
Abstract
Ropalidia cyathiformis is a primitively eusocial tropical paper wasp usually with a single dominant queen per colony. When the queen is removed, one and only one individual increases her aggression and becomes the new queen of the colony, unchallenged by any other worker. We refer to such a successor as a potential queen (PQ) until she lays her first egg. By removing the queen and successive PQs, we show that there is not just one successor but a strict reproductive hierarchy of up to 3 PQs, who succeed the queen one after the other. Of many variables tested, we found that only the frequency of dominance behaviour is a significant predictor of whether or not an individual is part of the reproductive hierarchy and also of her position in the hierarchy. Dominance behaviour, however, does not perfectly predict the position of an individual in the reproductive hierarchy because we show that an average of three more dominant individuals, are bypassed when an individual becomes the next queen or PQ. This is in contrast to the reproductive hierarchy in the conspecific Ropalidia marginata, where age rather than dominance behaviour is a predictor (though imperfect once again) of an individual’s position in the queue. Taken together, our results suggest that (a) these two sister species have evolved two rather different mechanisms of reproductive caste differentiation, (b) that neither of them strictly conform either to the so-called “temperate” or “tropical” patterns of queen succession seen in most other species studied so far.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Reproductive caste differentiation into one or a small number of fertile kings and queens and a large number of sterile workers is a hallmark of social insect colonies (Michener 1969; Wilson 1971). In highly eusocial species such as honeybees and most ants, non-reproductive individuals cannot usually replace dead or lost reproductive individuals because replacement reproductives have to be raised from the pre-imaginal stage. In primitively eusocial species such as most social bees and wasps on the other hand, workers can mate, develop their ovaries and become replacement queens following the death or loss of existing queens (de Wilde and Beetsma 1982; Fletcher and Ross 1985; Watson et al. 1985; Gadagkar et al. 1988; O’Donnell 1998).
Unlike the process of raising a new queen from the pre-imaginal stage, queen succession from among the adult workers is expected to provide more opportunities for the expression of conflict. For example, the death of a queen can result in scramble competition among the workers leading to a chaotic situation in the colony, until one of the workers becomes the next queen. This would imply that there is no pre-designated successor to the queen. Alternatively queen succession might happen more smoothly without much overt competition because there is a pre-designated successor, perhaps even a pre-designated queue of several successors who take over the role of the queen as and when their turn arises. The latter scenario in fact appears to be the case in the primitively eusocial wasp Ropalidia marginata. Upon loss or experimental removal of the queen, one and only one individual becomes temporarily hyper-aggressive and goes on to become the next queen. This hyper-aggressive ‘potential queen’ (PQ), however, is never challenged by any other worker (Premnath et al. 1996; Deshpande et al. 2006). We have reason to believe that the unidirectional aggression from the potential queen to many different workers is not really an expression of conflict (Lamba et al. 2007; Shukla et al. 2014). If the queen and potential queens are successively removed, a series of similar temporarily hyper-aggressive, unchallenged potential queens become apparent, indicating the presence of a long reproductive queue of up to five individuals. In this species the age of individuals and not their dominance behaviour is a statistically significant correlate of the position of an individual in the reproductive queue of the colony (Bang and Gadagkar 2012).
The fact that the reproductive queue is not dominance based in R. marginata is not surprising when considering the fact that their colonies are headed by meek and docile, non-interactive ‘sitter’ queens (Gadagkar and Joshi 1983; Chandrashekara and Gadagkar 1991; Kardile and Gadagkar 2002; Sumana and Gadagkar 2003) unlike other primitively eusocial wasps, where the queens are highly aggressive and are the most dominant individuals in the colony (Pardi 1948; West-Eberhard 1969; Jeanne 1972; Strassmann and Meyer 1983; Hughes et al. 1987; Bridge and Field 2007). Ropalidia cyathiformis, another tropical primitively eusocial wasp and a sister species to R. marginata has highly aggressive, interactive ‘fighter’ queens, usually at the top of the dominance hierarchies of their colonies (Gadagkar and Joshi 1982, 1984; Deshpande et al. 2006). Here we have performed serial PQ removal experiments, similar to those we have previously performed in R. marginata. Our aim was to determine whether queen succession in this species with aggressive queens is based on scramble competition or whether there is a conflict free succession based on a pre-designated reproductive queue, and if the latter, whether the queue is based on age or dominance behaviour.
