
Abstract
c-Myc is a well-characterized proto-oncogene that induces cellular transformation and modulates programmed cell death. While recent studies have demonstrated high expression of c-Myc protein in advanced and metastatic melanoma, the clinical and biological implications remain to be fully elucidated. In this study, we investigated the effect of c-Myc overexpression in melanoma tumorigenesis. Clinicopathological analysis demonstrated that c-Myc expression positively correlated with the formation of vasculogenic mimicry (VM) and linearly patterned programmed cell necrosis (LPPCN). Clinically, high c-Myc expression was significantly associated with distant metastasis and poor prognosis, while biologically, c-Myc overexpression led to significant increases in cell motility, invasiveness and metastasis. Moreover, c-Myc induced the formation of VM and promoted the expression of epithelial-mesenchymal transition (EMT)-associated protein Snail both in vivo and in vitro. High expression of c-Myc increased Bax expression in hypoxic conditions and induced cell apoptosis. Taken together, we conclude that c-Myc overexpression promotes the formation of VM by EMT and LPPCN in melanoma. Our improved understanding of the clinical and biological effects of c-Myc overexpression in melanoma highlights the incomplete understanding of this oncogene, and indicates that c-Myc is a potential therapeutic target of this disease.
Key message
-
High c-Myc expression is associated with tumor metastasis and poor prognosis in human melanoma.
-
c-Myc upregulates Snail expression to promote EMT via the TGF-β/Snail/Ecadherin signal pathway.
-
c-Myc leads to cell death by upregulating Bax expression causing a lower Bcl2/Bax ratio under severe hypoxic conditions.
-
c-Myc promotes vasculogenic mimicry and linearly patterned programmed cell necrosis.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Melanoma is a type of cancer with an aggressive clinical course and poor outcome associated with acquired resistance to existing therapies [1]. Tumor growth and metastasis depend on angiogenesis [2]. Inhibition of angiogenesis has become a clinical anti-cancer strategy [3–7]. Vascular mimicry (VM) represents an extension of tumor angiogenesis [8]. VM has been identified in numerous types of cancers including melanoma and is associated with metastasis and a poor prognosis [9]. However, the mechanisms leading to VM remain poorly characterized, and few investigations have evaluated the nature of the structure or formation of VM.
Emerging evidence suggests the involvement of epithelial-mesenchymal transition (EMT) in the formation of VM [10–12]. The physical constraints of the microenvironment around a rapidly growing tumor have led researchers to question how tumors are able to create sufficient space to induce VM. Recently, Sun et al. reported a previously undescribed mechanism of programmed cell death, “linearly patterned programmed cell necrosis” (LPPCN), in which distinct channel-shaped, vessel-like empty spaces are created via cell necrosis thereby providing a spatial structure for the formation of VM and endothelial-dependent vessels (EDVs) [13]. Zhang and colleagues recently highlighted the importance of LPPCN, describing a correlation between this phenomenon and both metastasis and poor prognosis in melanoma. Ultimately, the biological triggers leading to LPPCN cell death remain unknown.
c-Myc is a proto-oncogene known to promote cell proliferation, resulting in cellular transformation and modulation of programmed cell death, among a multitude of other functions [14–17]. Cellular transformation and cell death are the opposing cellular fates. In 2001, Kraehn et al. reported c-Myc amplification in advanced and metastatic melanoma [18]. However, the role of c-Myc in the complex microenvironment of melanoma has not been well described. In the present study, we evaluated the effect of c-Myc protein overexpression on the formation of VM via promotion of both cellular transformation (EMT) and cell death (LPPCN).
Materials and methods
Collection of patient samples
A total of 135 primary tumor specimens were obtained from the Tumor Tissue Bank of the Tianjin Cancer Hospital (Tianjin, China). Specimens were excised from melanoma patients who underwent surgical resection at the Tianjin Medical University Cancer Institute and Hospital, in China, between January 1999 and October 2005, and July 2006 and December 2010.
All tissue sections were reviewed by a pathologist according to the 2010 World Health Organization Classification of Tumors Pathology and Genetics of Skin Tumors. The use of patient specimens was approved by the Institutional Research Committee.
CD34 (or Endomucin for mice tissue) immunohistochemistry and periodic acid-schiff (PAS) staining
The tissue sections were deparaffinized, hydrated and rehydrated according to standard protocols. Antigen retrieval was performed, and non-specific binding sites were blocked. The sections were then incubated with the primary antibody overnight at 4 °C and the second antibody for 30 min at 37 °C. The color was developed using 3,3′-diamino-benzidine-chromogen (DAB). Finally, the sections were subjected to PAS staining and counterstained with hematoxylin.
Immunohistochemical staining methods was performed as above without PAS staining. Details of the primary antibody are shown in Supplementary Table S1.
Immunohistochemical scoring
Evaluation of sections was performed by two independent pathologists. The sections were semi-quantitatively assessed according to both the percentage of positive neoplastic cells and the immunostaining intensity of individual tumor cells (extension + intensity) [19]. For statistical analysis, a total score of 0–2 was considered as negative/low expression, whiles scores of 3–6 were considered as positive/high expression.
