apoptosis. Moreover, HCQ exhibits a good synergism with CYT997 on induction of apoptosis in HNSCC xenografts without cytotoxicity, suggesting combined treatment of CYT997 with autophagy inhibitors would increase the anticancer efficacy of CYT997. These findings unveil the importance of ROS in crosstalk between autophagy and apoptosis in CYT997 treatment, raising concerns that genetic or pharmacologic blockade of autophagy should be considered in the design of new therapeutics for HNSCC.
Key messages
• CYT997 exhibits anticancer activity by induction of ROS-associated apoptosis.
• mTOR-dependent cytoprotective autophagy prevents CYT997-induced apoptosis.
• Blockade of autophagy augments CYT997 efficacy by enhanced ROS-associated apoptosis.
• Combination of autophagy inhibitors with CYT997 is more effective against HNSCC.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Microtubules are filamentous intracellular structures that are responsible for cell division and movement [1]. Dysfunction of microtubules is often seen in cancer, making them a very suitable target for the development of chemotherapeutic drugs against rapidly dividing and aggressive tumor cells. Several clinical microtubule-targeting drugs potently suppress microtubule dynamics without affecting microtubule polymer mass and retain their ability to block mitotic progression and induce apoptosis [2,3,4]. Such drugs are frontline treatments for a wide variety of human cancers, including breast, prostate, and head and neck cancers [5,6,7]. Although microtubule-targeting agents have substantial drawbacks, their effectiveness in cancer therapy has inspired new generation drug discovery.
Paclitaxel has long been recognized as a microtubule-stabilizing drug, leading to cell death in a subset of the arrested population. Disappointingly, recent evidence demonstrates that intratumoral concentrations of paclitaxel are too low to cause mitotic arrest and result in multipolar cell division instead [8]. CYT997, a novel microtubule-disrupting agent screened from Cytopia’s small molecule library, has addressed the shortcomings of the classic agents such as the need for intravenous administration [9]. Besides its anti-angiogenic activity, CYT997 has recently been shown to possess varying degrees of activity against cancers [10]. By inhibiting tubulin polymerization and disrupting cellular microtubules, CYT997 effectively kills acute myeloid leukemia cells [11]. In addition, this drug has potent cytotoxic activity in vitro and oral activity in prostate cancer xenograft models [12]. The efficacy and safety of CYT997 in patients with solid malignant tumors have been investigated in Phase I trials, and it is now in Phase II clinical trials for the treatment of selected cancers [9].
Head and neck squamous cell carcinoma (HNSCC) is a devastating disease with limited treatment options [13,14,15]. In this study, we present evidence for the first time that CYT997 effectively inhibits growth and viability of HNSCC cells and induces reactive oxygen species (ROS)-associated apoptosis. We also show that CYT997 triggers autophagy through inhibiting mTOR-dependent signaling, which attenuates drug-induced apoptotic death. Autophagy blockade sensitizes the HNSCC cells to CYT997, resulting in ROS-induced apoptosis. These findings provide a rational basis for CYT997 in the treatment of HNSCC and strongly suggest that inhibition of autophagy may enhance the therapeutic efficacy of CYT997.
Materials and methods
An expanded “Materials and methods” section is available in the online-only Data Supplement.
HNSCC xenograft, drug administration, and immunohistochemistry
Six-week-old female NSG (NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ) mice were purchased from the Jackson Laboratory (Bar Harbor, ME) and housed in a specific pathogen-free animal facility. The tumor-bearing NSG mouse model was used as described previously [16, 17]. In brief, mice were injected subcutaneously with 1 × 106 HN12 cells, then randomized for different treatment when the established xenografts reach a size of 100 mm3. The mice were treated with vehicle, HCQ (60 mg/kg once every other day by oral gavage), CYT997 (15 mg/kg twice daily by oral gavage), or in combination of HCQ and CYT997, respectively. Tumor growth was measured externally every 3 to 5 days using vernier calipers as length × width2 × 0.52. The mice were sacrificed on treatment day 16, and the xenografts were removed and processed for immunohistochemistry (IHC) with antibodies against c-PARP and CD31 as described previously [17, 18]. All animal experiments were reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) of Augusta University.