Methods
This study was conducted on the campus of the Indian Institute of Science, Bangalore (13°00′N and 77°32′E), India, between December 2011 and October 2014. Ropalidia cyathiformis is a tropical primitively eusocial wasp, in which nests are initiated throughout the year by one or a small group of female wasps leaving their natal nest. For this reason there is no sharp difference between foundresses in pre-emergence nests and female offspring produced in post-emergence nests. There is no morphological difference between egg-layers and non egg-layers (Kardile and Gadagkar 2005).
A total of 19 post-emergence nests of R. cyathiformis were studied in their natural sites of initiation. All adults were marked using Testors® enamel paints for individual identification. The nests used in the current study contained 10–20 female wasps; there were no more than one or two males in one or two nests which were not included in the analysis. We did not witness the founding of these nests and, therefore, do not know whether they were founded by a single foundress or multiple foundresses.
Detecting the reproductive queue
Nine selected nests were observed for 10 h each during two consecutive days in their queen-right condition (unmanipulated nests with queen present). Behavioural observations were made from 0800 to 1030 hours and 1300 to 1530 hours on day 1, and from 1030 to 1300 hours and 1530 to 1800 hours on day 2. Behavioural observations consisted of equal numbers of ‘scans’ (in which the behavioural state of every individual was recorded at randomly chosen instants of time) and ‘all occurrence sessions’ (AOS) (in which every occurrence of every selected behaviour by every individual was recorded during 5 min intervals), randomly intermingled with each other with a 1 min gap between two consecutive sessions. Hence at the end of 2 days there were 10 h of observations with 50 scans and 50 all occurrence sessions. Scans were used to estimate the proportions of time spent by each wasp in different behaviours. All occurrence sessions were used to estimate the frequency per hour with which different wasps perform different behaviours. Although the scans provided us information on the proportion of time spent by each wasp in over 100 behaviours, only the proportion of time spent being absent from the nest has been used in this study; data on other behaviours will be used in separate studies. From the AOS sessions we obtained data on frequency per hour at which each wasp performed over 50 behaviours. Of these we have used all dominance behaviours, maintenance behaviours and feeding behaviours (see below for a more detailed description of the sub-components of these behaviours) in this study.
On the morning of day 3, at 0800 hours, the queen (who was identified based on egg laying during behavioural observations on days 1 and 2) was removed. To identify the potential queen (PQ), the rest of the nest was observed for 6 h from 0800–1000 hours, 1100–1300 hours and 1400–1600 hours to yield 30 scans and 30 all occurrence sessions. To be designated as a PQ an individual was required to possess both of the following criteria namely, (1) show at least five acts of dominance behaviour and (2) show more dominance behaviour than any other individual in the colony. Although the criterion of showing at least five acts of dominance behaviour is admittedly arbitrary, we included it to ensure that the PQ was unique among the workers and not simply one of the several individuals in a continuum of dominance behaviours. On each subsequent day, the PQ (PQ1, PQ2 or PQ3, as the case maybe) identified on the previous day was removed and the rest of the nest was observed as on day 3. After removal of PQ2 (in 2 nests) and after removal of PQ3 (in 6 nests), no subsequent PQs were apparent by the criteria described above after one more day of observation, so the experiment was, therefore, terminated. One nest was abandoned after PQ3 was identified and, therefore, could not be observed on the following day. Using ten additional nests and conducting similar serial queen/PQ removal experiments, 4 PQ1s, 3 PQ2s and 3 PQ3s were not removed to see whether they would go on to become the sole egg-layers in their colonies. We monitored these nests until egg laying was observed after which the colony was again observed to identify the egg layer. Finally all the individuals were collected and stored at −20 °C.
The wasps were then dissected in Insect Ringer’s solution. The following ovarian parameters were measured for each wasp: (1) number of mature oocytes, (2) mean length of all six proximal oocytes, (3) mean width of all six proximal oocytes, (4) total number of oocytes with yolk and (5) total number of oocytes. Proximal oocytes included mature oocytes if present. These parameters were then subjected to principal components analysis (PCA) and principal component 1 was used as the ovarian index for each wasp (Chandrashekara and Gadagkar 1991; Gadagkar 2001). To be designated as the sole egg layer of the colony, an individual was required to possess three criteria: (1) be the only individual who laid any eggs, (2) have the highest ovarian index among all the members of the colony and (3) be the only individual who has mature oocytes. Criteria 2 and 3 are required because we also find a secondary egg layer in very rare cases.