Melanoma cell lines and animals
Melanoma cell lines A375, MUM-2C, and MUM-2B were obtained from China Infrastructure of Cell Line Resources (Beijing, China). Forty male NIH BALB/c nu/nu mice (4–6 weeks of age), were obtained from HFK Bioscience Co. Ltd. (Beijing, China). The mice were housed in the Animal Center of Tianjin Medical University (Tianjin, China) at 1 week before experiments started and bred under specific pathogen-free conditions. All mice were handled according to the recommendations of the NIH Guidelines for the Care and Use of Laboratory Animals. The experiment was approved by the Institutional Animal Care and Use Committee.
c-Myc expression plasmids and c-Myc gene silencing
To functionally characterize the relationships of c-Myc with VM and LPPCN, we established stable c-Myc-expressing cells. c-Myc suppression was mediated by lentivirus infection using OmicsLink short hairpin RNA (shRNA) Expression Clones (target sequence: ACAGCTACGGAACTCTTGT, Catalog no. HSH011597-HIVmU6; GeneCopoeia) (shown in Figures as “sh-c-Myc”), whereas c-Myc overexpression was achieved by an ORF lentiviral expression clone with high expression of the c-Myc gene (Catalog no. EX-Z2845-Lv105–5) (shown in Figures as “c-Myc”). A non-silencing shRNA sequence without the c-Myc shRNA component was used as the negative control (Catalog no. CSHCTR001-HIVmU6) (shown in Figures as “vector”). Cells were transfected using a Lenti-Pac HIV Expression Packaging Kit (Catalog no. HPK-LvTR-40) according to the manufacturer’s protocol. Stably expressing cell lines were obtained by selection with 0.2 μg/ml puromycin. The transfection efficiency was confirmed by western blot analysis and semi-quantitative RT-PCR.
Transwell assay, wound healing assay and matrigel-based tube formation assay
Transwell assay, wound healing assay and matrigel-based tube formation assay were performed as previously mentioned [20]. The number of invading/migrating cells was counted in five fields under an inverted light microscope (Nikon) at ×200 magnification. The speed of wound closure was monitored after 12 and 24 h by measuring the ratio of the distance of the wound at 0 h. The cells of matrigel-based tube assay were photographed at ×200 magnification. Each experiment was performed in triplicate.
Immunofluorescence
Cells were plated onto coverslips and fixed in ice cold methanol for 10 min. The cells were blocked with 5 % FBS and incubated with the primary antibodies and the FITC-conjugated secondary antibodies. The sections were counterstained with 4,6-diamidino-2-phenylindole and observed under a fluorescence microscope (80i, Nikon) at ×200 magnification.
Western blot analysis
Blots were blocked and incubated with antibodies and the enhanced chemiluminescence method was used to measure protein expression. Bands were imaged and analyzed using a C-Digit ®Blot Scanner (Gene Company).
Co immunoprecipitation (Co-IP) assay
The Co-IP products of c-Myc/Snail were immunoprecipitated from melanoma cell lysates by a Pierce Crosslink Immunoprecipitation Kit (Thermo Fisher Scientific). Mouse IgG was used as a control. Input and immunoprecipitated proteins were analyzed by western blotting.
Luciferase reporter gene assay
The VE-cadherin, c-Myc and Snail promoters were purchased from GeneCopoeia. VE-cadherin and Snail promoters was cotransfected with c-Myc overexpression plasmids into A375, MUM-2B cell lines by way of percutaneous ethanol injection. Transactivation assays were performed with the Secrete-PairTM Dual Luminescence Assay Kit (GeneCopoeia). Luciferase activities were measured using the Synergy 2 microplate reader system (Gene Company). TGF-β was purchased from PEPROTECH and used at 30 ng/ml.
Semi-quantitative reverse-transcription polymerase chain reaction (RT-PCR)
Total RNA was reverse transcribed using a QuantScript RT Kit (Tiangen Biotech). Semi-quantitative RT-PCR was performed according to the recommended thermal profile. The amplified products were subjected to electrophoresis in a 1 % agarose gel containing ethidium bromide (BIO-RAD Company). The expression of GAPDH was used to standardize the amount of cDNA added to each PCR tube. The primers used for semi-quantitative RT-PCR are shown in Supplementary Table S3.
Cell proliferation and apoptosis assay
Cells were plated in triplicate and analyzed over time using a 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay read on a Synergy 2 plate reader (Bio Tek). In addition, cells with or without Bcl2 or Bax siRNA treatment were incubated with 1 % O2 for 12 h and then collected and stained using an Annexin V-FITC/PI Kit (KeyGen Biotech) according to the manufacturer’s instructions. Apoptotic cells were detected and analyzed using a flow cytometer (Accuri C6; BD, USA).
CoCl2 treatments
The cells were seeded in a 6-well plate and grown to 70 % confluence in complete medium. The medium was then removed and the cells were washed with phosphate-buffered saline. The cells were treated with CoCl2 at 0, 100, 200, 300, 400, and 800 μM and then incubated for 6, 12, 24, and 36 h. Then, semi-quantitative RT-PCR and western blotting were performed to detect mRNA and protein expression. The results were analyzed by multi-factor analysis of variance (ANOVA).
Murine xenograft model, metastasis assay, and colonization assay
Six-week-old male NIH BALB/c-null mice were housed in the animal facilities of Tianjin Medical University as approved by the Institutional Animal Care and Use Committee. All mice were handled according to the recommendations of the Guidelines for the welfare and use of animals in cancer research [21]. Approximately 1 × 106 melanoma cells were injected into the backs of nude mice (N = 10 per group). Following a 12 day observation period, the mice were sacrificed. For metastasis assay, approximately 1 × 106 of melanoma cells were injected into the backs of nude mice (N = 6 per group). Moreover, for colonization assay, 4 × 105 cells were injected via the tail veins of mouse. All of these mice were allowed to proliferate for 10 weeks (N = 6 per group). The tumors, lungs and livers were harvested, embedded in paraffin, and stained with H&E.
Statistical analysis
All data were analyzed with SPSS 17.0 (SPSS, Chicago, IL, USA). Measurement data are shown as the mean ± standard error of mean. P < 0.05 was considered as statistically significant. Kaplan Meier survival analysis was also performed. Differences in survival curves were assessed using the log-rank test. The Student’s t test was performed to determine differences between two groups. Measurement data in more groups were compared by ANOVA.
