
Abstract
Megakaryocytes are rare cells found in the bone marrow, responsible for the everyday production and release of millions of platelets into the bloodstream. Since the discovery and cloning, in 1994, of their principal humoral factor, thrombopoietin, and its receptor c-Mpl, many efforts have been directed to define the mechanisms underlying an efficient platelet production. However, more recently different studies have pointed out new roles for megakaryocytes as regulators of bone marrow homeostasis and physiology. In this review we discuss the interaction and the reciprocal regulation of megakaryocytes with the different cellular and extracellular components of the bone marrow environment. Finally, we provide evidence that these processes may concur to the reconstitution of the bone marrow environment after injury and their deregulation may lead to the development of a series of inherited or acquired pathologies.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Bone marrow (BM) is a complex tissue protected by bones and dedicated to the production of white, red blood cells and platelets [1]. Packaged into the cavity of bones, the BM is a tridimensional network of branching sinusoids surrounding islets of hemopoietic cells immersed in a mesh of extracellular matrix (ECM) components and soluble/humoral factors [2–5]. Mechanisms of single cell differentiation and transit to the bloodstream must be tightly regulated to satisfy physiological blood cell requests and supplies. To this regard, recent insights in the characterization of BM structure and function has allowed the identification of specialized microenvironment implicated in the direction of hemopoietic stem cell (HSC) quiescence, differentiation or mobilization [6, 7]. Specifically, cell composition and localization has let to discriminate a peri-vascular niche mainly composed of endothelial, CXCL12+ reticular cells and Nestin+ mesenchymal stromal cells [8, 9] from an endosteum-associated niche occupied by osteoblasts and osteolineage cells and NG2+ (CSPG4) pericytes [10–12]. Despite the fact that a coordinated crosstalk and connection between these niches is indispensable for efficient HSC differentiation and BM function, the anatomical definition of those districts as separated or overlapping entities is still debated [13]. Within this context, megakaryocytes (Mks) migrate, while differentiating from HSCs, from the endosteal to the vascular niche where platelet are shed into the bloodstream [14, 15]. Mks are historically associated with the vascular BM niche as they were detected adjacent to marrow sinusoids ex vivo as well as in vivo by intra-vital microscopy fluorescence [16–18]. However, ex vivo analysis of Mk localization in murine BM demonstrated that these cells can be located in the different anatomical portion and associated with endosteum, arterioles and sinusoids niche of the BM [5, 19]. The main Mk function within BM vascular niche is platelet production. During this process a giant endomitotic Mk converts into thousands platelets through a highly controlled formation of long, branching protrusions, designated proplatelets, comprised of platelet-sized swellings connected by thin cytoplasmatic bridges [14, 20]. The proplatelet model of platelet biogenesis has been identified both in vitro as well as in vivo [21, 22]. Nevertheless, how proplatelets are extended through sinusoidal endothelial cells and signals that regulate terminal Mk maturation remain poorly understood. Importantly, it is known that positioning of Mks in close proximity of BM sinusoids relies on chemokines that are released by several BM cell types, such as endothelial and perivascular stromal/mesenchymal cells. Stromal-Derived Factor-1 (SDF-1) was the first chemokine implicated in Mk redirection to the vascular niche [23]. Although produced by both endosteal osteoblasts and vascular endothelial cells, SDF-1 seems to regulate Mk maturational chemotaxis through engagement of its specific receptor CXCR4 (also known as CD186) [24]. Moreover, the elegant work by Avecilla and colleagues definitely assessed the importance of chemokines, including SDF-1 and fibroblast growth factor-4 (FGF-4) in restoring thrombopoiesis Thrombopoietin−/− (Thpo−/−) and Myeloproliferative Leukemia Protein−/− (Mpl−/−) mice by enhancing vascular cell adhesion molecule-1 (VCAM-1)- and very late antigen-4 (VLA-4)-mediated localization of CXCR4+ Mks to the vascular districts, thus promoting their survival, maturation and platelet release [25]. Consistently, both administration of SDF-1 and stabilization of endogenous SDF-1 were demonstrated to increase Mk-vasculature association and thrombopoiesis [26]. In particular, in the setting of radiation injury, dynamic fluctuations in marrow SDF-1 distribution spatially and temporally correlate with variations in Mk vascular niche positioning [26]. Recently, also angiogenic signals have been involved in Mk relocation to the vascular niche. A novel pro-inflammatory vascular endothelial growth factor receptor 1 (VEGFR1)-mediated pathway was seen to stimulate the maturation and up-regulation of CXCR4 on Mks, leading to their redistribution to sinusoids, thereby enhancing platelet production in vivo [27]. Therefore, the picture of Mks as static vascular single cells within BM vascular districts has evolved in dynamically and maturing cells that may change their location under specific stimuli, as, for example, during regenerative hemopoiesis. Despite this understanding, still little is known about the exact role of Mks in the BM beside being platelet progenitor cells.
Here we will review what is known about the reciprocal interaction of Mks with the BM environment in terms of cell function, BM homeostasis and related pathologies.
Mk interaction with the different cellular components of the bone marrow environment
While migrating between BM endosteal and vascular niches, Mks interact with different cellular components. Several studies have demonstrated that Mks establish interactions with both stromal lineages arising from the mesenchymal stem cells as well as lineages of hemopoietic origin. In some cases, these interactions are supported by strong in vivo evidences and come to light as reciprocal regulatory connections, while others emerged as weak interactions that have been demonstrated only in vitro. Therefore, in the next section of this review we will discuss the most studied Mk-BM cell interaction divided on the basis of their belonging to the BM connective or hemopoietic tissues (Table 1).
Mk-BM stromal cell interactions
Mesenchymal stem cells
Mesenchymal stem cells (MSCs) are a rare population of multipotent stem cells that constitute an important component of the hemopoietic niche in the BM. They are clonogenic, self-renewing cells that can differentiate into tissues of mesodermal origin, in particular osteoblasts (OBs), adipocytes and chondrocytes [28]. MSCs are able to recapitulate and organize a functional hemopoietic niche when transplanted in heterotopic sites [29], underling their importance as regulators of BM physiology and homeostasis.
The contribution of Mks to MSCs in vivo is an unexplored field. On the contrary, many evidences implicated MSCs as important regulator of Mk function. Several studies showed that MSCs express and secrete a broad array of cytokines and soluble factors [30, 31], among these Interleukin-6 (IL-6), Interleukin-11 (IL-11), Stem Cell Factor (SCF) and Leukemia Inhibitory Factor (LIF) that represent important modulators of Mk development and maturation [32–35].
An early report published by Cheng et al. [36] showed that BM expanded MSCs are able to support megakaryocytic differentiation in vitro. In this paper the authors indirectly identified the association of MSCs and Mks in human BM using a co-purification assay. They observed Mk-like cells expressing the CD41 marker in MSCs cultures isolated from BM and assumed that if MSCs and Mks are in direct contact inside the BM niche, it would be possible to co-purify them. Indeed, by using anti-CD41 conjugated magnetic beads, they were able to observe the appearance of fibroblast-like colonies after 2 weeks of culture in MSC medium, while just few colonies were obtained using a control IgG or an anti-CD14 antibody. Moreover when co-cultured with CD34+ cells from BM, MSCs were able to maintain hemopoietic progenitors and support Mk differentiation in absence of exogenous cytokines. Interestingly proplatelet formation was also observed in the co-cultures and platelet-sized particles expressing CD41 and CD62 and capable of a partial activation were recovered from the supernatant. Cheng et al. found also that expanded MSCs expressed Thpo transcript, but were not able to measure the secreted protein in the supernatant, suggesting that other factors may be involved.