Determinants of becoming a PQ and of the position in the queue
To investigate possible determinants of becoming a PQ and of the position of an individual in the reproductive hierarchy, the following predictor variables were measured:
-
1.
frequency/hour of dominance behaviour, which includes attack, nibble, aggressive bite, peck, hold in mouth and chase,
-
2.
frequency/hour of maintenance behaviour which includes build new cells, extend old cells, mouth walls of old cells, shape cell walls, coat pedicel, mouth pedicel, clean empty cells, nibble the back of the nest, remove meconium, antennate nest, fan the nest,
-
3.
frequency/hour of feed larva,
-
4.
proportion of time spent foraging (being away from the nest), all measured in queen-right colonies and
-
5.
age in days (since eclosion) on day 1 of the experiment. For a more detailed description of the above behaviours see Gadagkar (2001).
In six nests for which age data were used, the nests were followed before commencing the experiment (after marking all individuals and conducting daily census of wasps present) until all wasps of unknown age except the queen, disappeared.
We investigated the possible role of the above mentioned predictor variables in influencing the response variables (becoming a PQ or not and becoming PQ1, PQ2 and PQ3) using binary and ordinal logistic regression models, respectively. To account for possible lack of independence between different values of the response variables (becoming a PQ or not and becoming PQ1, PQ2 or PQ3), the binary logistic regression models and the ordinal logistic regression models described above were re-run 10,000 times after randomising the predictor variables (Zanette and Field 2009; Green et al. 2013). All the analysis was done in R 2.12.1.
To see whether dominance behaviour is a perfect predictor of the position of an individual in the reproductive queue we performed a ‘queue-jumping’ analysis by counting the number of individual or individuals showing more dominance behaviour that were bypassed whenever individuals became PQs.
Results
Reproductive queue in Ropalidia cyathiformis
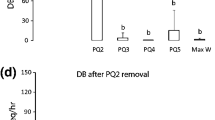
In a total of 19 colonies of R. cyathiformis, we removed the queen and/or PQs to determine the identity of the next PQ or to confirm that the individual identified as PQ would indeed go on to become the next sole egg layer of the colony. Upon experimental removal of the queen, a single individual became an unchallenged potential queen (PQ1) (by the criteria defined in the “Methods”) in 12 out of 12 colonies. Upon removal of PQ1, a single unchallenged individual became the next potential queen (PQ2) in 12 out of 12 colonies. Upon removal of PQ2, a PQ3 became apparent in 10 of 12 colonies. In the remaining two colonies no subsequent potential queens were apparent. Upon removal of the PQ3 no subsequent potential queens were apparent in any of the six colonies tested. In all cases in which the PQ was identifiable, a single individual became sufficiently aggressive to be classified as PQ according to the criteria given in the Methods section, and no PQ was ever challenged by any of the workers in her colony. Thus, we observed a reproductive queue of up to three individuals in all colonies studied (Figs. 1, 2). It does not appear that our results are sensitive to the criteria for defining PQs as given in the “Methods”. The individuals that were not designated as PQs did not stand out as unique in their dominance behaviour in their respective colonies. This is quite evident in Fig. 1 where all individuals we identified as PQs stood out from the rest (see panels b–d) and there was really no unique individual when we failed to find PQ4 (see panel e). In the two colonies where we failed to find a PQ3, no individual showed any dominance behaviour in one of the nests. In the other nest, two individuals were tied with two acts of dominance behaviour each while another individual showed one act of dominance behaviour and the remaining five individuals showed no dominance behaviour at all.