Results
CYT997 effectively inhibits cell viability and induces apoptosis in HNSCC cells
To determine the effective dose range of CYT997 in HNSCC cells, three cell lines, HN4, HN12, and HN30, were treated with CYT997 concentrations ranging from 20 to 200 nM for 48 h. CYT997 exhibited strongly proliferative inhibitory effects with IC50 of 100 nM, and these effects were dose-dependent (Fig. 1a). Cell growth assays confirmed these observations and indicated that HN4 cells were more sensitive to CYT997 at 100 nM during the first 48 h compared with HN12 and HN30 (Fig. 1b). We next determined cell viability at different time points using 100 nM of CYT997. Cell death was increased within the CYT997 treatment period (0–72 h) (Fig. 1c). To study the possible mechanisms that may underlie CYT997-induced cell death, we determined apoptosis in the presence or absence of CYT997. A significantly increased apoptotic rate was found following CYT997 exposure (Fig. 1d), which was accompanied by increased levels of cleaved PARP and caspase 3 (Fig. 1e).

CYT997 inhibits cell viability and induces apoptosis in HNSCC cells. a HNSCC cells were treated with the indicated concentrations of CYT997 for 48 h, and cell viability was determined by MTS assays. b HNSCC cells were seeded into six-well plates and treated with 100 nM CYT997 for 24 h, and the number of attached cells was determined by crystal violet staining. Representative images were shown in the left panel and quantitative data are shown in the right panel. c HNSCC cells were treated with 100 nM CYT997 for the indicated times, and cell viability was determined by MTS assays. d, e HNSCC cells were treated with the indicated concentrations of CYT997 for 24 h, and apoptosis was determined by flow cytometry with Annexin V-FITC staining (d) and Western blot with the antibodies against cleaved PARP and caspase3 (e). *p < 0.05; **p < 0.01
CYT997-induced apoptosis is associated with elevated oxidative stress in HNSCC cells
Oxidative stress refers to elevated intracellular levels of ROS that often leads to cell death [19]. To examine the effect of oxidative stress on CYT997-induced cell apoptosis, the DCFH-DA fluorescent reagent was used to evaluate ROS production, which showed a significant increase of fluorescence in the cells following exposure to CYT997 (Fig. 2a). These observations suggest that CYT997 induces HNSCC cell death, at least in part, by enhancing oxidative stress. We then monitored O2•− release using electrochemical biosensors [20, 21]. The maximal amount of O2•− generation was increased more than 20% in HN4 and HN30 cells and 12% in HN12 cells after CYT997 treatment, as compared with untreated control cells, respectively (Fig. 2b). HN4 and HN12 cells reached a maximum O2•− level at 8 h after CYT997 treatment and then decreased (Fig. 2b), whereas O2•− in HN30 cells peaked at 4 h after CYT997 treatment (Fig. 2b), suggesting the CYT997-induced O2•− release is cell line dependent. These data indicate that elevated intracellular O2•− and ROS levels in HNSCC cells contribute to CYT997-induced oxidative stress. NAC is an antioxidant that has the ability to minimize oxidative stress and the associated downstream negative effects [22]. To test the possibility that suppression of an increase in the amount of ROS could block CYT997-induced apoptosis, we treated HN12 cells with CYT997 in the presence or absence of NAC. NAC significantly attenuated the induction of ROS by CYT997 (Fig. 2c), which also alleviated PARP cleavage following CYT997 treatment (Fig. 2d).

CYT997 promotes oxidative stress-associated apoptosis in HNSCC cells. a, b HNSCC cells were treated with 100 nM CYT997 for the indicated times. ROS generation was determined and quantitated by DCFH-DA staining (a), and O2•− generation was determined by electrochemical biosensor (b). c, d HN12 cells were treated with 100 nM CYT997 for 8 h, in the presence or absence of 5 mM NAC. ROS generation was quantitated by DCFH-DA staining (c), and apoptosis was determined by Western blot with an antibody against cleaved PARP and flow cytometry with Annexin V-FITC staining (d)
CYT997 triggers mTOR-dependent autophagy in HNSCC cells
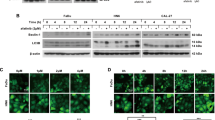
Autophagy is regulated by numerous stresses such as hypoxia, oxidative stress, and chemical drugs [23, 24]. The observation of intracellular ROS levels in CYT997-treated HNSCC cells prompted us to investigate the autophagic changes following treatment with CYT997. Interestingly, CYT997-treated cells had the prominent morphological features with the appearance of numerous cytoplasmic vesicles and vacuoles (Fig. 3a). We then explored autophagy activation by staining with dansylcadaverine, an autofluorescent compound used for the labeling of autophagic vacuoles. Compared with the control group, dot-like structures in the cytoplasmic and perinuclear regions of the drug-treated cells were detected (Fig. 3a), suggesting that CYT997 can induce autophagosome formation. Electron microscopy further revealed numerous double-membraned vacuoles in CYT997-treated cells that contained fragments of the endoplasmic reticulum and other cytoplasmic components (Fig. 3b). Consistent with these observations, reduced protein levels of p62 (Fig. 3c), an effective marker to estimate autophagic flux [25], was observed following exposure to CYT997 in all three HNSCC cell lines examined. LC3B-II levels correlate with the steady-state levels of autophagosomes [26]. Increased accumulation of LC3B-II was also found in CYT997-treated cells (Fig. 3c), suggesting that CYT997 is a potent inducer of autophagy.

