
Abstract
The existence of transport networks is vital for leaf-cutting ant foraging but may generate overcrowding, reducing the input of food into the nest. We studied how ants turn at branching points, a problematic sector for ant traffic, in leaf-cutting ant species that vary in ant flow and trail design. If the walking of turning ants reduces the chance of collisions, we expected that (a) ants that keep the same lane while turning suffer less collisions than those who change lane, (b) the behavior of keeping the same lane will be higher than expected by chance, and (c) lane fidelity of turning ants should increase as ant flow increases. We recorded the turning movements of 1355 individuals in trail bifurcations from 25 ant nests. Each ant was categorized according to the fidelity to its traffic lane while turning, the number of collisions, and the ant flow at the moment of turning. Ants faithful to their lane had fewer collisions than ants unfaithful to their lane when turning, but only in the two Atta species. Lane fidelity when turning was the most frequent behavior in all species, but this behavior did not increase with increments in ant flow. Leaf-cutting ants appear to follow simple rules to reduce overcrowding: keep walking on the same lane when turning. We discussed the influence of ant flow and trail design on this behavior and the dual role of collisions (information exchange and traffic delay) in trail sectors conflicting for traffic circulation.
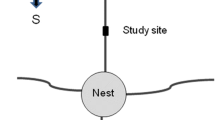
Graphical Abstract
Bifurcations may be a conflictive sector for smooth traffic in leaf-cutting ants. We showed that ants appear to follow simple rules to reduce overcrowding at theses trail sectors: keep walking on the same traffic lane when turning.

Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The existence of transport networks is considered vital to the success of leaf-cutting ant foraging. Trunk trails guide foragers to plant sources and facilitate resource monitoring, worker locomotion, information transfer, and transport efficiency (Shepherd 1982; Rockwood and Hubbell 1987; Wirth et al. 2003; Kost et al. 2005; Farji-Brener et al. 2010). However, the use of trail systems also entails costs. First, the maintenance of this large and permanent trail system free of debris requires time and energy (Lugo et al. 1973; Shepherd 1982; Bochynek et al. 2017). For example, in tropical wet forests leaf-cutting ants invest an average of 11,000 ant days of work and the energy equivalent of 8000 leaf burdens to maintain a trail system free of debris (Howard 2001). Moreover, the probability of litter fall affects trail branching (Farji-Brener et al. 2015). Second, trail design may impose physical constraints on the foraging rate: food delivery declines dramatically as trail length increases (Bruce and Burd 2012) and the vegetation of trail margins retards the speed of loaded ants (Farji-Brener et al. 2012; Bochynek et al. 2017). Finally, the existence of transport networks imposes adequate traffic rules to avoid overcrowding. It is expected that natural selection selected for organizational rules that can maximize the traffic flow in order to ensure a high rate of food return to the nest (Burd et al. 2002; Burd 2006; Dussutour et al. 2009). A better knowledge of the behaviors that reduce traffic congestions is essential to understand the success of the trunk trail system despite its costs.
Branching points along foraging trails are problematic sectors for ant traffic. A typical leaf-cutting ant trail system consists of a persistent, dendritic network radiating out to food sources, with up to 50 trail bifurcations (Kost et al. 2005). At these branching points, ingoing ants enter to the main trail from the bifurcation at the same time that outgoing ants split between the bifurcation and the main trail. This convergence of ingoing and outgoing ants in a relatively small area increases the probability of collisions between workers moving in opposite directions (Elizalde and Farji-Brener 2012). Each collision induces a short delay in the progression of individuals, which in turn, when summed over the thousands of collisions that occur along a foraging trail, decreases the rate of food return to the nest (Burd et al. 2002; Burd and Aranwela 2003). Therefore, ants should exhibit behaviors to reduce overcrowding (Dussutour et al. 2004; Fourcassié et al. 2010).