A more recent study performed by our group further confirmed these results. To reproduce the cellular interactions inside the BM osteoblastic niche, we differentiated human Mks on confluent monolayers of MSCs or MSC-derived OBs. Though both the co-culture systems yielded the same amount of Mks, a significant difference in the maturation levels was observed, with a higher number of large, polyploid cells in the MSCs/Mks co-cultures and an increased proplatelet formation compared to the OBs/Mks co-cultures [15].
These data suggest that MSC interaction plays an important role in an advanced stage of Mk development rather than acting on early progenitors. In addition to secreting cytokines, MSCs can directly interact with hemopoietic cells and Mks through the expression of cell adhesion molecules like Intercellular Adhesion Molecule-1 (ICAM-1) and 2, Vascular Cell Adhesion Molecule-1 (VCAM-1) and E-Selectin [37]. Indeed, we confirmed that BM MSCs express VCAM-1 and demonstrated that antibod- mediated inhibition of VLA-4/VCAM-1 binding in MSCs/MKs co-cultures resulted in a decreased proplatelets formation compared to IgG-treated controls. However, since our system was dependent on IL-6, IL-11 and Thpo administration, it was not possible to determine the contribution of MSCs secreted factors to Mk maturation [15].
It is hence conceivable that direct cell/cell interaction and paracrine-secreted cytokines act in concert to assist Mk terminal differentiation and platelet production inside the BM niche. Some evidences reinforcing these results come from in vivo studies of MSCs/HSCs co-transplantation.
In 2003 Angelopoulou et al. [38] demonstrated that co-transplantation of human-derived MSCs and HSCs in NOD/SCID mice reinforced myelopoiesis and megakaryocytopoiesis. The authors showed that MSCs were able to enhance HSCs engraftment into irradiated mice and skew the hemopoiesis toward myeloid and megakaryocytic lineages. This was particularly evident when HSCs were transplanted at a limiting dose and resulted in a higher percentage of human Mks in the transplanted animal’s BM and an increased number of circulating platelets.
In addition to their role in regulating Mk maturation, MSCs can direct their migration inside the BM by secreting SDF-1. This chemokine is produced in the BM by endothelial cells, OBs and perivascular mesenchymal progenitors cells. The latter include CXCL12-abundant reticular (CAR) cells and Nestin+ stromal cells, two subtypes of early mesenchymal progenitors cells that retain the potential to differentiate toward OBs and adipocytes in vitro [9, 39].
All these evidences underlie a fundamental role for MSCs in orchestrating Mk development and function within the BM, providing the cellular, chemical and physical stimuli to support their terminal maturation and migration.
Osteoblasts/osteoclasts
Bone remodelling dynamics result from the activity of two distinct cell types, osteoblasts (OBs), that represents the bone forming cells and osteoclasts (OCs) that resorb bone. Many in vitro and in vivo evidences clearly stated a role of Mks in modulating the replication and differentiation of both those cells, with significant outcomes for human pathologies. Specifically, four different mouse models, characterized by megakaryocytosis and osteosclerosis, indicated a positive effect of the megakaryocytic lineage in bone cell dynamics [40–48]. Further, in vitro studies demonstrated that Mks express many factors responsible of OC development inhibition (Osteoprotegerin (OPG), RANKL, Transforming Growth Factor-β1 (TGF-β1), Granulocyte Monocyte-Colony Stimulating Factor (GM-CSF), IL-10, IL-13) [49–58] as well as factors known to modulate osteoblastogenesis and bone formation [49, 59, 60]. The role of Mks on OBs appears dual as they were demonstrated to enhance both their proliferation and differentiation [47, 50, 61]. Proliferation seemed to be mediated by cell–cell contacts, while more conflicting results were proposed about the role of Mks on OB differentiation. Direct cell–cell contacts, promoting Mk-mediated OB expansion, were dependent on the engagement of integrin signalling. To this regard, the fibronectin integrins α4β1 and α5β1 and the glycoprotein GPIIb (CD41) were demonstrated to play a critical role in Mk-induced OB proliferation, as assessed by neutralizing antibody studies [62]. Recently, Cheng et al. [63] demonstrated the importance of the non receptor tyrosine kinase Pyk2, a downstream mediator of integrin signalling, in Mk-induced OB proliferation. Although the increase in Pyk2 expression, obtained co-culturing Mks and OBs, was not related to β1 integrin activation, this up-regulation resulted critical for Mk-mediated OB expansion, both in vitro and in vivo, as demonstrated by the reduced responsiveness to Mk stimulation of Pyk2−/− OBs with respect to wild type. The importance of direct cell–cell interactions and β1 signaling in Mk-mediated proliferation of OBs were further confirmed by the same group in a recent work demonstrating a β1 dependent reduction of p53 and Rb cell cycle regulators upon Mk-OB interaction [64]. Moreover, direct contact between Mks and OBs in the regulation of skeletal homeostasis implies the communications via gap junctions. Gap junction-mediated intercellular communications were shown to be involved in the negative regulation of Mk-mediated OB proliferation by gap junction pharmacological inhibition [61].
Studies on the impact of Mks on OB differentiation were unexpectedly divergent. Ciovacco and colleagues, showed in vitro the inhibition of OB differentiation when co-cultured with Mks, in terms of type I collagen, osteocalcin and alkaline phosphatase expression and OB mineralization [65], Bord and colleagues showed, by in vitro studies, that Mks promoted OB differentiation as assessed by an increase in type I collagen and osteoprotegerin production [50]. However, divergences in these results may be ascribed to differences in culture timing. Interestingly, the increase in OB differentiation, described by Bord et al., was paralleled by a suppression of RANKL expression, highlighting an indirect negative effect of Mks on OC formation.
Mk-mediated effects on OBs appeared clearly dependent on cell–cell interactions, while the less characterized effects on OCs seemed to be mediated by the release of soluble factors, as demonstrated by the effect of Mk-conditioned media on OC culture. Specifically, Kacena et al. [55] demonstrated that Mks were able to release molecules with a molecular mass comprised between 10 and 50 kDa that inhibited OCs development and that seemed to be unrelated to the already known OC inhibitory factors (Interferon-γ (IFN-γ), TGF-β1, OPG, IL-4, IL-10).
While the Mk-mediated effects on osteoblastogenesis and osteoclastogenesis were independent on the developmental stage of megakaryocytic maturation, Mk number and longevity resulted critical for the outcomes on bone cells [65]. These results were confirmed in mouse models characterized by dysmegakaryopoiesis and osteosclerosis. As an example, mice overexpressing or repeatedly injected with the major Mk growth factor, Thpo, showed an important increase of Mk number at all stages of differentiation in the BM. The phenotype manifested in these mice was a myelofibrotic syndrome with associated osteosclerosis. Importantly, these mice presented high plasmatic levels of two Mk-secreted growth factors previously reported to influence bone cells, TGF-β1 and platelet-derived growth factor (PDGF) [40–44]. The involvement of these molecules in supporting an osteosclerotic phenotype was further confirmed by Chagroui et al. [66], who demonstrated that in the absence of TGF-β1, Thpo overexpressing mice failed to exhibit the myelofibrotic/osteosclerotic syndrome. The same group demonstrated that also overexpression of OPG was critically involved in the development of the osteosclerotic phenotype in Thpohigh mice, although no dependency between TGF-β and OPG secretion was found [67]. The Thpo/megakaryocytic lineage axis has been further claimed to be fundamental in osteoblastic niche expansion and thus in the engraftment of donor HSCs, in post total body irradiation (TBI). In this work Olson et al. [69], confirmed previous results of a SDF-1 dependent Mk relocation to the endosteal osteoblastic niche after TBI, which resulted concomitant to the marked expansion of endosteal OBs [68]. Furthermore, TBI-induced Mk migration was demonstrated to be dependent on MPL signalling and on the CD41-mediated adhesion, while OB proliferation required Mk-secreted PDGF-BB.