CYT997 induces mTOR-dependent autophagy in HNSCC cells. a, b HN12 cells were treated with 100 nM CYT997 for 24 h, and autophagy was determined by dansylcadaverine staining (a) and TEM (b). In b, phagophores (a), autophagosomes (b), lysosomes (c), and autolysosomes (d) were indicated. c HNSCC cells were treated with 100 nM CYT997 for 24 h, and cell lysates were collected for Western blot
Several signaling cascades can regulate autophagy in mammalian cells, including the mammalian target of rapamycin (mTOR)-dependent and independent pathways [27, 28]. We thus determined the activation of mTOR signaling in the presence or absence of CYT997. mTOR is a major negative regulator of autophagy, and p70 ribosomal S6 kinase (p70S6K) is an in vitro substrate for mTOR that subsequently phosphorylates the S6 ribosomal protein [29, 30]. The phosphorylation of mTOR, p70S6K, and S6 was significantly inhibited by CYT997 in all cell lines examined in this study (Fig. 3c and Supplementary Fig. S1), indicating that CYT997 induces autophagy, at least in part, through the mTOR-p70S6K signaling.
Knockdown of ATG7 enhances CYT997-induced apoptosis in HNSCC cells
Autophagy-related (ATG) proteins are engaged in autophagosome formation. We then assessed the expression of ATG proteins in CYT997 treatment. The level of ATG7 was markedly increased following CYT997 exposure in all three cell lines examined (Fig. 4a). The levels of ATG3 and ATG5 were only increased in HN4 and HN12 cells, but not in HN30 cells (Fig. 4a), suggesting that ATG7 functions as an essential enzyme for CYT997-induced autophagy. To determine whether ATG7 is a critical player between autophagy and apoptosis triggered by CYT997, we inhibited its expression in HN12 cells using shRNA (Fig. 4b). With CYT997 treatment, higher levels of cleaved PARP (Fig. 4c) and increased apoptotic rate (Fig. 4d) were observed in cells with ATG7 loss compared with control cells, suggesting ATG7-dependent autophagy attenuates CYT997-induced apoptosis in HNSCC cells.

Inhibition of ATG7 enhances CYT997-induced apoptosis in HNSCC cells. a HNSCC cells were treated with the indicated concentrations of CYT997 for 24 h, and cell lysates were collected for Western blot. Representative images of this assay are shown in the left panel and quantitative data of ATG7 protein levels are shown in the right panel. b–d HN12 cells were infected with lentiviral particles containing shRNA against ATG7, and the knockdown effect was determined by Western blot with an ATG7 antibody (b). The stable ATG7 knockdown HN12 and control cells were treated with 100 nM CYT997 for 24 h, and apoptosis was determined by Western blot with an antibody against cleaved PARP (c) and flow cytometry with Annexin V-FITC staining (d). *p < 0.05; **p < 0.01
Blockade of autophagy by HCQ promotes CYT997-induced apoptosis through enhancing ROS in HNSCC cells
Autophagy inhibitors (e.g., HCQ and 3-MA) are often used in combination with chemotherapeutic drugs to inhibit autophagic survival and exert their anticancer action [31]. As autophagy was induced more efficiently by CYT997 in HN12 cells compared with HN4 and HN30 cells (Fig. 3c), we first evaluated the role of HCQ in CYT997 treatment using this cell line. As expected, HCQ can inhibit autophagosome formation induced by CYT997 (Fig. 5a, b). Although there were no discernible differences in cell growth and viability in the presence or absence of HCQ, a significant decrease in cell growth and viability (Fig. 5d, e) and an increase in apoptosis (Fig. 5f, g) were observed for the addition of HCQ to CYT997. The similar tendency was seen from HN4 and HN30 cells when co-treated with HCQ and CYT997 (Supplementary Fig. S2), suggesting that inhibition of autophagy is broadly applicable for enhancing CYT997-induced apoptosis in HNSCC cells. To study whether the crosstalk between autophagy and apoptosis by CYT997 depends on its microtubule inhibition, we treated HN12 cells with the other microtubule depolymerizer colchicine [32]. Similar to CYT997, colchicine trigged both autophagy and apoptosis as evidenced by increased accumulation of LC3B-II and c-PARP (Supplementary Fig. S3A). Most importantly, the combined treatment with HCQ and colchicine also increased apoptotic response in HN12 cells than colchicine given alone (Supplementary Fig. S3B and S3C). These observations suggest that autophagy blockade can augment apoptosis induced by microtubule depolymerization. 