Results
c-Myc expression, VM and LPPCN in melanoma and their correlation with clinicopathological parameters and prognosis
We respectively evaluated the expression of c-Myc, VM and LPPCN across a cohort of 135 melanoma specimens collected from Cancer Hospital of Tianjin Medical University. Sixty-nine (51.1 %) cases in our cohort were positive for c-Myc (Table 1 and Fig. 1A).

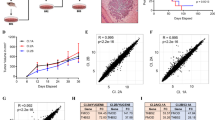
c-Myc expression, phenomena of VM and LPPCN in melanoma. A IHC staining of c-Myc a for negative and b for positive nuclear expression, (×200); B Phenomena of VM (red arrow, CD34-negative and PAS-positive) and EDV (green arrow, CD34-positive and PAS-positive) in melanoma specimens (×400); C LPPCN phenomena. Cells that underwent LPPCN are distributed in patterns of lines and/or networks (a and b; ×200) among the environmental tumor cells and darkly stained (c-e; ×400 ). Inflammatory cells are absent in the environmental district (c; ×400). These networks come with neovasculogenesis networks (b and d; ×200); D Kaplan-Meier survival curve demonstrated that high c-Myc expression, VM and LPPCN are significantly related to poor prognosis (p < 0.05, for all)
Tube cavities lined with PAS-positive, CD34-negative tumor cells and red blood cells observed within cavities were deemed to be VM channels (red arrows, Fig. 1B). Dual PAS and CD34 double positive channels were defined as EDVs (green arrows, Fig. 1B).
H&E staining showed LPPCN as dark cells with loss of cell-cell adhesion, concentrated cytoplasm, a nucleus exhibiting pyknosis, karyorrhexis or karyolysis, and homogeneous chromatin distributed in patterns of lines and/or networks (Fig. 1Ca-e). LPPCN cells displayed an integrated plasma membrane without an inflammatory reaction [22] (Fig. 1C-b and c). The LPPCN lines and/or networks were occasionally accompanied by neovasculogenic networks (Fig. 1C-d and e).
In 62 (45.9 %) specimens, VM formation was observed, while LPPCN was detected in 55.5 % (75/135) of the analyzed melanoma tissues. Notably, our results demonstrated that high c-Myc overexpression, VM and LPPCN respectively correlate with the tumor’s AJCC staging, Breslow’s depth, metastasis of tumors, and poor prognosis (P < 0.05, Tables 2 and 3, and Fig. 1D).
c-Myc expression correlates to the presence of VM and LPPCN in melanoma
In c-Myc positive cases, VM was identified in 41 (59.4 %) patients, while 53 (76.81 %) melanoma cases demonstrated LPPCN. We also observed a significant association of c-Myc expression with the formation of VM and LPPCN (P = 0.001, and P < 0.001, respectively). Our results suggest that c-Myc expression positively correlates to the formation of VM and LPPCN.
High c-Myc expression promotes cell migration, invasion, VM formation and proliferation in vitro
Cell migration and invasion
To evaluate the role of c-Myc in regulating the invasion and migration of melanoma cells, we performed the invasion/migration assay, and wound healing assays. Our results demonstrated that c-Myc knockdown significantly decreased the abilities of A375 and MUM-2B cells to invade and migrate, whereas c-Myc overexpression had the inverse effect in A375 and MUM-2C cells. (Fig. 2A and B, p < 0.05 for all).

High c-Myc expression promotes cell invasion, migration, VM formation. A The invasion and migration abilities of melanoma cells were increased following c-Myc overexpression, and decreased by c-Myc knockdown (bar 200um); B Wound healing assay (×40). Quantitative analysis showed a significant difference at 24 h; C c-Myc overexpression promoted VM formation and Knockdown of c-Myc inhibited the VM formation (bar 100um, *P < 0.05, n = 3); D MTT tests showed high c-Myc expressions increased cell proliferation (*P < 0.05, n = 3)
VM formation
To investigate the function of c-Myc in the formation of VM in vitro, we used a matrigel-based tube formation assay. c-Myc silencing in A375 cells inhibited VM formation on matrigel, whereas c-Myc overexpression facilitated the development of VM tubes. MUM-2B cells formed tube-like structures on the surface of the matrigel, whereas loss of c-Myc triggered the disappearance of these structures within 24 h. Moreover, MUM-2C cells lacking pseudopods formed VM tubes following exogenous overexpression c-Myc. (p < 0.05; Fig. 2C).
Cell proliferation
To further investigate the effect of high c-Myc expression on cell viability and proliferation, we performed MTT assays (Fig. 2D). The results showed that inhibition of c-Myc expression suppressed the growth of A375 and MUM-2B melanoma cells. In contrast, promotion of c-Myc expression accelerated the growth of A375 and MUM-2C cells.
High c-Myc expression promotes EMT by the TGF-β/snail signaling pathway in vitro
c-Myc promotes expression of EMT related proteins
EMT is an important cellular program during tumor migration and invasion. To explore the molecular mechanisms by which c-Myc promotes EMT, we evaluated EMT-related indexs using western blotting, semi-quantitative RT-PCR and immunofluorescence.
Western blot (Fig. 3A) and RT-PCR (Fig. 3B) results confirmed that high c-Myc expression upregualted Snail and downregulated E-cadherin expression. The gray analysis showed that the differences were statistically significant (Fig. 3C). Furthermore, immunofluorescence revealed that high E-cadherin expression and down-regulation of Vimentin, a mesenchymal marker, in MUM-2B cells occurs following c-Myc knockdown. Conversely, MUM-2C cells overexpressing c-Myc demonstrated loss of E-cadherin and up-regulation of Vimentin expression (Fig. 3D).