Leaf-cutting ants are ideal organisms to study the behavior of ant walking at branching points. First, they are a good example of absence of lane segregation between ingoing and outgoing ants, which results in high rate of ant collisions (Burd et al. 2002; Burd and Aranwela 2003). Second, the role of these head-on encounters is ambivalent. As discussed earlier they can retard the speed of foraging ants, but also may promote information transfer between individuals, increasing foraging efficiency and the chance to finding new food sources (Dussutour et al. 2004, 2007; Farji-Brener et al. 2010; Bouchebti et al. 2015). Finally, leaf-cutting ant species differ greatly in their population density, foraging flow and trail system. For example, colonies from the genus Atta can hold up to 7 million of workers, showing higher foraging flow and wider trails than the less dense Acromyrmex species, which show lower foraging flow and a less sophisticated trunk trail system (Hölldobler and Wilson 2011). In sum, these characteristics make leaf-cutting ants ideal organisms to study how ants behave at branching points. Here we analyze the walking of leaf-cutting ants at trail bifurcations in three species that differ in flow density and trail design. If, as proposed, turning ants walk in order to reduce the chance of collisions, we expect the following: (a) ants that keep the same traffic lane while turning suffer less collisions than those who change traffic lane, (b) the behavior of keeping the same traffic lane (lane fidelity) will be higher than expected by chance, and (c) lane fidelity of turning ants should increase as ant flow increases.
Materials and methods
Leaf-cutting ant species
We worked with Atta cephalotes, Atta vollenweideri, and Acromyrmex lobicornis. Both Atta species show high ant flow along foraging trails, a large cleared trunk trail system but differ in the habitat where they live and the kind of plants they cut. Atta cephalotes occurs in tropical wet forest and mainly cuts dicots, whereas A. vollenweideri inhabits subtropical savannas and cuts only herbs and forbs (Farji-Brener and Ruggiero 1994; Hölldobler and Wilson 2011). On the other hand, A. lobicornis occurs mainly in temperate deserts and scrublands cutting dicots and herbs, and shows lower worker density and foraging rates, and a less developed trunk trail system than Atta species (Farji-Brener and Ruggiero 1994; Farji-Brener and Tadey 2017; Fig. 1).

Trunk trail and bifurcations in leaf-cutting ants Atta cephalotes nests at La Selva Biological Station, Costa Rica (a), Atta vollenweideri in Estancia El Socorro, Corrientes, Argentina (b), and Acromyrmex lobicornis near the city of Bariloche, Patagonia, Argentina (c). Photo credit: AGFB
Study sites
Sampling on Atta cephalotes nests was conducted at La Selva Biological Station of the Organization for Tropical Studies (10°26′N, 8°59′W) in the Atlantic lowlands of Costa Rica. The area is a lowland wet forest that receives a mean annual rainfall of 4 m. A. cephalotes is the most common leaf-cutting ant species in La Selva, and shows an extensive and permanent trail system (Farji-Brener 2001; Kost et al. 2005; Farji-Brener et al. 2015). Sampling on Atta vollenweideri nests was conducted at two sites in the Province of Corrientes, north-east Argentina. Both sites are separated by approximately 100 km. Site one is located at “Reserva Rincón del Socorro” (28°32′S, 57°10′W) on the margin of Iberá wetlands. The reserve is a mix of temporarily flooded open pasturelands, wooded savannas, hydrophilic forests, and wetlands. Site two is located at ‘Estancia La Paz’ (29°20′S, 58°26′W) near the town of Mercedes. This site is mostly characterized by open pasturelands with patches of thorny deciduous scrubland forest dominated by Prosopis affinis. Sampling on Acromyrmex lobicornis nests was conducted on the eastern border of the Nahuel Huapi National Park, located in Patagonia, Argentina (41°S, 72°W). This area comprises herbaceous/shrub steppe vegetation. The weather in this region is usually dry and cold, with an average precipitation of 600 mm and a mean annual temperature of 8 °C.