Two mouse models, extensively studied to elucidate Mk influence on bone cells homeostasis, were the GATA-1 and NF-E2 deficient mice [45–48, 70, 71]. GATA-1 and NF-E2 are key transcription factors in the regulation of Mk development as their genetic ablation in mice resulted in marked megakaryocytosis, paradoxically thrombocytopenia and a significant increase in bone mass density detectable after 4 months after birth. Although these mice exhibited high levels of OPG, the reduction in OC development and function was not associated to the high bone mass [47]. Nevertheless the increase of Mk number played a central role in new bone mass formation in GATA-1 and NF-E2 deficient mice. Further, the direct contact between Mks (either mutant or wild type) and OBs was required to induce OB proliferation [47]. Recently Meijome et al. [72], studied the influence of GATA-1−/− Mks in the rescue of bone alterations in OPG−/− mice, characterized by reduced trabecular and cortical bone due to increased osteoclastogenesis. This study demonstrated that, although GATA-1−/− × OPG−/− mice still present the osteoporosis of the cortical bone they rescue trabecular bone parameters, suggesting that Mks can locally enhance trabecular bone volume but that they cannot access cortical bone to inhibit osteoclastogenesis.
Finally, the Platelet Type-von Willebrand Disease (Pt-vWD) mouse model developed by Suva et al. [48], harboring a gain of function mutation of the specific von Willebrand Factor (vWF) receptor GpIbα, showed a phenotype that mirrored the human disease, including platelet dysfunction and impaired hemostasis. Further, these mutant mice presented thrombocytopenia, a spleen specific megakaryocytosis and a high bone mass phenotype detectable at two months after birth. In this model the bone alteration appeared to be due to the decrease in OC number with consequent bone breakdown that was attributed to the Mk/platelet dysfunction. However, the exact mechanism by which spleen megakaryocytosis affects osteoclastogenesis remains to be elucidated.
Adipocytes
Adipose tissue occupies considerable part of adult BM, but its role has been for long time controversial and poorly understood. Initially thought to be just a “filler” of the BM cavity, in the last years this position has been revisited and fat contribution to BM physiology and homeostasis has been brought to light. As reported by Naveiras et al. [73] the amount of adipose tissue in the BM inversely correlates with the presence of hemopoietic progenitors and active hemopoiesis. Using an engineered strain of “fatless” mice, the authors showed that BM engraftment and hemopoietic recovery after irradiation was faster compared to wild-type mice. The same effect was obtained by pharmacological inhibition of Peroxisome Proliferator-Activater Receptor-γ (PPAR-γ) in wild-type mice, collectively indicating a repressive effect of adipocytes on hemopoiesis.
Adipose tissue secretes a discrete number of cytokines and adipose-derived hormones that can influence the immune system activity and energy metabolism. In addition to IL-6, whose role in Mk differentiation is critical, other factors like adiponectin and leptin have been described to influence Mk development and function, though complete evidence is still missing. Two groups reported adiponectin receptor expression on platelets, Mks and megakaryocytic cell lines [74, 75]. Specifically, Kato et al. [74] observed accelerated thrombus formation in adiponectin knock-out (K.O.) mice after laser injury. This effect was reverted by adenovirus-mediated adiponectin restoration, indicating its possible antithrombotic role. A less defined function of adiponectin in Mk development has been described so far. Sun et al. [75] reported a decreased Mk differentiation and megakaryocytic colony formation in liquid or semisolid assays, respectively, when adiponectin was added to the media. In addition, in liquid assays this effect was evident only when adiponectin was added at the beginning of the culture, therefore, it is not possible to exclude an effect of this factor on early hemopoietic progenitors rather than on committed cells.
Leptin has been shown to enhance ADP-mediated platelet activation [76], but it’s not able to induce activation alone, indicating that it may act as a positive modulator of platelets function. The same effect was described in the megakaryoblastic cell line MEG-01 where leptin administration was shown to enhance ADP-induced cytosolic calcium (Ca2+) in a synergistic manner [77]. High adipokines doses, comparable to the one measured in obese patients were also shown to produce insulin resistance in megakaryocytic cell lines and primary cells [78].
Despite adipocytes have been demonstrated to be central player in hemopoiesis homeostasis, their specific contribution to Mk and other lineages differentiation is still unclear and new studies are required to get further insights.
Endothelial cells
Mks mainly reside in the vascular district within BM environment and platelet shedding is thought to happen by a direct extravasation of proplatelet through sinusoid endothelium [17]. Sinusoidal endothelium is uniquely specialized to support hemopoiesis and unlike other vessels, it is devoid of pericytes and surrounded by specialized SDF-1 secreting reticular cells. BM sinusoids can be specifically identified by their expression of Vascular Endothelial Growth Factor Receptor-3, while endothelial cells of arterioles uniquely express Sca-1 [79]. Many evidences have highlighted an important role of endothelial cells in the regulation of Mk functions, in terms of differentiation, development and release of platelets [23–25, 80, 81]. On the contrary, few studies have examined the functional role of Mks in the regulation of BM vasculature development or recovery, despite Mks represent an important reservoir of bioactive hemopoietic and angiogenic factors. Technical limitations have slow down these studies both in vivo and in vitro. As an example, long-term endothelial cultures have been established form different tissues, while BM endothelial cells have been isolated using different technique [82–85], but they always showed in vitro the phenotypic and functional features of microvascular cells. Further, endothelial cells and Mks are considered “sister cells”, both deriving from the common stem cell for hemopoietic/endothelial lineages, the hemangioblasts [86]. Therefore, co-expression of several developmental regulators has made difficult the investigation of their functional role in mediating Mk/endothelial cells interaction. However, in vitro studies have attempted to elucidate the mechanisms involved in Mk-endothelial cell contact. Mohle and colleagues demonstrated that ex vivo-generated human Mks isolated from BM express and synthesize VEGF and that, in co-culture with BM endothelial cells can maintain endothelial layer in serum-free medium [87]. Further, Thpo was demonstrated to induce the active release of VEGF by Mks in vitro, confirming a potential role of Mks in supporting angiogenesis [88]. Recently, human Mks were shown to foster cell differentiation and colony formation of endothelial progenitor cell (EPCs) [89]. Moreover, in vivo models clarified some aspects of this connection in conditions of regenerative hemopoiesis. In 2006 Kopp et al. [90] showed that thrombospondins (TSP) act as an important angiogenic switch. Generation of TSP1 and TSP2 double KO mice (TSP-Double KO) demonstrated that these mice were characterized by a faster recovery of BM vasculature after myelosuppression than wild-type mice. Further, TSP-DKO Mks were more proficient in supporting the generation of new vessel within Matrigel with respect to wild-type Mks. Finally, evidences of Mk-endothelial cell interdependence come from patients affected from prolonged isolated thrombocytopenia (PT), a serious complication after allogeneic HSC transplantation characterized by lower Mk content within BM. Patients BM exhibited remarkable decreases in cellular elements of the BM vascular microenvironment, including endothelial cells and peri-vascular cells [91]. Even though endothelial cells and Mks are closely associated within BM sinusoids in vivo, yet precise mechanisms of interactions are still missing.