The influence of c-Myc in EMT related index expression. A and B Knockdown c-Myc induced down-regulation of Snail expression and upregulation of E-cadherin (CDH1), the inverse change was obtained with the overexpression c-Myc; C showed the Gray value analysis of Western blotting and RT-PCR results; D Immunofluorescence staining. Overexpression of c-Myc increased the protein expression of Vimentin and inactivated the expression of E-cadherin and c-Myc silencing expression inhibited Vimentin expression and upregulated E-cadherin expression (bar, 50um)
c-Myc may promote EMT by the TGF-β/snail signaling pathway
To further analyze the signaling pathway of c-Myc in regulation of EMT, we performed western blot, luciferase reporter gene assay and Co-IP assays.
Western blot results showed that TGF-β increased the protein expression of c-Myc and Snail (Fig. 4A). The gray assay showed that the differences were statistically significant (Fig. 4B). Luciferase reporter gene assay results showed that TGF-β (30 ng/ml) enhanced c-Myc gene expression (Fig. 4C). A375 and MUM-2B cells were cotransfected with a Snail promoter-luciferase construct and c-Myc-cDNA in an expression vector. c-Myc overexpression resulted in enhancement of transactivation of Snail promoter activity (Fig. 4D). These results indicated that TGF-β is one of the upstream signaling molecules of c-Myc, and the Snail promoter is also the target of c-Myc. Next, we determined whether c-Myc interacts with Snail using a Co-IP assay, and found that Snail was also be pulled down by c-Myc (Fig. 4E).

c-Myc promotes EMT via TGF-β/Snail signaling pathway and VM via VE-cadherin. A and B Western Blotting assay and Gray value analysis showed TGF-β promoted c-Myc and Snail protein expression; C showed TGF-β promotes c-Myc gene expression; D showed c-Myc promotes Snail gene expression; E showed there is interaction between c-Myc and Snail; F, G and H showed c-Myc promotes VE-cadherin protein and gene expression. (*P < 0.05)
c-Myc promotes VM by upregulation of VE-cadherin expression
To further delineate the signaling pathway of c-Myc in regulation of VM, we used western blot and luciferase reporter gene assays.
Western blot results showed that c-Myc overexpression can increased VE-cadherin protein expression (Fig. 4F and G). A375, MUM-2B cells were cotransfected with a VE-cadherin promoter-luciferase construct and c-Myc-cDNA in an expression vector. Luciferase reporter gene assay results showed that c-Myc overexpression enhanced VE-cadherin promoter activity (Fig. 4H). These results indicate that the VE-cadherin promoter is one of the targets of c-Myc.
c-Myc overexpression promotes LPPCN by the upregulating Bax expression
c-Myc overexpression sensitizes cells to CoCl2 treatment and hypoxia (1 % O2), leading to cell apoptosis by upregulating Bax expression
To assess the pro-apoptotic role of c-Myc in tumor cells in vivo, we treated melanoma cells with hypoxia-mimetic agent CoCl2 (200 μM, 12 h) [23, 24] and then used an AnnexinV-FITC/PI Apoptosis Detection Kit and flow cytometry to detect cell death.
Without CoCl2 treatments, there were no differences in the apoptotic index of high versus low-c-Myc expressing cells. However, the rate of apoptosis in MUM-2B cells transfected with sh-c-Myc was lower than that in controls, when the cells were treated with CoCl2. The expected opposing relationship was observed when c-Myc overexpressing MUM-2C cells were compared with controls (Fig. 5A and B, P < 0.05).

A Melanoma cell lines treated with or without CoCl2 (200 μM, 12 h) were stained with PI and Annexin V; B Early and late apoptotic cells are shown in the right quadrant. There were significant differences in cells with CoCl2 treatment; C and D Western blotting and gray analysis showed Bax expression was significantly upregulated in cells with CoCl2 treatment; E showed that it is BAX siRNA not BCL2 siRNA weakened the differences in the number of apoptotic cells under hypoxic conditions (1%O2).*P < 0.05, ** P < 0.001
To explore the roles of c-Myc in mediating apoptosis, we evaluated Bax protein expression in melanoma cells treated with or without 200 μM CoCl2 for 12 h. Our data demonstrated that Bax protein expressions was linked to c-Myc expression levels. c-Myc knock-down in MUM-2B cells downregulated Bax expression, whereas c-Myc overexpression in MUM-2C cells upregulated Bax expression only with CoCl2 treatment (Fig. 5C and D).
To further analyze the roles of Bax in the process that c-Myc sensitized cells to hypoxia (1 % O2) by c-Myc, we silenced Bax or Bcl2 expression by siRNAs. Under hypoxic conditions, the differences in apoptosis rates were similar to cells treated with CoCl2. Moreover, silencing the expression of Bax but not Bcl2 reduced these differences. (Fig. 5E, P < 0.05).
c-Myc overexpression plays different roles in cells treated with different levles of CoCl2 by regulating the ratio of Bcl2/Bax
To explore the role of c-Myc in cells treated with CoCl2, we conducted experiments to modulate the concentration and time in which cells were exposed to CoCl2 in vitro. We then evaluated the expression of apoptosis-related genes by RT-PCR and multi-factor ANOVA.
Bax expression was down-regulated by c-Myc knock-down expression, while the ratio of Bcl2/Bax was concomitantly reduced (Fig. 6A-a). The converse results were observed following the c-Myc overexpression in MUM-2C cells (Fig. 6A-b). More specifically, the results suggested that both Bax expression and the Ratio of Bcl2/Bax were related to HIF-1α expression, and this relationship was modulated by a concentration and time-dependent manner, relating to the exposure to CoCl2. Notably, Bax response to CoCl2 treatment was most prominent at 12 h (Fig. 6A-c), while the ratio of Bcl2/Bax increased over time. The ratio of Bcl2/Bax increased at 100 μM CoCl2 treatment and decreased upon exposure to higher concentrations of CoCl2 (200, 300, 400 and 800 μM) (Fig. 6A-d). Detailed results of multi-factor ANOVA are shown in Supplementary Table S2.