Reproductive queue in R. cyathiformis. The mean and standard deviations of the frequencies per hour of dominance behaviour of the queen, 3 PQs and M.W (max worker, which is the individual who showed the highest frequency of dominance behaviour other than the Q and all three PQs), in normal queen-right colonies (a) and in the absence of the queen and the preceding PQs (b–d) (n = 12 for a–c and 10 for d and 8 for b). Bars carrying different alphabets are significantly different from each other within each panel (Wilcoxon matched-pairs, signed-ranks test; to achieve P < 0.05, α was set to 0.008, 0.01 and 0.05 on account of Bonferroni correction for b, c and d, respectively). Bars carrying different numbers show the significant difference in the frequency per hour of dominance behaviour shown by the focal PQ in the queen-right session with itself after the removal of the queen or previous PQs (Wilcoxon matched-pairs, signed-ranks test; to achieve P < 0.05, α was set to 0.01 on account of Bonferroni correction). All the PQs showed significantly higher dominance behaviour after the removal of the queen and the previous PQs compared to itself in the presence of the queen. Also, after the removal of PQ3 (and in two nests after PQ2) there was no identifiable PQ4. The dominance shown by the M.W was not significantly different from the M.W in the queen-right session. Also there was no significant difference in aggression shown between the M.W and the next most dominant individual in the colony (data not shown, Wilcoxon matched-pairs, signed-ranks test; P > 0.05). DB dominance behaviour, Q queen, PQ1–PQ3 potential queens 1–3, M.W max worker

Mean and standard deviations of dominance behaviour shown and received by PQs, after the removal of queen/previous PQs. Bars carrying different alphabets are significantly different from each other for each PQ (Wilcoxon matched-pairs, signed-rank test; P < 0.05; n = 19, 15 and 10 colonies each for PQ1, PQ2 and PQ3). DB dominance behaviour, S− dominance received, PQ1–PQ3 potentials queens 1–3
PQs seem to show dominance behaviour to many individuals in the colony, ranging from 15% of the workers (5 out of 34 individuals in one colony) up to 79% (11 out of 14 individuals in another colony). It also appears that PQs do not direct their dominance behaviour toward the next PQ in the queue. In 25 successions for which we had the necessary data, there were 8 cases in which the next PQ received no dominance behaviour at all, although other wasps received dominance. In 12 cases, the next PQ received some dominance behaviour, but there were other workers that received more. In two cases the next PQ was tied with two other individuals in receiving the highest dominance. Only in three cases the next PQ received the highest amount of dominance behaviour compared to any other worker in the colony.
In all ten nests where no subsequent wasps were removed after a PQ was identified, the individual who was identified as PQ by dominance behaviour went on to become the sole egg layer in her colony, as judged by the criteria defined in the “Methods”. The time taken for laying the first egg was 1–9 days (mean ± SD = 5.2 ± 3.5) for PQ1s, 3–7 days (mean ± SD = 5.3 ± 2.1) for PQ2s and 4–14 days (mean ± SD = 7.7 ± 5.5) for PQ3s.
Determinants of becoming a PQ
As noted above no subsequent PQs were apparent after removing the PQ2 in two colonies and after removing the PQ3 in six colonies. Assuming that the remaining workers that did not become PQs were incapable of doing so, we attempted to identify the determinants of becoming a PQ. Of all the variables tested only the coefficient associated with dominance behaviour was statistically significant (Table 1). The remaining variables, frequency per hour of maintenance behaviour and feed larva, proportion of time spent in foraging and age (measured in days since eclosion) were not significant. Hence individuals showing more dominance behaviour were more likely to become a PQ.
Determinants of the position of an individual in the reproductive queue
In model testing the role of different predictor variables in determining the position of an individual in the reproductive queue, only dominance behaviour was statistically significant (Table 2). As in the previous case all remaining variables were not significant. Hence the PQ showing the highest dominance behaviour becomes PQ1, next highest becomes PQ2 and third highest becomes PQ3.
We confirmed that the coefficients obtained in the logistic regression models for dominance behaviour could not have been obtained by chance. In the randomised data sets we obtained coefficient values equal to or higher than those in the original data in only 20 out of 10,000 simulations in the binary logistic regression models and in only 50 out of 10,000 simulations in the ordinal logistic regression models.
Queue-jumping analysis
Of 34 PQs studied, 24 jumped the dominance queue by by-passing one or more individuals more dominant than themselves. On average 3.1 ± 3.0 more dominant individuals were bypassed during queen/PQ successions; 1.2 ± 1.9 more dominant individuals were bypassed by PQ1s, 4.1 ± 3.2 more dominant individuals were bypassed by PQ2s and 4.2 ± 3.0 more dominant individuals were bypassed by PQ3s (Table 3). On average, the differences in dominance behaviour were smaller when there were by-passes than when there were no by-passes (Mann–Whitney U test, P < 0.05).