Methods
The sampling procedure was the same for the three leaf-cutting ant species. We randomly selected adult nests with cleared trunk trails. In each nest we selected between 1 and 2 trail sectors that include a well-defined bifurcation to video record the turning walking of ingoing and outgoing ants (Fig. 1). Trails selected were 14 ± 2, 12 ± 2, and 7 ± 1 cm of width (mean ± SE) for Atta cephalotes, Atta vollenweideri, and Acromyrmex lobicornis, respectively. We selected the first branching points from the nest entrance to ensure a potential high ant flow, which varied among species, by mean, between 0.035 and 0.028 ants/cm/s. To avoid off-axis distortion, we placed a ruler parallel to ant walking direction and filmed from above and perpendicular to the trail. We filmed this trail sectors during ~10 min at different times of the day to include a wide range of ant flow conditions. Each video was carefully observed in the lab and a number between 10 and 30 turning ants (ingoing and outgoing) was randomly selected. The recorded/selected ingoing ants were always loaded. On the video, we divided the sampled trail and the bifurcation in three imaginary traffic lanes of equal width parallel to the trail border (left, central, and right) and (1) we categorized sampled ants according to the fidelity to the traffic lane while turning (faithful and unfaithful, Fig. 2), (2) we counted the number of collisions of each sampled ant with workers moving in the opposite direction, and (3) we estimated ant flow at that moment of turning as the number of ants/20 s/cm2 of trail. We sampled a total of 1355 ants (656 loaded and 699 unloaded) from 25 ant nests (6 from Atta vollenweideri, 13 from Atta cephalotes, and 6 from Acromyrmex lobicornis). The number of ants sampled per species was 230 ants (106 loaded and 124 unloaded) from 8 bifurcations in A. vollenweideri, 925 ants (450 loaded and 475 unloaded) from 22 bifurcations in A. cephalotes, and 200 ants (100 loaded and 100 unloaded) from 10 bifurcations in A. lobicornis.

Diagram of how we characterized turning ants according to whether they kept walking on the same lane (lane fidelity) or switched lanes (lane infidelity) while turning, for ingoing (above) and outgoing (below) ants. In the video record, we divided the trail and bifurcation in three imaginary sections of similar width parallel to the trail borders. Unfaithful ants may cross one or two traffic lanes when turning (central and right side of the diagram), potentially increasing the chance of collisions with ants walking in opposite direction
Statistical analyses
First, we confirmed whether ants faithful to their traffic lane had fewer collisions than unfaithful ants by a three-factor mixed ANCOVA. The number of collisions per second was the response variable, the ant walking while turning (faithful or unfaithful to its traffic lane) and ant direction (ingoing and outgoing) were considered as fixed factors, and bifurcation was considered as a random factor. Ant flow was included as a co-variable. ANCOVA assumptions were checked and data were log-transformed when necessary. Second, we compared the frequency of faithful and unfaithful walking when turning (as defined above) using Chi-square tests. Since we predefined three traffic lanes and two possible behaviors (to be or not to be faithful), the expected probabilities were estimated as follows. Ants in the central lane do indeed have two opportunities to be unfaithful during a turn, veering into either the left lane or the right lane. But ants in the left lane can only remain faithful or veer to the right. Once they cross into the central lane, they are unfaithful; whether they continue to veer further into the right lane does not change their status as unfaithful turners. So in that sense, these outer-lane ants have only two opportunities to continue in the lane or veer right. The ants in the outer right-hand lane are symmetrical. Now if we assume that one-third of the total traffic flow occurs in each lane, the probability of observing faithful turning becomes 1/2 for the 1/3 of ants in the left lane (=1/6 overall); similarly, 1/6 for the ants in the right lane; and 1/3 for the 1/3 of ants in the central lane (=1/9 overall). The total expected probability of observing faithful turning would then be 1/6 + 1/6 + 1/9 = 4/9, and the expected probability for unfaithful turning would be 5/9. Finally, we analyzed whether the probability of lane faithful (that should decrease collision rates) increased with ant flow using logistic mixed regressions. Bifurcation was included as a random variable. All the analyses were performed for each leaf-cutting ant species separately using R statistical software (R Development Core Team 2015).
Results
The collision rate depended on ant direction (outgoing and ingoing) and ant walking category (lane faithful and unfaithful) for Atta species but not for Acromyrmex lobicornis (ESM1). First, outgoing (unladen) ants collided more with ants walking in opposite direction than ingoing (laden) ants (0.19 ± 0.02 vs. 0.09 ± 0.002 head-on encounters/s for Atta cephalotes; 0.17 ± 0.03 vs. 0.10 ± 0.05 for Atta vollenweideri, all P < 0.01, mean ± SE). Second, ants faithful to their traffic lane had fewer collisions than ants unfaithful to their traffic lane while turning (0.12 ± 0.02 vs. 0.16 ± 0.02 for Atta cephalotes; 0.10 ± 0.03 vs. 0.17 ± 0.03 for A. vollenweideri, all P < 0.01). The collision rate in Atta was also affected by the sampled bifurcation (i.e., by non-measured, intrinsic characteristics of each colony) and slightly by ant flow (ESM 1). On the other hand, the collision rate in A. lobicornis was only affected by the sampled bifurcation. Ant direction and ant walking category did not affect the number of head-on encounters with ants walking in opposite direction (0.055 ± 0.015 vs. 0.050 ± 0.015 ongoing vs. ingoing ants; 0.051 ± 0.05 vs. 0.051 ± 0.05 faithful vs. unfaithful ants, respectively, mean ± SE of head-on encounters/sec, all P >0.40; Fig. 3).