A a and b showed c-Myc high expression upregulated Bax expression and increased the ratio of Bcl2/Bax in cells with CoCl2 treatment. c showed the ratio of Bcl2/Bax was at its lowest level at this time point 12 h; d the ratio of Bcl2/Bax peaked at concentration of 100 μM of CoCl2, decreased with increasing of levels of CoCl2; B Western Blotting assay showed the similar results to RT-PCR. *P < 0.05, **P < 0.001, n = 3
To examine the roles of hypoxia-inducible factor 1α (HIF-1α) and c-Myc, we detected the expression of these proteins by western blotting. The results were shown in Fig. 6B. High HIF-1α expression inhibited the expression of c-Myc and Bcl2, and enhanced Bax expression. Taken together, these results suggest that c-Myc overexpression can cause cell death via the c-Myc/Bax signaling pathway by decreasing the ratio of Bcl2/Bax.
Examing the effects of c-myc using a murine xenograft model
High c-Myc expression promotes tumor growth in vivo
The effects of c-Myc on tumor growth in nude mice were reflected by growth curves. The growth curves showed that murine xenografted melanoma tumors with high c-Myc expression developed faster than those with low c-Myc expression (p < 0.05, Fig. 7A). The weight of tumors with high c-Myc expressions was higher than that of tumors with low c-Myc expression (p < 0.05, Fig. 7B).

The roles of c-Myc in melanoma murine xenograft tumor. A and B Tumor growth curve and tumor weight showed high c-Myc expression promotes tumor growth; C tumors with high c-Myc expression have more VM tubes; D high c-Myc expression increased the number of LPPCN cells; E LPPCN cells are darkly stained and distributed in patterns of lines and/or networks among the environmental tumor cells with no inflammation cells. (×400, bar 50um); F EDV with positive PAS staining and Endomucin expression (the black arrow) and VM channels lined with tumor cells and PAS-positive substances and negative Endomucin expression, and red cells remained visible at the center of the VM channels (the red arrow) (×400, bar 50um); G H&E staining showed lung metastases of melanoma. H showed that tumors with higher c-Myc expression had loss of E-cadherin proteins and elevated Snail, VE-cadherin expression. HIF-1α, Bcl2 Bax expression exhibited in linear pattern (×400, bar 50um). I A model and the molecular mechanisms of c-Myc on the formation of VM and LPPCN. a Under mild hypoxia, the ratio of Bcl2/Bax was up-regulated and c-Myc triggered the epithelial-mesenchymal transition (EMT) and led cell survive; b Under severe hypoxia, the ratio of Bcl2/Bax was down-regulated and c-Myc sensitized tumor cells to oxygen deprivation and led cell death; c Tumor cells formed VM tube following the LPPCN cells disappeared, EMT occurrence and red cell got into. *P < 0.05, n = 10
c-Myc overexpression increases the formation of VM and LPPCN in vivo
To evaluate the relationship between c-Myc expression and VM in vivo, we performed Endomucin/PAS double staining. VM (Fig. 7F, red arrow) formation increased following c-Myc overexpression in MUM-2C xenograted tumor and decreased upon gene silencing of c-Myc in MUM-2B xenograted tumor (Fig. 7C, p < 0.05).
To evaluate the correlation between c-Myc expression and the formation of LPPCN, we performed H&E staining. Notably, there were significantly more LPPCN cells in MUM-2B tumors as compared to MUM-2B-sh-c-Myc tumors. Moreover, the number of LPPCN cells in the MUM-2C-vector groups was significantly lower than that in MUM-2C-c-Myc tumors (Fig. 7D and E, p < 0.001).
High c-Myc expression correlates with tumor metastasis in vivo
To detect the role of c-Myc in the invasion and metastasis of tumor, we perfomed metastasis assay in melanoma xenograft model. When the tumor was injected into the back of the mice, only the MUM-2B-vector tumors (1/6) has lung metastases (Fig. 7G). For colonization assay, tumors in lung were observed in 83.3 % (5/6) of MUM-2B-vector xenografted tumor (VS 0/6 of MUM-2B-sh-c-Myc tumors) and 1/6 of MUM-2C-c-Myc tumors (VS 0/6 of MUM-2C-vector tumor) following tail vein injection of tumor cells. The results indicated that c-Myc may play important roles in the tumor metastasis.
High c-Myc expression promotes VM by upregulatipn of EMT and LPPCN related protein expression in vivo
To further clarify the role of c-Myc in the formation of VM and LPPCN in vivo, we evaluated the expression of EMT and LPPCN related proteins by immuohistochemistry (Fig. 7H). Consistent with our results, loss of E-cadherin, increased Snail expression, and VE-cadherin expression was observed in high c-Myc expressing tumors, while the converse relationship was observed upon c-Myc shRNA-mediated silencing. LPPCN cells specifically showed to be Bax positive. Additionally, HIF-1α, Bcl2, and Bax expression was arranged in lines or in a net. We deduced that the formation of VM was related to c-Myc over-expression, and that LPPCN was induced by c-Myc-related cell death.
Discussion
It is important to note that, it is the metastasis, rather than the primary tumor, that ultimately leads to patient death, caused by metastatic growth that compromises the function of vital organs including, most notably, the lymph nodes, liver, lungs and brain.
The proto-oncogene c-Myc encodes a nuclear phosphoprotein transcription factor that plays an integral role in a variety of cellular processes [25, 26]. Interestingly, studies have now shown that c-Myc induction of programmed cell death may contribute to carcinogenesis [27]. However, the shift in the paradoxical properties of c-Myc in which c-Myc induces a pro-apoptotic effect versus EMT and results in metastasis of melanoma remains unclear.