Discussion
As seen from this study on R. cyathiformis and a previous study on R. marginata (Bang and Gadagkar 2012), removal of a queen result in a smooth, apparently conflict free succession of a new queen based on a reproductive queue, rather than any form of scramble competition among the workers. Although there was a great deal of dominance behaviour upon removal of the queen, all of it emanated from a single potential queen who did not receive any dominance behaviour from any of the workers. Moreover, aggression of the PQs was not directed towards the next PQ in the queue and was instead widely distributed among the members of the colony. Such unidirectional and widely distributed dominance behaviour by the potential queen is thus not likely to be an indication of inter-individual conflict for the position of the next queen. We find that dominance behaviour is a significant predictor both of the probability of an individual becoming a PQ and their position in the queue for those that became PQs. Dominance behaviour, however, is not a perfect predictor of an individual’s position in the queue; much queue-jumping happens. About 70% of the PQs bypass one or more dominant individuals when they become PQs although queue-jumping is more common when the differences in dominance behaviour between individuals are relatively small. Such queue-jumping coupled with the observation that there is no scramble competition (PQs are never challenged) suggests that there may be other factors that may be better determinants of an individual’s position in the queue—factors which may be only imperfectly correlated with dominance behaviour.
One difference between the two species is that in R. marginata five successive PQs were identified and perhaps more could be identified if the experiment had been continued (Bang and Gadagkar 2012). In R. cyathiformis, however, we obtained a maximum of three PQs and no further PQs were evident after removing PQ3. Although we cannot be sure that none of the individuals could ever become a PQ after removal of PQ3, it is clear that the individuals who did not become PQs are distinctly different from those that did become PQs. It must be noted that when we did identify a PQ, that individual showed about an order of magnitude more dominance behaviour than the next most dominant individual. This was clearly not the case for individuals that we did not designate as PQs (see Fig. 1). It thus appears that the queue is shorter in R. cyathiformis as compared to R. marginata. One reason for this could be that R. cyathiformis colonies are generally smaller than those of R. marginata. A more interesting reason could be that the queue is shorter in R. cyathiformis because it is based on dominance behaviour and not on age. In both species queen succession happens in an orderly fashion without scramble competition. To the extent that such orderly succession is adaptive (see below) one can consider that the function of the reproductive queue is to bring about orderly succession and avoid scramble competition. In R. marginata where the queue is age based, new individuals can join only at the bottom of the queue. In R. cyathiformis the queue is not based on age, but only on dominance behaviour. So, young individuals can join the queue at the top or in the middle. This would make the queue more unstable and subject to change. A third reason for the shorter queues in R. cyathiformis could be that because the queue is based on age in R. marginata, all individuals may have a finite chance of becoming future queens because they would eventually become the older individuals. In R. cyathiformis on the other hand, individuals incapable of showing higher dominance behaviour may never have a chance of becoming queens because there is no guarantee that their dominance behaviour will increase with time. In other words, there may be a subset of workers who are more or less doomed to remain sterile throughout their lives. Indeed dominance behaviour is a significant predictor of whether or not an individual can become a potential queen.
Although queen or serial queen removal experiments have been performed in a few species, it is not entirely clear how common or unique the smooth transition from one queen to the next, unaccompanied by overt signs of inter-individual conflict, is among primitively eusocial wasps. In Polistes canadensis, Polistes fuscatus and Polistes chinensis (Morimoto 1961) “heightened aggressiveness among workers” has been observed after queen removal (West-Eberhard 1969). In Polistes exclamans queen succession “did not involve a great increase in aggression” with “one notable exception”. However, queen succession also appears to depend on dominance because “before becoming queen, replacement queens … were more aggressive than females who never became a queen” (Strassmann and Meyer 1983). In Polistes annularis queen succession is based on dominance, involving less aggression when foundresses become new queens, but relatively more aggression when workers become new queens, although workers become queens only when no foundresses are left (Hughes et al. 1987). In Liostenogaster flavolineata queen succession is based on age which in turn is perfectly correlated with dominance rank, but “there is no evidence for an increase in aggressive activity during succession” (Cronin and Field 2007). Thus, it appears that what is unique about queen succession in R. marginata and R. cyathiformis is that there is a great deal of aggression, but it is unidirectional and attributable to a single individual namely the successor (PQ). This type of succession with heightened aggression in the colony following the disappearance of the queen, but with no conflict among the workers for the position of the queen has also been described in one other species, Mischocyttarus cassununga, although no quantitative data about the dominance shown and received by the successor are given (Murakami and Shima 2009).