Mk-hemopoietic tissue interactions
Hemopoietic stem cells
Previous research on the cellular components that contribute to the regulation of the HSC niches has been focused on the role of OBs, reticular stromal cells, endothelial cells and nerve cells [92]. However, the hypothesis that Mks can directly regulate HSCs was confirmed in several studies demonstrating that Mks produce and secrete significant amount of IL-6, which has been shown to dramatically expand spleen colony forming unit in vivo [56, 93]. Furthermore, Kirouac et al. [94] proposed that the self-renewal of umbilical cord blood-derived HSCs is regulated by a coupled positive–negative intercellular circuit composed of Mk-derived stimulatory growth factors (VEGF, PDGF, Epidermal Growth Factor and serotonin) and monocyte-derived inhibitory factors.
Nilsson’s group was the first to explore the possible role of Mks in maintaining mature HSCs in their niche. They found, by both in vivo and in vitro approaches, that transplanted HSCs preferentially lodge within two cells of mature Mks in murine BM and that HSCs isolated from the mice endosteum showed a significant increase in cell proliferation while in co-culture with mature sorted Mks. Furthermore, this endosteal HSC progeny was shown to retain HSC potential and to maintain long-term multi-lineage reconstitution capacity when transplanted into lethally ablated recipients. The mechanisms of increased HSC proliferation was not Mk contact dependent, but could be recapitulated with media supplemented with two factors identified in Mk-conditioned media: insulin-like growth factor binding protein-3 (IGFBP-3) and insulin-like growth factor-1 (IGF-1) [19]. The role of Mk as HSC-regulating niche cells was further confirmed by Zhao and colleagues [95]. In this study the authors showed that Mks are the major source of TGF-β1in the BM and maintain HSC quiescence under homeostatic conditions through TGF-β signaling. In particular, authors generated a Pf4-cre induced DTR (encoding diphtheria toxin receptor) mice (Pf4-cre; iDTR) in which Mks are rendered sensitive to diphtheria toxin. Removal of Mks after diphtheria toxin injection resulted in the increase of absolute numbers of short-term HSCs and multipotent progenitor cells. On the contrary, under conditions of chemotherapeutic conditions, when rapid expansion of HSCs is required, Mks were demonstrated to promote HSC expansion through FGF-1 production. Conditional deletion of FGF-1 in Mks in the Pf4-cre +; Fgfr1 flox/flox mice led to reduced Mk expansion and HSC regeneration after 5-fluorouracil challenge. Contemporary, Bruns and colleagues, intercrossed Cxcl4-cre mice with iDTR animals. Also in this mouse model, selective in vivo Mks depletion using inducible diphtheria toxin receptor expression resulted in specific loss of HSC quiescence with a marked expansion of functional HSCs. However, in this work Mk impact on HSC function was attributed to Mk expression of a regulator of HSC cell cycle activity, CXCL4, also known as Platelet Factor 4 (Pf4) [96].
Plasma cells
Long-lived Antibody-secreting Plasma cells (PCs) represents an example of mature hemopoietic cells that exhibit their own niche within BM. They are critical for long-term protection post-infection or immunization and persist for decades throughout the body, namely at sites of inflammation, but predominantly in the BM compartment [97]. A complex orchestration guides these cells to exit the germinal centers of lymphoid organs and migrate toward the BM, a privileged location where they receive additional signals required for their final differentiation and prolonged survival [98]. There is strong evidence that the longevity of BM PCs is dependent on environmental factors [99]. Different cells type displayed preferential contacts with PC and expressed putative PC survival factors, such as OBs, reticular cells and monocytes [100, 101]. The finding that only approximately 30 % of BM PCs colocalize with Mks suggested a transient contact between these mature cells and led Winter and colleagues to explore the possibility of Mks as cells producing important factors, such as IL-6 and APRIL (a proliferation inducing agent), in supporting PC growth and survival [102]. The authors demonstrated that injection of Thpo increased megakaryopoiesis and allowed the accumulation and persistence of a larger number of PCs generated in the course of a specific immune response in wild-type mice. On the contrary, c-mpl −/− mice, which exhibited reduced megakaryopoiesis, showed an impaired PC response in the BM. Together, these results demonstrate that Mks constitute a functional component of the PC niche in this tissue, thus establishing a novel link between PC memory and megakaryopoiesis.
Monocytes/macrophages
Few experimental evidences have highlighted a possible interaction between these two mature cell lineages deriving from the HSC. Data available suggest a prevalence of macrophage effects on thrombopoiesis rather than a reciprocal interaction. Landoni et al. [103] suggested that BM and spleen macrophages-derived signaling regulates negatively the Mk compartment as liposomal-encapsulated clodronate treatment provoked enhancement of megakaryopoiesis characterized by a dose-dependent increase in the number of BM and spleen Mks that was inversely correlated to the macrophage count in these organs. Furthermore, higher dose of liposomal-encapsulated clodronate induces enhanced thrombopoiesis in mice as demonstrated by an increased frequency of reticulated platelets and an improvement in the total platelet count two days later. D’Atri et al. [104] have recently showed in vitro that resting macrophages release soluble factors that sustain Mk growth, cell ploidy, a size increase, proplatelet production, and platelet release. Specifically, stimulation of macrophages with lipopolysaccharide triggered the secretion of cytokines that exerted opposite effects together with a dramatic switch of CD34+ commitment to the megakaryocytic lineage toward the myeloid lineage. Those effects seemed to be IL-8-dependent as neutralization of this cytokine partially reversed the inhibition of Mk growth.
Mk contribution of soluble factors and extracellular matrices within BM environment
Beyond cell–cell contacts, hemopoiesis is regulated by soluble factors and extracellular matrix components [105, 106]. Interestingly, mature Mks in the BM, present three different types of secretory granules in their cytoplasm: the alpha-granules, dense granules and lysosomes [107]. While dense granules predominantly contain serotonin, adenosine diphosphate (ADP) and polyphosphates, and lysosomes are fulfilled of enzymes, α-granules comprise more than eight hundred proteins, including hemostatic factors (vWF), angiogenic factors (VEGF), anti-angiogenic factors (endostatin, Platelet Factor 4), growth factors (EGF, PDGF, FGF, SDF-1α), metalloproteinases (MMP2, MMP9), necrotic factors (TNFα, TNFβ) and Extracellular Matrix components [108]. Delivery of these growth factors and cytokines represent the most intuitive way by means of which Mk might support and regulate cell function around the BM niches. However, while secretion of platelet granules is a key event in normal response to vascular damage in physiology, the mechanisms of Mk release of granule content are less clear. In general, what we know on Mk granule release come from studies on platelets and only hypothesis can be formulated on the ability of Mks to set an exocytic control of the external environment. Therefore, several questions still need to be addressed such as: are all α-granule cargoes, synthesized by Mks, completely delivered to nascent platelets in vivo or some of them are actively released to finely regulate the surrounding environment by an autocrine/paracrine manner? Which mechanisms are exploited by Mks to control the kinetics of granule release? Are differentially packaged α-granule cargoes released during physiologic hemopoiesis or in conditions of BM injury? Despite this paucity of knowledge, in 2008 Italiano and colleagues demonstrated that angiogenic and anti-angiogenic proteins were segregated into different sets of α-granules in Mks and platelets and that the selective release of those granules led to the regulation of angiogenesis [109]. Similar mechanisms were identified for the coagulation factors vWF and fibrinogen [110] as well as for inhibitors of matrix MMPs [111]. Moreover, pro-fibrotic factors of Mk α-granule derivation, such as TGF-β and PDGF, are critically involved in the onset of BM diseases and will be discussed in depth in the next sections of this review. Beside cytokines and growth factors, an intriguing research field concerns the mechanisms of ECM rearrangement by Mks both in physiological conditions and disease. Growing evidences demonstrated that Mks differently interact with the BM extracellular environment during their maturation and positioning in the BM niche [112]. ECM components constitute a dynamic structure that is continuously degraded and deposited. Interestingly, Mks possess collagenase activity, as they express several MMPs, such as MMP-2, MMP-9, MMP-14, MMP-24, MMP-25 [113], assemble podosomes that rearrange the ECM through proteolytic activity [114] and express several ECM components such as fibronectins, laminin isoforms, type IV collagen, nidogen and entactin [5, 115, 116]. In addition, our group has recently demonstrated that expression of many of those ECMs is not related to the physiological production of platelets but is increased concomitantly with the regeneration of BM environment following myelosuppression [5].