In this study, we demonstrated that c-Myc accelerates metastasis by promoting the formation of VM via two mechanisms. First, c-Myc upregulates Snail expression to promote EMT via the TGF-β/Snail/E-cadherin signal pathway. Second, c-Myc upregulates Bax expression, resulting in a decrease in the ratio of Bcl2 and Bax under severe hypoxic conditions and leading to cell death, thereby promoting the formation of LPPCN.
Across our cohort, nearly 50 present of melanoma cases demonstrated high c-Myc protein expression. High c-Myc expression tumors are associated with a poor prognosis and an elevated incidence of metastasis. A study has previously demonstrated that VM significantly correlates with metastasis and a poor prognosis [9]. VM was observed in almost 60 % of melanomas with high c-Myc expression. In vivo and in vitro experiments highlighted the role of c-Myc in VM channel formation. Specifically, tumors with high c-Myc protein expression formed VM channels, whereas low c-Myc-expressing tumors did not show the same effect. Our results suggest the involvement of c-Myc in the formation of VM and promotion of disease progression.
Kang et al. demonstrated that overexpression of c-Myc mediates EMT, which plays an important roles in the formation of VM, through loss of functional E-cadherin [11, 28–30]. However, the manner by which c-Myc promotes EMT remains to be investigated in melanoma. Our results showed that exogenous c-Myc overexpression or high expression leads to increased expression of Snail and Vimentin and loss expression of E-cadherin in melanoma cells. Notably, a similar signaling axis was observed in our xenograft models. TGF-β increased the protein and gene expression of c-Myc and Snail in melanoma cells. Moreover, we demonstrated that c-Myc overexpression enhanced motility and invasiveness of A375 and MUM-2C cells, whereas c-Myc knockdown had the opposite effect. As such, we hypothesize that c-Myc may promote EMT via the TGF-β/Snail/E-cadherin signaling pathway, leading to the formation of VM.
Previous studies have demonstrated that c-Myc/Max heterodimers bind to canonical E-box elements in the promoter region of BAX [31]. Substantial evidence supports the notion that c-Myc sensitizs cells to apoptosis by altering the balance of pro- and anti-apoptotic factors, priming the cells for death when such conditions are appropriate [32]. A study has identified c-Myc up-regulated p53 to activate Bax and promotes the release of holo-cytochrome C from mitochondria [33]. Bcl-2/Bcl-xL proteins suppress holo-cytochrome C release by inhibition of c-Myc-induced apoptosis and promotion of c-Myc oncogenicity [34]. As such, the ratio of Bcl2 and Bax determines whether cells survive or undergo apoptosis. Our results demonstrate a relationship bwtween c-Myc-induced apoptosis and the formation of LPPCN via the competition between Bcl2 and Bax.
Nearly 80 % (54/69) of melanomas with high c-Myc expression were shown to contain LPPCN. In support of these findings, we demonstrated that c-Myc suppression in murine xenograft models reduced the incidence of LPPCN. In vitro, flow cytometry data suggested that high c-Myc expression sensitized tumor cells to hypoxia and hypoxia-mimetic agent CoCl2, and leads to apoptosis. Western blotting and RT-PCR analysis supported this hypothesis and highlighted the hypoxic-specific relationship between c-Myc and Bax expression. Our results indicate that c-Myc overexpression induced up-regulation of Bax and decreases the ratio of Bcl2/Bax when the cells was treated with elevated levels of CoCl2. c-Myc may regulate HIF-1α to enhance tumor angiogenesis [35]. Furthermore, HIF-1α may inhibit DNA damage reduced by c-Myc [36]. A study has found that the HIF-1α-c-Myc pathway promotes cell survival only in apoptosis-deficient malignant cells [37]. Another study has suggested that c-Myc sensitization to oxygen deprivation-induced cell death depends on Bax/Bak [38]. LPPCN was recently reported in highly aggressive melanoma by Zhang et al. In their study, LPPCN cells were shown to be positive for Bax by immunohistochemistry, which was verified again in this study, realing a significant relationship between Bax and LPPCN [13].
We suggest that when the blood supply of tumor is occluded and results in severe hypoxia, elevated expression of c-Myc decreases the ratio of Bcl2/Bax and activates apoptotic pathways in tumor cells. Cell death is distributed in specific patterns of lines and networks, in a LPPCN-like manner. We conclude that the c-Myc-dependent apoptotic response via Bax-dependent signaling represents important elements in the formation of LPPCN. A study has previously suggested that the formation of LPPCN provides a basis for the spatial organization of VM and EDV structures [22]. We believe that c-Myc promotes the formation of VM by regulation of LPPCN.
The center of a bulk tumor is under hypoxic or even anoxic conditions [39, 40]. In a hypoxic microenvironment, c-Myc promotes EMT in tuomor cells, thereby inducing cell survival, metastasis and the formation of VM. However, upon acute or perfusion-limited hypoxia (such as temporal fluctuations in blood perfusion) or in instances of chronic hypoxia, high expression of c-Myc induces cells death, driving the formation of LPPCN. In this manner, c-Myc drives the creation of spatial structure necessary for VM. Taken together, we suggest that c-Myc positive cells that have undergone EMT form VM in the space created through the disappearance of LPPCN cells and resulting in tumor metastasis. All of these processes are driven by c-Myc, as shown in Fig. 7I.
The definition of LPPCN was, until now, solely a morphological description while the molecular characteristics have thus far remained unclear. We can no longer categorize LPPCN as the kind of cell death described by the Nomenclature Committee on Cell Death in 2012 [41]. Our study provides a possible mechanism of LPPCN. Additionally, LPPCN is an active gene-regulated process, that plays an important role in cancer development and progression. Therefore, greater emphasis on characterizing the molecular features of LPPCN could improve our understanding of programmed cell death.