Ant collision rate (number of head-on encounters/s) according to the walking condition (faithful or unfaithful to their traffic lane while turning) for Atta cephalotes (a), Atta vollenweideri (b), and Acromyrmex lobicornis (c). Asterisk indicates statistical significant differences (P < 0.05)
Keeping the same traffic lane while turning was the most frequent ant behavior in all leaf-cutting ant species; this behavior was always more frequent than expected by chance (X 2 = 13.8; 47.3; and 18.3 for Atta cephalotes, A. vollenweideri, and Acromyrmex lobicornis, respectively, all P < 0.01, Fig. 4). In general, between 50–70% of the times ants maintain their own traffic lane while turning. Around 20–30% of the times ants crossed one lane while turning and only in less than 10% of the cases ants crossed two lanes (i.e., the whole trail) while turning. However, the probability of being faithful to the traffic lane while turning did not increase with ant flow increments in any of the studied species (X 2 = 0.22; 1.28; and 1.34 for Atta cephalotes, A. vollenweideri, and Acromyrmex lobicornis, respectively, all P > 0.25).

Number of turning ants (%) according to lane fidelity while turning for Atta cephalotes, Atta vollenweideri, and Acromyrmex lobicornis (from top to bottom, respectively). The number of faithful behaviors observed was higher than expected by chance in all species (P < 0.05, see text for details)
Discussion
Collisions between foraging ants produce traffic congestions that may reduce the input of food to the colony (Burd et al. 2002; Burd 2006; Burd and Aranwela 2003). Consequently, it is reasonable that ants follow traffic rules to reduce collisions, especially in crowded-prone locations as the trail corners. Here we demonstrated that leaf-cutting ants often follow a classical traffic rule at bifurcations that reduces the chance of collisions: keep walking on the same lane while turning. In general, ants faithful to its lane while turning had fewer collisions than ants switching lanes. This “lane fidelity” behavior was more frequent than expected by chance. But unexpectedly, the probability of keeping the traffic lane while turning did not increase with ant flow. These evidences suggest that leaf-cutting ants follow simple rules to avoid traffic congestions in trail sectors especially conflicting for circulation, but also that these rules may vary in importance among species.
We showed that for the sampled Atta species, ants faithful to their traffic lane while turning showed ~40% fewer collision rates than those unfaithful. Crossing lanes while turning increased the collision rate because it enhances the probability to block entities walking in opposite direction, especially during high flow (Wang and Nihan 2004; Moussaïd et al. 2011). This traffic condition is common in Atta species, which can reach up to 5000 ants/m2 at the peak of their foraging rate (Bruce and Burd 2012). Accordingly, Atta ants showed this behavior more often than expected by chance, an expected result if ants walk aiming to reduce collisions. Moreover, crossing two lanes while turning (e.g., crossing from side to side the trail)—clearly the worst walking behavior to avoid collisions—was by far the less frequent conduct (<10% of the cases).
Despite Acromyrmex lobicornis foragers being faithful to their traffic lane while turning, this lane fidelity did not reduce the rate of collisions (Fig. 3). Two non-mutually exclusive reasons may explain this pattern. First, as we showed, trails of Acromyrmex species are narrower than Atta trails. Consequently, crossing a traffic lane when turning implies traveling shorter distances for Acromyrmex than for Atta workers, and shortest paths entail smaller chance of collisions with workers walking in opposite directions. Second, it is well known that foraging rate in Acromyrmex is lower than in Atta species (Hölldobler and Wilson 2011; Bruce and Burd 2012). The lower forager density on trails of Acromyrmex species probably reduces the chance of collisions. Supporting these hypotheses, the collision rate in A. lobicornis measured on this work was three times lower than those of Atta (Fig. 3). In sum, trail design and ant flow may help to understand why workers of A. lobicornis faithful and unfaithful to their traffic lane have similar collision rates while turning.