3-MA mainly suppresses the mTOR-independent induction of autophagy by inhibiting the activity of class III PI3K complexes; however, high concentrations of 3-MA can modulate autophagy via inhibition on both class I and III PI3K [33]. To determine the role of 3-MA in HNSCC cells, 25 μM 3-MA was used to treat HN12 cells (Supplementary Fig. S4). Surprisingly, 3-MA downregulated mTOR signaling, which was confirmed by its suppressive effect on S6 protein phosphorylation (data not shown), a surrogate marker for mTOR activity. We next determined the efficacy of CYT997 in combination with 3-MA. Unlike HCQ, 3-MA did not have a notable effect on CYT997-induced apoptotic death and repression of cell survival (Supplementary Fig. S4), which may be attributed to incapability of 3-MA in inhibiting autophagosome maturation and autolysosome formation [34].

Addition of HCQ sensitizes HNSCC cells to CYT997-induced apoptotic death through enhancing ROS. a–g HN12 cells were treated with 100 nM CYT997 for 24 h in the presence or absence of 20 μM HCQ. Autophagy was determined by dansylcadaverine staining (a) and TEM (b; arrow indicates autophagic vacuoles and quantitative data were shown in the right panel; n = 20). The number of survival cells was determined by crystal violet staining (c), cell viability was determined by MTS assays (d) and flow cytometry with an amine-reactive fluorescent dye Zombie Aqua™ that is non-permeant to live cells but permeant to cells with compromised membranes (e), and apoptosis was determined by flow cytometry with Annexin V-FITC staining (f) and Western blot with an antibody against cleaved PARP (g). h, i ROS generation was determined and quantitated by DCFH-DA staining (h; representative images and quantitative data are shown in the left and right panel, respectively), and O2•− generation was determined by electrochemical biosensor (i). j Schematic representation of CYT997-induced interplay between autophagy and apoptosis in HNSCC cells. *p < 0.05; **p < 0.01
To investigate the mechanism underpinning CYT997-induced interplay between autophagy and apoptosis, we determined the status of oxidative stress in single or combination treatment. Although either HCQ or CYT997 had the capacity to promote ROS generation and O2•− release in HN12 cells, their combination induced much higher levels of oxidative stress than the single agents alone (Fig. 5h, i). Taken together, these data illustrate that the mTOR-dependent autophagy triggered by CYT997 suppresses drug-induced apoptosis through inhibiting excessively high ROS (Fig. 5j).
HCQ augments anticancer activity of CYT997 in a xenograft mouse model of HNSCC
We next evaluated the synergistic effects of CYT997 and HCQ on tumorigenic potential. HN12 cells were implanted into the right flank of NSG mice and measurement of tumor volume was commenced 5 days following implantation. The tumor-bearing animals were randomly assigned to treatment (drug alone or in combination) or control groups. Treatment with CYT997 significantly reduced the size and weight of HN12-derived xenografts (Fig. 6a, b). HCQ did not exert any effect on tumor growth and body weight, but it augmented the anticancer effects of CYT997 in the subcutaneous xenograft model (Fig. 6a–c). As a microtubule binding agent, CYT997 exhibits some degree of vascular disrupting activity [10]. Thus, we assessed the in vivo antivascular ability of CYT997. IHC staining with a CD31 antibody revealed that microvessel density (MVD) of tumor tissue from CYT997-treated mice was much lower than that from vehicle-treated mice (Fig. 6d, f). Addition of HCQ in CYT997 treatment did not show a significant alteration in MVD compared with the group treated with drug alone (Fig. 6d, f). In contrast, IHC with an antibody against cleaved PARP showed that HCQ improved the anticancer effect of CYT997 by enhancing apoptotic rate in tumor cells (Fig. 6e, g). These data suggest that the combination of HCQ and CYT997 has a superior inhibitory activity on HNSCC tumor survival compared to monotherapy.