The BM extracellular microenvironment is composed of structural fibrils and extracellular matrix components. The most common structural fibrils in the BM are collagen, reticulin, laminin and fibronectin [4, 5, 117–119], embedded in a matrix of glycosaminoglycans and glycoproteins, most of which have not been well characterized. Moreover, a wide variety of benign and malignant disorders are characterized by an increase in BM stromal fibers [120]. These fibers are commonly composed solely of reticulin fibers but may also include collagen fibers. Immunohistochemical detection of reticulin with silver-based stains and collagen with trichrome stains represent routine stains performed on BM biopsy specimens in diagnostic laboratories. Two slightly different, but widely supported grading scales for these stains have been created [121, 122]. The original Bauermeister scheme [121] has been simplified into a five grade system, while, the more recent Thiele scale includes only four categories [122]. Both of these grading scales take into account both the type of fibers seen (reticulin or collagen) and the overall amount of fibrosis. Interestingly, when an increase in BM reticulin staining does occur, it is usually accompanied by an increase in BM Mks, often morphologically atypical.
The contribution of Mks to BM fibrosis remains only partially understood and it is likely that cytokines, growth factors, cross-linking enzymes and ECMs might all be involved. The pivotal role of Mks in the onset of BM fibrosis associated with hematological malignancies will be discussed in the following section of this review. However, new features of Mks as pro-fibrotic cells have emerged. As an example, a linkage between BM fibrosis and lysyl oxidase (LOX) derived from low ploidy Mks has been recently described [123]. LOX is a copper-dependent enzyme that cross-links collagen or elastin by oxidative deamination of peptidyl lysine or hydroxylysine and peptidyl lysine residues, respectively, and contributes to the accumulation of ECMs by promoting intrapeptide and interpeptide chain cross-linking [124]. Interestingly, LOX is highly expressed in low ploidy, proliferating Mks, while mature Mks have a scarce expression of this enzyme. Consistently, LOX is also abundant in the GATA-1low mouse model with pathologically high levels of low-ploidy Mks associated with an extensively fibrotic matrix. Inhibition of LOX enzymatic activity in GATA-1low mice by β-aminopropionitrile (BAPN), inhibited the progression of myelofibrosis, demonstrating for the first time the potential role of LOX derived from low ploidy Mks in sustaining BM fibrosis [123].
Megakaryocytes and diseases
Germline mutations in genes encoding for different proteins relevant for the regulation of Mk biology have been described to dramatically affect platelet count, thus resulting in human diseases known as inherited thrombocytopenias. Several different forms have been identified so far and classified according to the presence or absence of other inherited defects associated with the low peripheral blood platelet numbers [125]. The clinical phenotype of these patients can vary widely depending on the type of mutation. In some cases the pathology is recognized early in infancy, for example in Jacobsen syndrome, the thrombocytopenia is associated with physical growth retardation and characteristic facial dysmorphism, while congenital amegakaryocytic thrombocytopenia always evolves into BM aplasia. However, in other forms, the clinical phenotype becomes evident in adulthood, such as in thrombocytopenia associated with sitosterolemia (STSL), which may display anemia, tendon xanthomas and atherosclerosis [126].
Thrombocytopenia is the only common feature of all inherited thrombocytopenias, thus indicating that, regardless of funding mutation, any alteration in proteins involved in the control of specific Mk function may result in defective platelet formation. Our knowledge of the cellular mechanisms of altered platelet production in many of these pathologies has greatly expanded in recent years due to the opportunity to culture human Mks differentiated from patients’ hemopoietic progenitor cells in vitro. To this regard, we have demonstrated that Mks from MYH9-Related Disease patients, carrying mutations in the MYH9 gene encoding for the heavy chain of non-muscle myosin IIA, show a significant defect in proplatelet branching and completely lose the physiologic suppression of proplatelet extension exerted by interaction with type I collagen, thus suggesting that a premature platelet release within BM may contribute to the appearance of thrombocytopenia [127, 128]. Moreover, in heterozygous Bernard-Soulier syndrome type Bolzano, mutations of the membrane glycoprotein GPIbα determine a profound reduction in Mk ability to form proplatelet on different ECM components, such as fibrinogen and vWF, with altered distribution of cytoskeletal tubulin and increased size of proplatelet tips, consistent with the increased diameters of peripheral blood platelets [129]. Consistently, Mks from patients with the homozygous Bernard-Soulier syndrome are completely unable to extend proplatelets in vitro [130]. The key role for membrane glycoproteins in regulating Mk interaction with components of the extracellular environment that sustain platelet formation is further confirmed by the finding that a constitutive activation of αIIbβ3-mediated outside-in signaling negatively influences proplatelet formation and severely impairs cell adhesion because of altered actin distribution and consequent defective stress fibers assembly [131]. The relevance of cytoskeleton contractility in the regulation of platelet formation and released is also evident in a recently described macrothrombocytopenia caused by mutations in the alpha-actinin 1 gene (ACTN1), that occurs within the actin-binding domain of alpha-actinin 1 that cross-links actin filaments into bundles [132]. In vitro transfection of Mks with disease-associated ACTN1 variants causes a disorganized actin-based cytoskeleton that is responsible for the abnormal proplatelet formation with proplatelet branches displaying few large tips [133]. Beside cytoskeleton and glycoproteins, a fine regulation of biochemical signaling is also important to ensure proper platelet formation. In familial thrombocytopenia 2 (THC2) point mutations in the 5′ UTR of ankyrin repeat domain 26 (ANKRD26) are responsible for the constitutive expression of this gene in the late stages of Mk maturation, when is normally silenced [134]. This leads to increased signaling via the Thpo pathway that in turn is responsible for impaired proplatelet formation. Interestingly, the main signaling molecule which appears to be deregulated is the Mitogen Activated Protein Kinase Extracellular Related Kinase (ERK), given that its selective inhibition completely rescues the in vitro defect [134]. Importantly, a feature of ANKRD26 Mks is the presence of a large amounts of PaCSs, cytoplasmic structures with selective immunoreactivity for polyubiquitinated proteins and proteasome, normally detected in solid cancers and their pre-neoplastic lesions, suggesting a link of these structures with oncogenesis [135]. Interestingly, ANKRD26 patients are exposed to an increased risk of leukemia, but the mechanisms explaining the relationship between dysmegakaryopoiesis and leukemia have not been yet investigated.