The probability to keep the traffic lane while turning did not increase with increments in ant flow in any of the sampled species. This could be attributed to several reasons. First, and despite we sampled at different times of the day, our measured ant flow did not include the typical high foraging peak of these species (Herz et al. 2007; Bruce and Burd 2012; Hölldobler and Wilson 2011). Lane fidelity while turning may increase in their frequency only in situations of very high ant flow. Second, the maintenance of a traffic lane while turning could be a behavior selected for shortening travel distances rather than for reducing collisions, and thus unaffected by ant flow. To appropriately test this hypothesis, we should experimentally reduce ant flow and monitor whether the frequency of lane fidelity drops or not.
Regardless of whether ants were faithful or unfaithful to their traffic lane while turning, Atta ants showed a high rate of head-on encounters (6–10/min depending if ants were faithful or unfaithful, see Fig. 3). Interestingly, outgoing workers were significantly more likely to contact a worker coming from the opposite direction than returning workers, a similar pattern obtained by Lutz (1929) and confirmed by Bollazzi and Roces (2011). This result is consistent with the idea that head-on encounters are not just simple collisions. Outgoing ants actively search for contacts and/or information regarding food, while ingoing ants mainly try to return as fast as possible to the nest and do not actively search for contacts. This adaptive function of head-con encounters has been demonstrated in some Atta species. Several studies showed that higher rates of head-on encounters increase foraging efficiency through the interchange of information between loaded and unloaded ants (Dussutour et al. 2004, 2007; Farji-Brener et al. 2010; Bouchebti et al. 2015). Our results support these ideas. On the other hand, Acromyrmex lobicornis showed a smaller rate of collisions than Atta ants (3/min, Fig. 3). As discussed above, this can be attributable to the intrinsic low density of workers on foraging trails, or because workers from Acromyrmex species, which are less derived and socially complex than Atta (Hölldobler and Wilson 2011), do not exchange information in head-on encounters. In sum, the consequence of head-on encounters for the colony may range from positive, when food information is exchanged, to negative, when they retard considerably the ant flow. This balance between benefits and costs may vary according the colony starvation level and foraging phases (Bollazzi and Roces 2011). Here we propose that trail design may also affect this balance. In trail sectors intrinsically complicated for traffic flow, like bifurcations, elevated rate of head-on encounters may be disadvantageous despite that they potentially allow information exchange (Elizalde and Farji-Brener 2012). The next step should be to experimentally analyze whether traffic rules are stricter at critical trail sectors for the circulation of foragers, as this work suggests.
References
Bochynek T, Meyer B, Burd M (2017) Energetics of trail clearing in the leaf-cutter ant Atta. Behav Ecol Sociobiol 71:14–19
Bollazzi M, Roces F (2011) Information needs at the beginning of foraging: grass-cutting ants trade off load size for a faster return to the nest. PLoS One 6:e17667
Bouchebti S, Ferrere S, Vittori K, Latil G, Dussutour A, Fourcassié V (2015) Contact rate modulates foraging efficiency in leaf-cutting ants. Sci Rep 5:18650. doi:10.1038/srep18650
Bruce AI, Burd M (2012) Allometric scaling of foraging rate with trail dimensions in leaf-cutting ants. P Roy Soc B-Biol Sci 279:2442–2447. doi:10.1098/rspb.2011.2583
Burd M (2006) Ecological consequences of traffic organisation in ant societies. Phys A 372(1):124–131
Burd M, Aranwela N (2003) Head-on encounter rates and walking speed of foragers in leaf-cutting ant traffic. Insectes Soc 50:3–8
Burd M, Archer D, Aranwela N, Stradling DJ (2002) Traffic dynamics of the leaf-cutting ant, Atta cephalotes. Am Nat 159:283–293
Dussutour A, Fourcassie V, Helbing D, Deneubourg JL (2004) Optimal traffic organization in ants under crowded conditions. Nature 428:70–73
Dussutour A, Beshers S, Deneubourg JL, Fourcassie V (2007) Crowding increases foraging efficiency in the leaf-cutting ant Atta colombica. Insectes Soc 54:158–165