Combination of HCQ and CYT997 inhibits HNSCC growth more efficiently than CYT997 treatment alone in a xenograft mouse model. a–c The effect of combination therapy with CYT997 and HCQ on tumor growth. When HN12-derived xenografts have been established, NSG mice were randomly divided into four groups for treatment with vehicle, CYT997, HCQ, or the combination of CYT997 and HCQ (n = 5/group). Tumor growth was measured by tumor volume (a), tumor burden at the end of the experiment was calculated as tumor weight (b), and mouse bodies (minus tumor) were also weighed (c). d–g The xenografts removed from the drug-treated tumor-bearing mice were processed for IHC with the antibodies against CD31 (d) and cleaved PARP (e). Representative images of IHC and quantitative data are shown in d, e and f, g, respectively. n.s., non-significant; *p < 0.05; **p < 0.01
Discussion
The oral activity of CYT997 is a key advantage over most other microtubule-targeting agents, which are administered intravenously, thereby markedly improving its clinical value [9,10,11,12]. Successful practical use of CYT997 will benefit from understanding its efficacy and its underlying mechanisms, alone or in combination with other anticancer agents. This study reports the anticancer activity of CYT997 in HNSCC cells and demonstrates that autophagy triggered by CYT997 attenuates drug-induced apoptosis through suppression of excessively high ROS. We present data indicating that CYT997 together with autophagy inhibitors such as HCQ exhibits potent oncolytic effects on HNSCC cells, suggesting that this polytherapy may be an effective regimen for subsets of HNSCC lesions.
Oxidative stress resulting from elevated intracellular levels of ROS leads to damage to lipids, proteins, and DNA and is suspected to be linked to cellular microtubule dynamics and apoptosis [19, 35]. In this study, we found that disruption of microtubules by CYT997 enhanced ROS in HNSCC cells, which is also evident with increased cellular O2•− release. We also identified that apoptosis accompanied CYT997-mediated ROS accumulation and that NAC, a specific ROS scavenger, can block CYT997-induced apoptosis. These findings indicate that CYT997 exerts pro-apoptotic activity on HNSCC cells, at least partially through ROS generation. In addition, our study indicates that autophagy plays a cytoprotective role following CYT997 treatment, which limits the production of ROS to suppress pro-apoptotic signaling in HNSCC cells.
Autophagy, a self-protective response of living cells or organisms to various stress conditions, is increasingly recognized as an important factor in the initiation and progression of cancer and interventions in cancer therapy [36]. Autophagy promotes cell survival during starvation; however, it can switch from death inhibition to death promotion under different contexts. An evidence from Sentelle and his colleagues shows that the direct interaction between C18-ceramide and LC3B-II on mitochondrial membranes can induce lethal autophagy via selective targeting of mitochondria by LC3B-II-containing autophagolysosomes [37]. Melatonin, a potent free radical scavenger, can be added into rapamycin treatment for HNSCC as it repressed the negative feedback loop from the specific downstream effector of mTOR activation S6K1 to Akt signaling [38]. Interestingly, this combined treatment can simultaneously increase apoptosis and autophagy in HNSCC cells [38], suggesting that autophagy plays a critical role in favor of apoptosis. Therefore, understanding the stimulus- and cell type-specific differences in the autophagy machinery is needed if we consider using this pathway for therapeutic benefit in the future.
Autophagy modulators have two distinct mechanisms of action, acting through mTOR-dependent or mTOR-independent pathways [36, 39]. CYT997 appears to trigger autophagy in HNSCC cells through inhibition of mTOR activity. In this study, we tested two widely used autophagy inhibitors, 3-MA, and HCQ. HCQ blocks autophagic flux that is increased by allosteric inhibition of mTOR, and high concentrations of 3-MA can also inhibit mTOR activity in cancer cells [34, 36]. However, compared with HCQ, 3-MA enhances autophagic flux not due to suppression of maturation of autophagosomes or lysosomal function [34, 40]. To our surprise, HCQ, but not 3-MA, significantly enhanced the cytotoxic effects of CYT997 and increased the rate of apoptosis in HNSCC cells, suggesting that inhibiting autophagosome maturation and autolysosome formation is required to augment CYT997-induced apoptosis. Our study also implicates that selection of suitable autophagy inhibitors as an adjunct to chemotherapy is critical for achieving better therapeutic effectiveness in cancer treatment.