Overall, these mutations specifically affect proplatelet formation, the final stage of Mk differentiation, while preserving HSC commitment, polyploidization and maturation, thus suggesting that Mk participation in BM homeostasis is not significantly influenced. One exception is the gray-platelet syndrome (GPS), in which α-granules are reduced or absent in mature Mks because of mutation in the NBEAL2 gene [136]. The associated bleeding syndrome is usually mild to moderate, but can be also severe. In most patients thrombocytopenia is accompanied by high risk of developing evolutive myelofibrosis and splenomegaly with a profound change of BM architecture that dramatically compromise hemopoiesis [137]. Mks usually appear highly vacuolated; but their number is often normal and all the other organelles are normally present [138]. The major hypothesis for the development of BM fibrosis is that proteins stored inside Mk α-granule, such as PDGF and TGF-β1, are normally synthesized, but spontaneously released from Mk cytoplasm into the marrow because of deregulated storage, thus promoting ECM synthesis and deposition. Interestingly, a GPS murine model was recently shown to recapitulate the typical platelet phenotype observed in patients including splenomegaly and myelofibrosis [139]. This model demonstrated that expression of macrophage inflammatory protein (MIP) 1α and 1β is significantly increased in GPS Mks. These well-known pro-inflammatory chemokines are increased also in primary myelofibrosis, a myeloproliferative disease characterized by BM fibrosis and splenomegaly as well. Actually, beside inherited mutations, it has been described that also numerous somatic genetic alterations, usually acquired by the HSCs, may affect Mk differentiation and platelet formation [140, 141]. Specifically, the appearance of a mutate hemopoietic clone, that can escape the proliferation arrest, leading to the abnormal accumulation of BM Mks, is the typical feature of polycythemia vera (PV), essential thrombocythemia (ET) and primary myelofibrosis (PMF), three diseases belonging to the family of the Philadelphia-negative classical myeloproliferative neoplasms (MPNs) [142–145]. BM from PV patients presents neoplastic proliferation and maturation of erythroid, megakaryocytic and granulocytic elements. Mks are increased in number and display characteristic morphological abnormalities, such as hyperlobated nuclei. Histology of BM from ET patients usually reveals marked Mk hyperplasia. There are frequently giant polyploidy Mks, but the overall morphology is fairly normal. Mks from ET patients have an increased tendency to form high-branched proplatelet, in vitro [146]. Consistently, ET is often discovered incidentally in asymptomatic patients because of the abnormally high platelet count, which may be cause of thrombotic complications. BM biopsies of individuals with PMF are characterized by important Mk hyperplasia and dysplasia, with hypolobated and cloud-like Mk nuclei. Platelet count may differ between the onset and the progression of the disease, from high to low numbers [147], and also platelet formation has been described to present several structural alterations in vitro [146]. Only PMF patients show increased deposition of reticulin and collagen fibers. A condition that progressively drives reactive BM fibrosis, which in turn is the major responsible for the final BM failure, with consequent splenomegaly because of the appearance of extramedullary hemopoiesis. A myelofibrotic phenotype may be secondary to ET or to an early/prefibrotic myelofibrosis state, characterized by the absence of relevant reticulin fibrosis in BM, but with evident Mk proliferation and dysplasia [148]. The close link among these different diseases is not surprising, given the evidence that they all share the same molecular pathogenesis, even if for a little percentage of patients the causing mutation remains still elusive. Nearly 95 % of PV and 50 % of ET and PMF present a somatic point mutation in the gene encoding for the Janus Kinase-2 (JAK2) (JAK2V617F). This genetic alteration, which was the first to be characterized, is selectively acquired by the myeloid compartment, and leads to production of a protein with kinase activity that is constitutively activated [149, 150]. JAK/STAT signaling, together with Phosphatidylinositol-4,5-bisphosphate 3-kinase (PI3 K)/Akt and Ras/ERK pathways, are the major determinant of Thpo-mediated control of megakaryopoiesis and Mk polyploidisation upon interaction with its receptor, c-Mpl [151, 152]. It has been shown that enhanced activation of JAK/STAT signaling promotes HSC proliferation and contributes to render them hypersensitive to Thpo [153]. However, why a single mutation contributes to different clinical outcomes is still unknown. One of the possible explanations is the difference in the allelic burden [154, 155]. Recently, different knock-in mouse models of the mutant JAK2V617F, have been created. Interestingly, all of them developed an MPN that is closely reminiscent of human PV. However, loss of wild-type JAK2 allele in homozygous JAK2V617F mice results in markedly increased white blood cells, neutrophils, reticulocytes and platelets in the peripheral blood, together with accelerated BM fibrosis and significantly larger spleen size, suggesting that loss of wild-type cells increases the severity of the MPN [156]. However, it is becoming clear that although HSCs harboring JAK2V617F mutation promote MPN, additional components maybe necessary for disease progression. To this regard, it has been recently described that loss of the Thpo receptor significantly ameliorates MPN development in JAK2V617F transgenic mice, thus suggesting that c-Mpl expression is a fundamental trigger of MPNs development [157]. Moreover, JAK2 mutations may occur in ET or PMF patient in concomitance with Lymphocyte Specific Protein (LNK) mutations [158–160], which can be acquired either early or late during the clonal evolution of the disease [161]. Loss-of function mutations of LNK lead to further activation of JAK/STAT signalling given that the protein normally down-regulates this pathway upon c-Mpl activation [162–164]. Of note, mice with a complete loss of LNK display many features in common with human MPN, including splenomegaly, thrombocytosis and extramedullary hemopoiesis [162, 163, 165].
The importance of Thpo signaling in the pathogenesis of MPNs is confirmed by the description of activating mutations in gene encoding for c-Mpl in about 10 % of JAK2-negative ET and PMF patients, but not PV [166–168]. c-Mpl mutation is significantly associated with thrombocytosis [169, 170], and severe anemia [171], and cells form these patients display spontaneous formation of Mk colonies [172, 173]. Mice engineered to permanently express high doses of Thpo, through retroviral infection, develop a myeloproliferative syndrome, characterized by hyperproliferative Mks and thrombocytosis, with associated spleen and BM fibrosis, while injections of high doses of Thpo have been used to induce in mice and rats a myelofibrotic phenotype associated with thrombocytosis and increased number of BM Mks [42, 44, 174, 175]. Consistent with these evidences, therapeutic treatment with Thpo mimetics in humans, beside the rescue of platelet count to normal levels in thrombocytopenic patients, may determine the development of BM fibrosis as side effect [176, 177].
The last identified somatic mutations responsible for about 25 % of all PMF and ET, and some JAK2-negative PV, involves the gene encoding for calreticulin, CALR [178–180]. CALR mutations are mutually exclusive with mutations in both JAK2 and c-Mpl [178, 179], indicating that CALR is the second most frequently mutated gene in myeloproliferative neoplasms. CALR, is a multifunctional protein that binds with high-capacity Ca2+ ions that must be restrained into the endoplasmic reticulum (ER) [181]. All the mutations that have been described so far, affect a region of the gene that encodes for the C-terminal peptide containing a negatively charged calcium-binding domain, and the ER retention motif. The resulting mutant proteins share a novel amino acid sequence containing positively charged amino acids, thus probably affecting Ca2+ binding capacity, and loose the reticulum retention motif, suggesting possible protein dislocation. All patients with mutated CALR display thrombocytosis, but thrombosis-free survival is significantly longer compared with other mutations [178, 179]. Recently, a fine regulation of intracellular Ca2+ signaling has been described as a major trigger of both proplatelet formation and Mk interaction with ECM components that sustain Mk motility, through the activation of the mechanism of store operated Ca2+ entry (SOCE) [182]. These data, together with the finding that Mks express the highest amount of mutated CALR among BM cells [183], suggest that altered Ca2+ homeostasis may contribute to this phenotype. Interestingly, it has been shown that CALR is actively involved in the modulation of SOCE dynamics [184, 185], and mice that present constitutive activation of SOCE, exclusively in BM Mks, resemble many of the alterations of PMF patients, especially fibrosis and splenomegaly [186]. Moreover, it has been demonstrated that the same mechanisms is up-regulate in human endothelial cells from PMF patients [187]; however insight into the role of Ca2+ signaling in human Mk in the pathogenesis of PMF and ET are currently missing. Importantly, in an effort to try to understand the pathogenic mechanisms promoted by CALR mutations, it has been demonstrated that, as in the case of JAK2 and c-Mpl mutations, JAK/STAT signaling is over-activated in these patients [178, 188]. An important finding that, taken together with data from JAK2 and c-Mpl mutated patients, definitively suggests that deregulated JAK/STAT signaling is a common future of nearly all PMF and ET regardless of founding driver mutation and clinical diagnosis [188].