Dussutour A, Deneubourg J-L, Beshers S, Fourcassie V (2009) Individual and collective problem-solving in a foraging context in the leaf-cutting ant Atta colombica. Anim Cogn 12:21–30
Elizalde L, Farji-Brener AG (2012) To be or not to be faithful: flexible fidelity to foraging trails in the leaf-cutting ant Acromyrmex lobicornis. Ecol Entomol 37:370–376
Farji-Brener AG (2001) Why are leaf-cutting ants more common in early secondary forests than in old-growth tropical forests? An evaluation of the palatable forage hypothesis. Oikos 92:169–177
Farji-Brener AG, Ruggiero A (1994) Leaf-cutting ants (Atta and Acromyrmex) inhabiting Argentina: patterns in species richness and geographical range sizes. J Biogeogr 21:391–399
Farji-Brener AG, Tadey M (2017) Consequences of leaf-cutting ants on plant fitness: integrating negative effects of herbivory and positive effects from soil improvement. Insectes Soc 64:1–10
Farji-Brener AG, Amador-Vargas S, Chinchilla F, Escobar S, Cabrera S, Herrera MI, Sandoval C (2010) Information transfer in head-on encounters between leaf-cutting ant workers: food, trail condition or orientation cues? Anim Behav 79:343–349
Farji-Brener AG, Morueta-Holme N, Chinchilla F, Willink B, Ocampo N, Bruner G (2012) Leaf-cutting ants as road engineers: the width of trails at branching points in Atta cephalotes. Insectes Soc 59:389–394
Farji-Brener AG, Chinchilla F, Umaña MN, Ocasio-Torres ME, Chauta-Mellizo A, Acosta-Rojas D, Amador-Vargas S (2015) Branching angles reflect a trade-off between reducing trail maintenance costs or travel distances in leaf-cutting ants. Ecology 96:510–517
Fourcassié V, Dussutour A, Deneubourg JL (2010) Ant traffic rules. J Exp Biol 213:2357–2363
Herz H, Beyschlag W, Hölldobler B (2007) Herbivory rate of leaf-cutting ants in a tropical moist forest in panama at the population and ecosystem scales. Biotropica 39:482–488
Hölldobler B, Wilson EO (2011) The leafcutter ants: civilization by instinct. W. W. Norton, New York
Howard JJ (2001) Costs of trail construction and maintenance in the leaf-cutting ant Atta colombica. Behav Ecol Sociobiol 49:348–356
Kost C, De Oliveira EG, Knoch TA, Wirth R (2005) Spatio-temporal permanence and plasticity of foraging trails in young and mature leaf-cutting ant colonies (Atta spp.). J Trop Ecol 21:677–688
Lugo AE, Farnworth EG, Pool D, Jerez P, Kaufman G (1973) The impact of the leaf cutter ant Atta colombica on the energy flow of a tropical west forest. Ecology 54:1292–1301
Lutz FE (1929) Observations on leaf-cutting ants. Am Mus Novit 388:1–21. doi:10.1206/0003-0082
Moussaïd M, Helbing D, Theraulaz G (2011) How simple rules determine pedestrian behavior and crowd disasters. Proc Natl Acad Sci 108:6884–6888
R Development Core Team (2015) R: a language and environment for statistical computing. The R Foundation for Statistical Computing, Vienna
Rockwood LL, Hubbell SP (1987) Host-plant selection, diet diversity, and optimal foraging in a tropical leafcutting ant. Oecologia 74:55–61
Shepherd JD (1982) Trunk trails and the searching strategy of a leaf-cutter ant, Atta colombica. Behav Ecol Sociobiol 11:77–84
Wang Y, Nihan NL (2004) Estimating the risk of collisions between bicycles and motor vehicles at signalized intersections. Accid Anal Prev 36:313–321
Wirth R, Herz H, Ryel R, Beyschlag W, Hölldobler B (2003) Herbivory of leaf-cutting ants. A case study on Atta Colombica in the tropical rainforest of Panama. Springer, Berlin
Acknowledgements
This work was partially supported by the Fondo para la Investigación Científica y Tecnológica, Argentina (FONCYT; PICT 2011-1406 and 2015-1319) and CONICET, Argentina (PIP 2014-2016, 11220130100665-CO) to AFGB, and Emily Forester and Hovore-Horn fellowships from the Organization for Tropical Studies to LCM. The Organization for Tropical Studies (OTS) and The Conservation Land Trust (CLT) also provided logistical support. Sabrina Amador-Vargas and two anonymous reviewers made helpful comments on earlier version of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Cibils-Martina, L., Elizalde, L. & Farji-Brener, A.G. Traffic rules around the corner: walking of leaf-cutting ants at branching points in trunk trails. Insect. Soc. 64, 549–555 (2017). https://doi.org/10.1007/s00040-017-0576-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00040-017-0576-5