From the literature we have located information about queen succession, natural or induced (by queen removal), usually for a single succession, but occasionally for successive successions (as in our study) for 18 species of primitively eusocial wasps. The most important generalisation to emerge from these studies is that there are two different patterns of queen succession, a so-called “temperate” pattern in which older/oldest wasps become future queens and a so-called “tropical” pattern in which younger wasps become future queens (Strassmann and Meyer 1983; Suzuki 2003). One of the unique aspects about our results is that queen succession is based on dominance behaviour, but not age in R. cyathiformis and age, but not dominance behaviour in R. marginata. This is unique also because both dominance behaviour and age are associated with queen succession in most other primitively eusocial wasps. Dominance behaviour predicts future queens in all primitively eusocial wasps studied until now except for R. marginata. Age also predicts future queens in other primitively eusocial wasps, but the relationship of age with queen succession is of two types. The oldest/older wasps in the colony become future queens (“temperate pattern”) in the case of Polistes dominulus (Pardi 1948), P. jadwigae (Yoshikawa 1963), P. fuscatus (West-Eberhard 1969), M. flavitarsis (Litte 1979), P. metricus (Dew and Michener 1981), P. exclamans (Strassmann and Meyer 1983), P. chinensis antennalis (Miyano 1986), P. annularis (Hughes et al. 1987), P. instabilis (Hughes and Strassmann 1988), P. lanio (Giannotti and Machado 1997), Belonogaster petiolata (Keeping 1997), Liostenogaster flavolineata (Bridge and Field 2007) and Mischocyttarus cassununga (Murakami and Shima 2009). On the other hand relatively younger wasps supersede the queen (“tropical pattern”) in Polistes canadensis (West-Eberhard 1969), Mischocyttarus drewseni (Jeanne 1972), Metapolybia aztecoides (West-Eberhard 1978) and Parapolybia indica (Suzuki 2003). Dominance behaviour might be functioning as an honest signal of an individual’s fitness and age might indicate the timing of ovarian maturity. Thus, the temperate or tropical patterns of queen succession may correspond to the different patterns in age dependent maturation of ovaries in different species (Suzuki 2003).
R. cyathiformis does not seem to conform to either of these two queen succession patterns because age is not a significant predictor in spite of the fact that the age distribution in the colonies studied ranged from 1 to 115 days. Indeed, the ages of the individuals who became potential queens ranged all the way from 7 to 86 days. There seems to be one other species with a similar pattern of age being unimportant, Belonogaster juncea juncea, although the range here was not quite so large (7–28 days) (Tindo et al. 2002). What appears to be even more unique about queen succession in R. marginata is that it is not based on dominance behaviour and only on age. In all other species where queen succession is age based, it is also based on dominance. It is interesting that two sister species, R. cyathiformis and R. marginata have evolved such different mechanisms of reproductive caste differentiation. We speculate that R. cyathiformis, like other primitively eusocial wasps discussed above, is a typical primitively eusocial species in which dominance behaviour is a key regulator of reproductive division of labour. On the other hand R. marginata, has attained some features of more highly eusocial species such as (1) use of pheromones for regulation of reproduction (Bhadra et al. 2010; Mitra et al. 2011), (2) a strong age polyethism (Naug and Gadagkar 1998) and (3) diminishing importance of dominance behaviour in reproductive caste differentiation and queen succession (Bang and Gadagkar 2012). This may explain why these two sister species have diverged in their mechanisms of queen succession.
Why should there be such a smooth conflict free queen succession in R. marginata and R. cyathiformis? Both species nest throughout the year in their tropical habitat which leads to many opportunities for becoming replacement queens in the natal nest or leaving to found their own nest. We speculate that the risks involved in scramble competition may not be worth taking in these species.