Previous studies have shown that autophagy can inhibit chemotherapy-induced apoptosis in cancer treatment, and interference with the autophagic response may be a good therapeutic strategy in such cases [41, 42]. Consistent with this concept, blockade of autophagic signaling sensitizes HNSCC cells to CYT997, which implies that judicious targeting of autophagy pathways may present an opportunity to improve the effects of microtubule-targeting drugs in the treatment of HNSCC. Based on the rationale that the current study has provided for this combination therapy. Further exploration of autophagy inhibitors such as HCQ combined with CYT997 in HNSCC is warranted.
Abbreviations
- 3-MA:
-
3-methyladenine
- CV:
-
cyclic voltammetry
- DCFH-DA:
-
2′, 7′-dichlorodihydrofluorescein diacetate
- HNSCC:
-
head and neck squamous cell carcinomas
- HCQ:
-
hydroxychloroquine
- IACUC:
-
Institutional Animal Care and Use Committee
- IHC:
-
immunohistochemistry
- JEOL:
-
JEM 1230 transmission electron microscope
- NAC:
-
N-acetylcysteine
- NaCAC:
-
sodium cacodylate
- MVD:
-
microvessel density
- O2 •− :
-
superoxide
- p70S6K:
-
p70 ribosomal S6 kinase
- ROS:
-
reactive oxygen species
- SOD:
-
superoxide dismutase
- TEM:
-
transmission electron microscopy
References
Komlodi-Pasztor E, Sackett D, Wilkerson J, Fojo T (2011) Mitosis is not a key target of microtubule agents in patient tumors. Nat Rev Clin Oncol 8:244–250
Mukhtar E, Adhami VM, Mukhtar H (2014) Targeting microtubules by natural agents for cancer therapy. Mol Cancer Ther 13:275–284
Thomas E, Gopalakrishnan V, Hegde M, Kumar S, Karki SS, Raghavan SC, Choudhary B (2016) A novel resveratrol based tubulin inhibitor induces mitotic arrest and activates apoptosis in Cancer cells. Sci Rep 6:34653
Dumontet C, Jordan MA (2010) Microtubule-binding agents: a dynamic field of cancer therapeutics. Nat Rev Drug Discov 9:790–803
Phillips GDL, Li G, Dugger DL, Crocker LM, Parsons KL, Mai E, Blättler WA, Lambert JM, Chari RV, Lutz RJ (2008) Targeting HER2-positive breast cancer with trastuzumab-DM1, an antibody–cytotoxic drug conjugate. Cancer Res 68:9280–9290
Sahra IB, Laurent K, Giuliano S, Larbret F, Ponzio G, Gounon P, Le Marchand-Brustel Y, Giorgetti-Peraldi S, Cormont M, Bertolotto C (2010) Targeting cancer cell metabolism: the combination of metformin and 2-deoxyglucose induces p53-dependent apoptosis in prostate cancer cells. Cancer Res 70:2465–2475
Riechelmann H, Sauter A, Golze W, Hanft G, Schroen C, Hoermann K, Erhardt T, Gronau S (2008) Phase I trial with the CD44v6-targeting immunoconjugate bivatuzumab mertansine in head and neck squamous cell carcinoma. Oral Oncol 44:823–829
Weaver BA (2014) How Taxol/paclitaxel kills cancer cells. Mol Biol Cell 25:2677–2681
Burns CJ, Harte MF, Bu X, Fantino E, Joffe M, Sikanyika H, Su S, Tranberg CE, Wilson N, Charman SA (2009) Discovery of CYT997: a structurally novel orally active microtubule targeting agent. Bioorg Med Chem Lett 19:4639–4642
Burns CJ, Fantino E, Phillips ID, Su S, Harte MF, Bukczynska PE, Frazzetto M, Joffe M, Kruszelnicki I, Wang B (2009) CYT997: a novel orally active tubulin polymerization inhibitor with potent cytotoxic and vascular disrupting activity in vitro and in vivo. Mol Cancer Ther 8:3036–3045
Chen X, Yang C, Xu Y, Zhou H, Liu H, Qian W (2013) The microtubule depolymerizing agent CYT997 effectively kills acute myeloid leukemia cells via activation of caspases and inhibition of PI3K/Akt/mTOR pathway proteins. Exp Ther Med 6:299–304