Our study suggest that over expressed c-Myc could be regarded as an important parameter in the malignant degree and poor prognosis of melanoma and highlights the pleiotropic nature of oncogenes in the development of cancers. Considering the central role of c-Myc in the pathobiology of numerous cancers, including melanoma, our results suggest that improved understanding and characterization of this gene could lead to the development of novel, targeted therapies.
References
Caini S, Boniol M, Botteri E, Tosti G, Bazolli B, Russell-Edu W, Giusti F, Testori A, Gandini S (2014) The risk of developing a second primary cancer in melanoma patients: a comprehensive review of the literature and meta-analysis. J Dermatol Sci 75:3–9
Ji H, Cao R, Yang Y, Zhang Y, Iwamoto H, Lim S, Nakamura M, Andersson P, Wang J, Sun Y, et al (2014) TNFR1 mediates TNF-α-induced tumour lymphangiogenesis and metastasis by modulating VEGF-C-VEGFR3 signalling. Nat Commun 5:4944
Hedlund EM, Yang X, Zhang Y, Yang Y, Shibuya M, Zhong W, Sun B, Liu Y, Hosaka K, Cao Y (2013) Tumor cell-derived placental growth factor sensitizes antiangiogenic and antitumor effects of anti-VEGF drugs. Proc Natl Acad Sci U S A 110:654–659
Folkman J, Ryeom S (2005) Is oncogene addiction angiogenesis-dependent? Cold Spring Harb Symp Quant Biol 70:389–397
Cao Y (2016) Future options of anti-angiogenic cancer therapy. Chin J Cancer 35:21
Cao Y (2014) VEGF-targeted cancer therapeutics-paradoxical effects in endocrine organs. Nat Rev Endocrinol 10:530–539
Cao Y, Arbiser J, D’Amato RJ, D’Amore PA, Ingber DE, Kerbel R, Klagsbrun M, Lim S, Moses MA, Zetter B, et al (2011) Forty-year journey of angiogenesis translational research. Sci Transl Med 3:114rv113
Maniotis AJ, Folberg R, Hess A, Seftor EA, Gardner LM, Pe’er J, Trent JM, Meltzer PS, Hendrix MJ (1999) Vascular channel formation by human melanoma cells in vivo and in vitro: vasculogenic mimicry. Am J Pathol 155:739–752
Cao ZF, Bao MM, Miele L, Sarkar FH, Wang ZW, Zhou QS (2013) Tumour vasculogenic mimicry is associated with poor prognosis of human cancer patients: a systemic review and meta-analysis. Eur J Cancer 49:3914–3923
Meng J, Sun B, Zhao X, Zhang D, Zhao X, Gu Q, Dong X, Zhao N, Liu P, Liu Y (2014) Doxycycline as an inhibitor of the epithelial-to-mesenchymal transition and vasculogenic mimicry in hepatocellular carcinoma. Mol Cancer Ther 13:3107–3122
Sun T, Zhao N, Zhao XL, Gu Q, Zhang SW, Che N, Wang XH, Du J, Liu YX, Sun BC (2010) Expression and functional significance of Twist1 in hepatocellular carcinoma: its role in vasculogenic mimicry. Hepatology 51:545–556
Kalluri R, Weinberg RA (2009) The basics of epithelial-mesenchymal transition. J Clin Invest 119:1420–1428
Zhang S, Li M, Zhang D, Xu S, Wang X, Liu Z, Zhao X, Sun B (2009) Hypoxia influences linearly patterned programmed cell necrosis and tumor blood supply patterns formation in melanoma. Lab Investig 89:575–586
Hoffman B, Liebermann DA (2008) Apoptotic signaling by c-MYC. Oncogene 27:6462–6472
Evan G, Littlewood T (1998) A matter of life and cell death. Science 281:1317–1322
Blum D, Hao H, McCarthy M, Reproducibility Project: Cancer B (2015) Registered report: transcriptional amplification in tumor cells with elevated c-Myc. Elife 4.e04024. doi:10.7554/eLife.04024
Meyer N, Penn LZ (2008) Reflecting on 25 years with MYC. Nat Rev Cancer 8:976–990
Kraehn GM, Utikal J, Udart M, Greulich KM, Bezold G, Kaskel P, Leiter U, Peter RU (2001) Extra c-myc oncogene copies in high risk cutaneous malignant melanoma and melanoma metastases. Br J Cancer 84:72–79
Zhao N, Sun BC, Zhao XL, Liu ZY, Sun T, Qiu ZQ, Gu Q, Che N, Dong XY (2012) Coexpression of Bcl-2 with epithelial-mesenchymal transition regulators is a prognostic indicator in hepatocellular carcinoma. Med Oncol 29:2780–2792
Lin X, Sun B, Zhu D, Zhao X, Sun R, Zhang Y, Zhang D, Dong X, Gu Q, Li Y, et al (2016) Notch4+ cancer stem-like cells promote the metastatic and invasive ability of melanoma. Cancer Sci. doi:10.1111/cas.12978
Workman P, Aboagye EO, Balkwill F, Balmain A, Bruder G, Chaplin DJ, Double JA, Everitt J, Farningham DA, Glennie MJ, et al (2010) Guidelines for the welfare and use of animals in cancer research. Br J Cancer 102:1555–1577
Han C, Sun BU, Wang W, Cai WJ, Lou D, Sun Y, Zhao X (2010) A pilot study on morphology and the mechanism involved in linearly patterned programmed cell necrosis in melanoma. Oncol Lett 1:821–826
Garayoa M, Martínez A, Lee S, Pío R, An WG, Neckers L, Trepel J, Montuenga LM, Ryan H, Johnson R, et al (2000) Hypoxia-inducible factor-1 (HIF-1) up-regulates Adrenomedullin expression in human tumor cell lines during oxygen deprivation: a possible promotion mechanism of carcinogenesis. Mol Endocrinol 14:848–862