References
Bang A, Gadagkar R (2012) Reproductive queue without overt conflict in the primitively eusocial wasp Ropalidia marginata. Proc Natl Acad Sci USA 109:14494–14499. doi:10.1073/pnas.1212698109
Bhadra A, Mitra A, Deshpande SA et al (2010) Regulation of reproduction in the primitively eusocial wasp Ropalidia marginata: on the trail of the queen pheromone. J Chem Ecol 36:424–431
Bridge C, Field J (2007) Queuing for dominance: gerontocracy and queue-jumping in the hover wasp Liostenogaster flavolineata. Behav Ecol Sociobiol 61:1253–1259. doi:10.1007/s00265-007-0355-9
Chandrashekara K, Gadagkar R (1991) Behavioural castes, dominance and division of labour in a primitively eusocial wasp. Ethology 87:269–283. doi:10.1111/j.1439-0310.1991.tb00252.x
Cronin AL, Field J (2007) Social aggression in an age-dependent dominance hierarchy. Behaviour 144:753–765
de Wilde J, Beetsma J (1982) The physiology of caste development in social insects. Adv Insect Physiol 16:167–246
Deshpande SA, Sumana A, Surbeck M, Gadagkar R (2006) Wasp who would be queen: a comparative study of two primitively eusocial species. Curr Sci 91:332–336
Dew HE, Michener CD (1981) Division of labor among workers of Polistes metricus (Hymenoptera: Vespidae): laboratory foraging activities. Insectes Soc 28:87–101. doi:10.1007/BF02223625
Fletcher DJC, Ross KG (1985) Regulation of reproduction in eusocial hymenoptera. Annu Rev Entomol 30:319–343. doi:10.1146/annurev.en.30.010185.001535
Gadagkar R (2001) The Social Biology of Ropalidia marginata: toward understanding the evolution of eusociality. Harvard University Press, Cambridge
Gadagkar R, Joshi NV (1982) Behaviour of the Indian social wasp Ropalidia cyathiformis on a nest of separate combs (Hymenoptera: Vespidae). J Zool 198:27–37. doi:10.1111/j.1469-7998.1982.tb02058.x
Gadagkar R, Joshi NV (1983) Quantitative ethology of social wasps: time-activity budgets and caste differentiation in Ropalidia marginata (Lep.) (Hymenoptera: Vespidae). Anim Behav 31:26–31. doi:10.1016/S0003-3472(83)80170-5
Gadagkar R, Joshi NV (1984) Social organisation in the Indian wasp Ropalidia cyathiformis (Fab.) (Hymenoptera: Vespidae). Z Für Tierpsychol 64:15–32. doi:10.1111/j.1439-0310.1984.tb00350.x
Gadagkar R, Vinutha C, Shanubhogue A, Gore AP (1988) Pre-imaginal biasing of caste in a primitively eusocial insect. Proc R Soc Lond B Biol Sci 233:175–189. doi:10.1098/rspb.1988.0017
Giannotti E, Machado VLL (1997) Queen replacement in post-emergent colonies of the social wasp, Polistes lanio (hymenoptera, vespidae). Rev Bras Entomol 41:9–11
Green JP, Leadbeater E, Carruthers JM et al (2013) Clypeal patterning in the paper wasp Polistes dominulus: no evidence of adaptive value in the wild. Behav Ecol 24:623–633. doi:10.1093/beheco/ars226
Hughes CR, Strassmann JE (1988) Age is more important than size in determining dominance among workers in the primitively eusocial wasp, Polistes instabilis. Behaviour 107:1–14
Hughes CR, Beck MO, Strassmann JE (1987) Queen succession in the social wasp, Polistes annularis. Ethology 76:124–132. doi:10.1111/j.1439-0310.1987.tb00678.x
Jeanne RL (1972) Social biology of the neotropical wasp Mischocyttarus drewseni. Bull Mus Comp Zool 144:63–150
Kardile SP, Gadagkar R (2002) Docile sitters and active fighters in paper wasps: a tale of two queens. Naturwissenschaften 89:176–179. doi:10.1007/s00114-002-0306-2
Kardile SP, Gadagkar R (2005) Observations on the natural history and behaviour of the primitively eusocial wasp Ropalidia cyathiformis (Fab.) (Hymenoptera: Vespidae). J Bombay Nat Hist Soc 102:265–273
Keeping MG (1997) Social behavior and brood decline in reproductive-phase colonies of Belonogaster petiolata (Degeer) (Hymenoptera: Vespidae). J Insect Behav 10:265–278. doi:10.1007/BF02765559
Lamba S, Kazi YC, Deshpande S et al (2007) A possible novel function of dominance behaviour in queen-less colonies of the primitively eusocial wasp Ropalidia marginata. Behav Process 74:351–356. doi:10.1016/j.beproc.2006.12.003
Litte M (1979) Mischocyttarus flavitarsis in Arizona: social and nesting biology of a polistine wasp. Z Für Tierpsychol 50:282–312. doi:10.1111/j.1439-0310.1979.tb01033.x