Ji Y-T, Liu Y-N, Liu Z-P (2015) Tubulin colchicine binding site inhibitors as vascular disrupting agents in clinical developments. Curr Med Chem 22:1348–1360
Siegel RL, Miller KD, Jemal A (2016) Cancer statistics, 2016. CA Cancer J Clin 66:7–30
Stransky N, Egloff AM, Tward AD, Kostic AD, Cibulskis K, Sivachenko A, Kryukov GV, Lawrence MS, Sougnez C, McKenna A (2011) The mutational landscape of head and neck squamous cell carcinoma. Science 333:1157–1160
Leemans CR, Braakhuis BJ, Brakenhoff RH (2011) The molecular biology of head and neck cancer. Nat Rev Cancer 11:9–22
Davis JE, Xie X, Guo J, Huang W, Chu W-M, Huang S, Teng Y, Wu G (2016) ARF1 promotes prostate tumorigenesis via targeting oncogenic MAPK signaling. Oncotarget 7:39834–39845
Teng Y, Ren X, Li H, Shull A, Kim J, Cowell JK (2015) Mitochondrial ATAD3A combines with GRP78 to regulate the WASF3 metastasis-promoting protein. Oncogene
Davis JE, Xie X, Guo J, Huang W, Chu W-M, Huang S, Teng Y, Wu G (2016). ARF1 promotes prostate tumorigenesis via targeting oncogenic MAPK signaling. Oncotarget 5
Ramanathan B, Jan K-Y, Chen C-H, Hour T-C, Yu H-J, Pu Y-S (2005) Resistance to paclitaxel is proportional to cellular total antioxidant capacity. Cancer Res 65:8455–8460
Gao L, Wang X, Tang Y, Huang S, C-AA H, Teng Y (2017) FGF19/FGFR4 signaling contributes to the resistance of hepatocellular carcinoma to sorafenib. J Exp Clin Cancer Res 36:8–17
Gao L, Teng Y (2016) Exploiting plug-and-play electrochemistry for drug discovery. Future Med Chem 8:567–577
Misu H, Takayama H, Saito Y, Mita Y, Kikuchi A, K-a I, Chikamoto K, Kanamori T, Tajima N, Lan F (2017) Deficiency of the hepatokine selenoprotein P increases responsiveness to exercise in mice through upregulation of reactive oxygen species and AMP-activated protein kinase in muscle. Nat Med 23:508–516
Lisanti MP, Martinez-Outschoorn UE, Chiavarina B, Pavlides S, Whitaker-Menezes D, Tsirigos A, Witkiewicz AK, Lin Z, Balliet RM, Howell A (2010) Understanding the “lethal” drivers of tumor-stroma co-evolution: emerging role (s) for hypoxia, oxidative stress and autophagy/mitophagy in the tumor microenvironment. Cancer Biol Ther 10:537–542
Rubinsztein DC, Gestwicki JE, Murphy LO, Klionsky DJ (2007) Potential therapeutic applications of autophagy. Nat Rev Drug Discov 6:304–312
Mizushima N, Yoshimori T, Levine B (2010) Methods in mammalian autophagy research. Cell 140:313–326
Ding J, Li T, Wang X, Zhao E, Choi J-H, Yang L, Zha Y, Dong Z, Huang S, Asara JM (2013) The histone H3 methyltransferase G9A epigenetically activates the serine-glycine synthesis pathway to sustain cancer cell survival and proliferation. Cell Metab 18:896–907
Sekulić A, Hudson CC, Homme JL, Yin P, Otterness DM, Karnitz LM, Abraham RT (2000) A direct linkage between the phosphoinositide 3-kinase-AKT signaling pathway and the mammalian target of rapamycin in mitogen-stimulated and transformed cells. Cancer Res 60:3504–3513
Dowling RJ, Zakikhani M, Fantus IG, Pollak M, Sonenberg N (2007) Metformin inhibits mammalian target of rapamycin–dependent translation initiation in breast cancer cells. Cancer Res 67:10804–10812
Hosokawa N, Hara T, Kaizuka T, Kishi C, Takamura A, Miura Y, Iemura S-i, Natsume T, Takehana K, Yamada N (2009) Nutrient-dependent mTORC1 association with the ULK1–Atg13–FIP200 complex required for autophagy. Mol Biol Cell 20:1981–1991
Tremblay F, Marette A (2001) Amino acid and insulin signaling via the mTOR/p70 S6 kinase pathway a negative feedback mechanism leading to insulin resistance in skeletal muscle cells. J Biol Chem 276:38052–38060