Cascio S, Bartella V, Auriemma A, Johannes GJ, Russo A, Giordano A, Surmacz E (2008) Mechanism of leptin expression in breast cancer cells: role of hypoxia-inducible factor-1alpha. Oncogene 27:540–547
Dang CV (2012) MYC on the path to cancer. Cell 149:22–35
Rahl PB, Young RA (2014) MYC and transcription elongation. Cold Spring Harb Perspect Med 4:a020990
Wang C, Tai Y, Lisanti MP, Liao DJ (2014) C-myc induction of programmed cell death may contribute to carcinogenesis. Cancer Biology & Therapy 11:615–626
Chen D, Huang J, Zhang K, Pan B, Chen J, De W, Wang R, Chen L (2014) MicroRNA-451 induces epithelial-mesenchymal transition in docetaxel-resistant lung adenocarcinoma cells by targeting proto-oncogene c-myc. Eur J Cancer 50:3050–3067
Oloumi A, McPhee T, Dedhar S (2004) Regulation of E-cadherin expression and beta-catenin/Tcf transcriptional activity by the integrin-linked kinase. Biochim Biophys Acta 1691:1–15
Sun T, Sun BC, Zhao XL, Zhao N, Dong XY, Che N, Yao Z, Ma YM, Gu Q, Zong WK, et al (2011) Promotion of tumor cell metastasis and vasculogenic mimicry by way of transcription coactivation by Bcl-2 and Twist1: a study of hepatocellular carcinoma. Hepatology 54:1690–1706
Mitchell KO, Ricci MS, Miyashita T, Dicker DT, Jin Z, Reed JC, El-Deiry WS (2000) Bax is a transcriptional target and mediator of c-myc-induced apoptosis. Cancer Res 60:6318–6325
Pelengaris S, Khan M, Evan GI (2002) Suppression of myc-induced apoptosis in beta cells exposes multiple oncogenic properties of myc and triggers carcinogenic progression. Cell 109:321–334
Westphal D, Dewson G, Czabotar PE, Kluck RM (2011) Molecular biology of Bax and Bak activation and action. Biochim Biophys Acta 1813:521–531
Wagner AJ, Small MB, Hay N (1993) Myc-mediated apoptosis is blocked by ectopic expression of Bcl-2. Mol Cell Biol 13:2432–2440
Chen C, Cai S, Wang G, Cao X, Yang X, Luo X, Feng Y, Hu J (2013) C-myc enhances colon cancer cell-mediated angiogenesis through the regulation of HIF-1alpha. Biochem Biophys Res Commun 430:505–511
Huang LE, Bindra RS, Glazer PM, Harris AL (2007) Hypoxia-induced genetic instability--a calculated mechanism underlying tumor progression. J Mol Med (Berl) 85:139–148
Fer N, Melillo G (2011) The HIF-1alpha-c-myc pathway and tumorigenesis: evading the apoptotic gate-keeper. Cell Cycle 10:3228
Brunelle JK, Santore MT, Budinger GR, Tang Y, Barrett TA, Zong WX, Kandel E, Keith B, Simon MC, Thompson CB, et al (2004) C-myc sensitization to oxygen deprivation-induced cell death is dependent on Bax/Bak, but is independent of p53 and hypoxia-inducible factor-1. J Biol Chem 279:4305–4312
Li S, Zhang J, Yang H, Wu C, Dang X, Liu Y (2015) Copper depletion inhibits CoCl2-induced aggressive phenotype of MCF-7 cells via downregulation of HIF-1 and inhibition of snail/twist-mediated epithelial-mesenchymal transition. Sci Rep 5:12410
Fardin P, Barla A, Mosci S, Rosasco L, Verri A, Versteeg R, Caron HN, Molenaar JJ, Ora I, Eva A, et al (2010) A biology-driven approach identifies the hypoxia gene signature as a predictor of the outcome of neuroblastoma patients. Mol Cancer 9:185
Galluzzi L, Vitale I, Abrams JM, Alnemri ES, Baehrecke EH, Blagosklonny MV, Dawson TM, Dawson VL, El-Deiry WS, Fulda S, et al (2012) Molecular definitions of cell death subroutines: recommendations of the nomenclature committee on cell death 2012. Cell Death Differ 19:107–120
Acknowledgment
This study was partly supported by the follow grants:
1) Key project of the National Natural Science Foundation of China (NO. 81230050);
2) The National Natural Science Foundation of China (NO. 81172046);
3) The National Natural Science Foundation of China (NO. 81173091);
4) The National Basic Research Program of China (973 Program, NO. 2010CB529403).
Author contribution statement
B.C.S. conceived the study, B.C.S., R.S. and X.L. designed experiments and wrote the manuscript. X.L., X.L.Z., X.M.Z., Q.G. and X.Y.D. performed experiments. D.W.Z. and D.F.Z. analyzed the data, Y.H.Z., R.S. and Y.L.L. collected the human pathological specimen of melanoma. All authors revised the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Disclosure
The authors declare that they have no conflict of interests.
Additional information
Xian Lin and Ran Sun contributed equally to this work.
Electronic supplementary material
ESM 1
(PDF 338 kb)
Rights and permissions
About this article

Cite this article
Lin, X., Sun, R., Zhao, X. et al. C-myc overexpression drives melanoma metastasis by promoting vasculogenic mimicry via c-myc/snail/Bax signaling. J Mol Med 95, 53–67 (2017). https://doi.org/10.1007/s00109-016-1452-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00109-016-1452-x