Michener CD (1969) Comparative social behavior of bees. Annu Rev Entomol 14:299–342. doi:10.1146/annurev.en.14.010169.001503
Mitra A, Saha P, Chaoulideer ME et al (2011) Chemical communication in Ropalidia marginata: Dufour’s gland contains queen signal that is perceived across colonies and does not contain colony signal. J Insect Physiol 57:280–284
Miyano S (1986) Colony development, worker behavior and male production in orphan colonies of a Japanese paper wasp, Polistes chinensis antennalis Pérez (Hymenoptera: Vespidae). Res Popul Ecol 28:347–361
Morimoto R (1961) On the dominance order in the Polistes wasps. I. (Studies on the social Hymenoptera of Japan. XII). Sci Bull Fac Agric Kyushu Univ 18:339–351
Murakami ASN, Shima SN (2009) Queen replacement in Mischocyttarus (Monocyttarus) cassununga (Hymenoptera, Vespidae, Mischocyttarini): a particular case. Sociobiology 53:247–257
Naug D, Gadagkar R (1998) The role of age in temporal polyethism in a primitively eusocial wasp. Behav Ecol Sociobiol 42:37–47
O’Donnell S (1998) Reproductive caste determination in eusocial wasps (Hymenoptera: Vespidae). Annu Rev Entomol 43:323–346. doi:10.1146/annurev.ento.43.1.323
Pardi L (1948) Dominance order in Polistes wasps. Physiol Zool 21:1–13
Premnath S, Sinha A, Gadagkar R (1996) Dominance relationship in the establishment of reproductive division of labour in a primitively eusocial wasp (Ropalidia marginata). Behav Ecol Sociobiol 39:125–132. doi:10.1007/s002650050274
Shukla S, Pareek V, Gadagkar R (2014) Ovarian development in a primitively eusocial wasp: social interactions affect behaviorally dominant and subordinate wasps in opposite directions relative to solitary females. Behav Process 106:22–26. doi:10.1016/j.beproc.2014.04.003
Strassmann JE, Meyer DC (1983) Gerontocracy in the social wasp, Polistes exclamans. Anim Behav 31:431–438. doi:10.1016/S0003-3472(83)80063-3
Sumana A, Gadagkar R (2003) Ropalidia marginata—a primitively eusocial wasp society headed by non-dominant queens. Curr Sci 84:1464–1468
Suzuki T (2003) Queen replacement without gerontocracy in the paper wasp Parapolybia indica in temperate Japan. Ethol Ecol Evol 15:191–196. doi:10.1080/08927014.2003.9522683
Tindo M, Mony R, Dejean A (2002) Colony development and serial polygyny in the primitively eusocial wasp Belonogaster juncea juncea (Vespidae: Polistinae). J Insect Behav 15:243–252. doi:10.1023/A:1015489017927
Watson JAL, Okot-Kotber BM, Noirot C (1985) Caste differentiation in social insects. Elsevier, Amsterdam
West-Eberhard MJ (1969) The social biology of polistine wasps. Museum of Zoology, University of Michigan, Ann Arbor
West-Eberhard MJ (1978) Temporary queens in metapolybia wasps: nonreproductive helpers without altruism? Science 200:441–443. doi:10.1126/science.200.4340.441
Wilson EO (1971) The insect societies. Belknap Press of Harvard University Press, Cambridge
Yoshikawa K (1963) Introductory studies on the life economy of polistine wasps: ii. Superindividual stage: 2. Division of labor among workers. Jpn J Ecol 13:53–57
Zanette L, Field J (2009) Cues, concessions, and inheritance: dominance hierarchies in the paper wasp Polistes dominulus. Behav Ecol 20:773–780. doi:10.1093/beheco/arp060
Acknowledgements
This work was supported by grants (to RG) from the Department of Science and Technology (including DST-FIST program), Department of Biotechnology (including DBT-IISc Partnership Program), Council of Scientific and Industrial Research, and Ministry of Environment, Forests and Climate Change, Government of India. SU was supported by a Research Fellowship from the University Grants Commission. We thank Dr. Kavita Isvaran for advice on statistical analysis. We thank Nitika Sharma for assisting in observations for two nests. RG and SU co-designed the study, SU conducted the study and RG and SU co-wrote the paper.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Unnikrishnan, S., Gadagkar, R. Dominance based reproductive queue in the primitively eusocial wasp, Ropalidia cyathiformis . Insect. Soc. 64, 495–503 (2017). https://doi.org/10.1007/s00040-017-0568-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00040-017-0568-5