A driving role for Thpo signaling in affecting BM homeostasis is confirmed also in acute megakaryoblastic leukemia (AMKL), another disease with impaired Mk maturation and a profound alteration of BM organization due to variable degree of fibrosis [141, 189, 190]. AMKL affects three major subgroups of patients: adults (adult AMKL), infants with down syndrome (DS-AMKL), and children without DS (non-DS paediatric AMKL). Interestingly, nearly all patients with AMKL present thrombocytopenia, although some may have thrombocytosis. Morphologically, BM histopathology shows the presence of uniform blasts, mainly of the Mk lineage, with presence of mature dysplastic Mks and only few residual foci of granulo and erythropoiesis. Constitutive activation of signaling associated with JAK/STAT cascade has been described in a significant proportion of adult AMKL, thus suggesting that, as in the case of myelofibrosis, alterations of biochemical pathways relevant for Mk differentiation result in AMKL phenotype. The molecular basis for the constitutive activation of this signaling cascades is due to mutations in genes encoding for c-Mpl and JAK [190, 191]. Moreover, several studies reported that activating mutations in Fms-like tyrosine kinase 3 (FLT3), as well as c-KIT, are associated with adult AMKL [192, 193].
Interestingly, activating mutations in FLT3, JAK2, JAK3 and c-Mpl can be present also in child with trisomy 21 in DS-AMKL [141, 194, 195]. However, the major cause of DS-AMKL is due to acquired somatic mutations in the hemopoietic transcription factor gene GATA1 preventing the synthesis of a full-length protein, but allowing the synthesis of a shorter variant (GATA-1 s) with altered trans-activation capacity [191, 196] that has been described to increase the proliferation of Mk progenitors and to represses final Mk differentiation [197, 198]. Interestingly, GATA-1low mice model usually develop a hematological picture that features human myelofibrosis [46, 59, 199] with Mk progenitors exhibiting striking hyper-proliferative rates, both in vivo and in vitro [200]. This phenotype is almost similar to that observed in wild-type mice subject to high concentration of Thpo [201]. Actually, mice treated with Thpo show a decrease of GATA-1 content in Mks, thus further strongly supporting the strict relationship between Mk and extrinsic (forced expression of Thpo, Thpohigh mice) or intrinsic (reduced GATA-1 expression, GATA-1low mice) factors in the development of myelofibrosis [201]. Conversely, expression of a mutant GATA-1 s protein in a knock-in mouse model of the disease is able to induce developmental stage-specific defects in yolk sac and fetal liver Mk progenitors, but is not sufficient to induce AMKL per se [197]. Therefore, this genetic mutation may not be sufficient to induce AMKL. Consistently, most DS-AMKL patients with constitutional trisomy 21 and GATA1 mutations present a transient myeloproliferative disorder at birth, with spontaneous remission and absence of further malignant disease in most instances [200] and humans with inherited mutant alleles fail to develop leukemia [202]. These observations indicate that there are multigenic contributions to the development of AMKL. To this regard it has been demonstrated that trisomy 21 alone provides a proliferative stimulus of fetal megakaryopoiesis and erythropoiesis [203]. In fact, cells harboring trisomy 21 overexpress the transcription regulator ERG which strongly cooperates with the GATA1 s mutated protein and interferes with Akt signaling molecule to immortalize Mk progenitors, thus contributing to aberrant megakaryopoiesis [204]. Accordingly, a fine regulation of Akt activation is required to guarantee an efficient platelet production [205, 206], and defective regulation of this pathway is associated with abnormal Mk proliferation [206].
Together, all these data suggest that, independently from the founding mutation, any signals that impair c-Mpl downstream transduction cascades may result in alteration of BM Mks with a significant tendency to develop fibrosis. But how aberrant megakaryopoieisis can be linked with the BM fibrosis which finally causes the ineffective hemopoiesis? Besides BM a variety of organs including lung, liver, kidney, intestine, heart, skin may develop fibrotic disease. It is currently assumed that in all of these organs the underlying mechanism of fibrotic reactions, leading to progressive organ imbalance, involves the over-stimulation of resident stromal cells, namely mesenchymal cells and myofibroblast, to produce and deposit collagens and other ECM components. The major cytokines known to have a role in mediating the fibrotic process are PDGF and TGF-β1 [207, 208].
TGF-β1 is the first member identified of pleiotropic cytokines involved in normal tissue repair and development. Although expressed by all cell types, is particularly abundant in platelets. TGF-β1 activity is regulated by proteases that convert the latent complex into an active form, thus inducing the expression of both extracellular matrix components (collagen type I and fibronectin) and their cognate receptors on target cells and its sustained production induces fibrosis in numerous organs, including myelofibrosis [66, 209].
The role of TGF-β1 and its downstream signaling are suggested by the evidence that development of myelofibrosis in wild-type mice treated with high doses of Thpo is associated with high TGF-β1 content in extracellular fluids of both BM and spleen [201], and that Gata1low mice Mks show increased TGF-β1 content [210]. Moreover, TGF-β1 deficient mice over-expressing Thpo are fibrosis resistant as compared to wild-type mice [66], and inhibition of TGF-β1 signaling is able to rescue the phenotype of Gata1low mice, reducing fibrosis/neovascularization/osteogenesis and increasing hemopoiesis in BM with its concomitant reduction within spleen [210]. In parallel to these evidences, it has been also reported that Mk from PMF and AMKL patients synthesize and release higher amount of both latent and bioactive TGF-β1 [211–214], and that this behavior is highly correlated with BM fibrosis grade [213].
Moreover, PDGF is a potent mitogen for cells of mesenchymal origin, including myofibroblasts; [215, 216]. Interestingly, PDGF expression is increased in BM Mks from patients with myeloproliferative disorders [217] and PMF patients display a significant association between BM fibrosis and aberrant expression of PDGF within BM [218]. Moreover, AMKL Mks secrete higher amount of PDGF and the highest PDGF secretion is associated with more severe fibrosis [219]. Thus, it is possible to assume that aberrant pro-fibrotic cytokines released by Mks of patients with BM fibrosis may be the major determinant for the appearance of this phenotype.
Both PDGF and TGF-β1 are stored in Mk α-granules and are constitutively released by BM Mks during the course of their physiological maturation [49, 220, 221]. Actually, it has been demonstrated that both these cytokines may exert an autocrine effect on Mks by regulating both megakaryopoiesis and platelet production [212, 222, 223]. Further, Thpo has been shown to promote α-granules release in human platelets [224]. Platelets and Mks share the same activating mechanisms, thus is seems reasonable to hypothesize that an over-stimulation of JAK/STAT signaling, as a consequence of the different driving alterations described, may be a major determinant of fibrosis by promoting the release of Mk pro-fibrotic cytokines that acting on stromal cells induces the excessive collagens and ECM components production.