Sui X, Chen R, Wang Z, Huang Z, Kong N, Zhang M, Han W, Lou F, Yang J, Zhang Q (2013) Autophagy and chemotherapy resistance: a promising therapeutic target for cancer treatment. Cell Death Dis 4:e838
Seiliez I, Belghit I, Gao Y, Skiba-Cassy S, Dias K, Cluzeaud M, Rémond D, Hafnaoui N, Salin B, Camougrand N, Panserat S (2016) Looking at the metabolic consequences of the colchicine-based in vivo autophagic flux assay. Autophagy 12:343–356
Vakifahmetoglu-Norberg H, Xia H-G, Yuan J (2015) Pharmacologic agents targeting autophagy. J Clin Invest 125:5–13
Wu YT, Tan HL, Shui G, Bauvy C, Huang Q, Wenk MR, Ong CN, Codogno P, Shen HM (2010) Dual role of 3-methyladenine in modulation of autophagy via different temporal patterns of inhibition on class I and III phosphoinositide 3-kinase. J Biol Chem 85:10850–10861
Teng Y, Cai Y, Pi W, Gao L, Shay C (2017) Augmentation of the anticancer activity of CYT997 in human prostate cancer by inhibiting Src activity. J Hematol Oncol 10:118
Gao L, Jauregui CE, Teng Y (2017) Targeting autophagy as a strategy for drug discovery and therapeutic modulation. Future Med Chem 9:335–345
Sentelle RD, Senkal CE, Jiang W, Ponnusamy S, Gencer S, Selvam SP, Ramshesh VK, Peterson YK, Lemasters JJ, Szulc ZM (2012) Ceramide targets autophagosomes to mitochondria and induces lethal mitophagy. Nat Chem Biol 8:831–838
Owino S, Sánchez-Bretaño A, Tchio C, Cecon E, Karamitri A, Dam J, Jockers R, Piccione G, Noh HL, Kim T (2018) Nocturnal activation of melatonin receptor type 1 signaling modulates diurnal insulin sensitivity via regulation of PI3K activity. J Pineal Res 64:e12462
Murrow L, Debnath J (2013) Autophagy as a stress-response and quality-control mechanism: implications for cell injury and human disease. Ann Rev Pathol 8:105–137
Kania E, Pająk B, Orzechowski A (2015) Calcium homeostasis and ER stress in control of autophagy in cancer cells. Biomed Res Int 2015:352794
Jain MV, Paczulla AM, Klonisch T, Dimgba FN, Rao SB, Roberg K, Schweizer F, Lengerke C, Davoodpour P, Palicharla VR (2013) Interconnections between apoptotic, autophagic and necrotic pathways: implications for cancer therapy development. J Cell Mol Med 17:12–29
Guo X, Li D, Hu F, Song J, Zhang S, Deng W, Sun K, Zhao Q, Xie X, Song Y (2012) Targeting autophagy potentiates chemotherapy-induced apoptosis and proliferation inhibition in hepatocarcinoma cells. Cancer Lett 320:171–179
Acknowledgements
This work was supported in part by Dental College of Georgia Special Funding Initiative and a grant from the Department of Defense (W81XWH-14-1-0412) (to Y.T.). We would like to thank the staff of the Electron Microscopy and Histology Core at Augusta University for assistance with EM data collection.
Funding
This work was supported in part by Dental College of Georgia Special Funding Initiative and a grant from the Department of Defense (W81XWH-14-1-0412) (to Y.T.).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The study was reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) of Augusta.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Electronic supplementary material
ESM 1
(DOCX 408 kb)
Rights and permissions
About this article

Cite this article
Gao, L., Zhao, X., Lang, L. et al. Autophagy blockade sensitizes human head and neck squamous cell carcinoma towards CYT997 through enhancing excessively high reactive oxygen species-induced apoptosis. J Mol Med 96, 929–938 (2018). https://doi.org/10.1007/s00109-018-1670-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00109-018-1670-5