Perspectives and concluding remarks
Research in recent years has made many progress in understanding the unique process of proplatelet formation and platelet shedding by Mks. Despite all these efforts, little is known about the signals that regulate these events and the interactions of Mks with the different components of the BM environment. In this context, the most recent research has ascribed to Mks and platelets new and variegated functions beside their roles in platelet production and hemostasis. Mks can sense different biological and physical stimuli and regulate different cellular processes accordingly. As an example, Mks can participate in angiogenesis by modulating and secreting their granule content [109], in bone formation and in matrix reorganization by releasing soluble factors, ECMs and MMPs [68, 113, 225]. All these discoveries prompt a central role of Mks in the regulation of BM homeostasis in steady state and “stressed” hemopoiesis as well as in the development of pathologies, such as myeloproliferative neoplasms, where alterations in Mk morphology and function are associated to BM fibrosis (Fig. 1). To this regard there are still two major questions to be answered: how the synthesis of ECM components by Mks is regulated? and in which conditions this process is activated? It is known that both ECM composition and structure in the BM affect Mk function and platelet release. Moreover, in vivo Mks are surrounded by a peri-cellular matrix mainly composed by type IV collagen, laminin and fibronectin with a fibrillar organization [5]. These ECM components may complex with growth factors and regulate their activity or constitute themselves autocrine regulatory factors by activating biochemical and/or mechanical signals upon binding to their receptors [225, 226]. Alterations of these processes, like in BM fibrosis, may lead to impaired modulation of the different Mk functions with one function prevailing on the others (e.g. Mk proliferation observed in PMF patient BM biopsies) [144]. How Mk and stromal-derived ECM components interact and reciprocally regulate their expression and function are still to be understood. As an example, we have demonstrated that Mks release and organize fibronectin upon adhesion to type I collagen and not fibrinogen through activation of the transglutaminase factor XIII [225]. Interestingly, we have recently demonstrated that intracellular calcium signaling is fundamental for Mk interactions with the different ECM components of the BM [182]. These data find an obvious link with the newly discovered mutation in CALR in myeloproliferative neoplasm [178, 179]. Here alterations in Ca2+ signaling may lead to abnormal interaction of Mks with BM ECM components with consequences in Mk functions and matrix homeostasis and remodeling. Interestingly, both Mk interaction with the BM environment and proplatelet formation are powered by cytoskeleton forces [227]. Specifically, the Rho-Rock-myosin axis regulates Mk contractility, interaction with type I collagen in the BM and inhibition of proplatelet formation [128]. Activation of this axis is dependent on the environment stiffness as demonstrated by the recovery of proplatelet formation on softer type I collagen [228, 229]. Thus, the reciprocal regulation of Mks and BM is also dependent on the mechanics of both cells and environment. Similarly, proplatelet formation occurs in the proximity of BM sinusoids and is regulated by the coordinate activity of different factors of that district. Here ECM is mainly composed by softer and non-fibrillar components (e.g., type IV collagen, fibronectin) that are known to promote both Mk differentiation and proplatelet formation [112]. Proplatelet elongation depends on microtubule remodeling mainly driven by dynein-facilitated microtubule sliding [230].

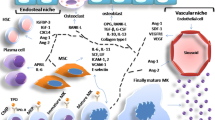
Schematic representation of Mk contribution to the bone marrow environment in a bone marrow steady state b bone marrow fibrosis and injury
On the top of these mechanisms a reciprocal regulation exists between Mks and all the other cellular components of the BM environment. Some interactions are well known, as for example Mks and OB/OC reciprocal regulation, and some are still to be completely understood, as the link between endothelial cells, adypocytes or monocytes/macrophages and Mks. All these are important points to be clarified as it is important to understand how Mks extend proplatelets through the vascular wall, as well as how Mk differentiation is regulated by the different composition in BM adypocytes that occurs during aging or diseases and, finally, how inflammation may impact platelet release.
Research on BM has been impaired by technical difficulties in obtaining an intact organ without bone decalcification in mouse models or BM biopsies in human. Multiphoton intravital microscopy has been used to visualize platelet production in vivo in murine BM [17, 18]. Although this live imaging technique has strengthened the physiological role of proplatelet formation during both steady state thrombopoiesis or thrombocytopenic conditions, it raised new questions on the roles of hydrodynamic forces in platelet morphogenesis or on the events that lead to proplatelet morphogenesis in peripheral blood.
For this reason, lately the study has been focused on the creation of ex vivo BM models to extrapolate new insight on BM function in vivo. To this regard we have recently proposed a 3D model, entirely made by silk biomaterial, in which we have recapitulated the vascular niche environment for functional human platelet production [231]. By this model we believe to be able to dissect the role of the different cellular, molecular and physical components of the BM in directing platelet production and, in reverse order, the role of Mks in modulating BM homeostasis. The advantage of our model is the use of silk biomaterial. Silk is a naturally derived, biocompatible and tunable biomaterial with outstanding optical properties [232], which can be prepared in a range of material formats and processed entirely in aqueous systems allowing the incorporation of labile compounds without loss of bioactivity [233]. Silk biomaterial has been proven to have non-thrombogenic features, while supporting functional platelet production. The importance of 3D tissue systems has grown substantively in recent years, moving from 2D and towards more complex and relevant biomaterials. A few years ago Matsunaga et al. [234] proposed a 3D system for platelet production using a three-phase culture system. More recently, different 3D models were described for hematopoietic stem cell migration and proliferation using co-culture systems [235] and for platelet production [231, 236]. Lately, different models have been developed in the attempt of mimicking the bone marrow niches to study hemopoiesis [237], to support platelet production by iPS cell-derived Mks [238].
In conclusion, Mks actively interact with the different components of the BM both physically and molecularly. By this connection both platelet production and BM homeostasis are regulated. Many aspects of these fascinating processes have still to be clarified and this will be possible only by joining different efforts and expertise in a multidisciplinary approach.
We apologize to those whose work was not included in this review due to space limitations.
References
Fliedner TM et al (1985) Bone marrow structure and its possible significance for hematopoietic cell renewal. Ann N Y Acad Sci 459:73–84
Li XM, Hu Z, Jorgenson ML, Slayton WB (2009) High levels of acetylated low-density lipoprotein uptake and low tyrosine kinase with immunoglobulin and epidermal growth factor homology domains-2 (Tie2) promoter activity distinguish sinusoids from other vessel types in murine bone marrow. Circulation 120(19):1910–1918
Smaniotto S et al (2013) Mouse basophils reside in extracellular matrix-enriched bone marrow niches which control their motility. PLoS One 8(9):e70292
Nilsson SK et al (1998) Immunofluorescence characterization of key extracellular matrix proteins in murine bone marrow in situ. J Histochem Cytochem 46(3):371–377
Malara A et al (2014) Megakaryocytes contribute to the bone marrow-matrix environment by expressing fibronectin, type IV collagen, and laminin. Stem Cells 32(4):926–937
Lo Celso C et al (2009) Live-animal tracking of individual haematopoietic stem/progenitor cells in their niche. Nature 457(7225):92–96
Arai F et al (2004) Tie2/angiopoietin-1 signaling regulates hematopoietic stem cell quiescence in the bone marrow niche. Cell 118(2):149–161
Kiel MJ, Yilmaz OH, Iwashita T, Terhorst C, Morrison SJ (2005) SLAM family receptors distinguish hematopoietic stem and progenitor cells and reveal endothelial niches for stem cells. Cell 121(7):1109–1121
Mendez-Ferrer S et al (2010) Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 466(7308):829–834
Calvi LM et al (2003) Osteoblastic cells regulate the haematopoietic stem cell niche. Nature 425(6960):841–846
Greenbaum A et al (2013) CXCL12 in early mesenchymal progenitors is required for haematopoietic stem-cell maintenance. Nature 495(7440):227–230
Kunisaki Y et al (2013) Arteriolar niches maintain haematopoietic stem cell quiescence. Nature 502(7473):637–643
Hanoun M, Frenette PS (2013) This niche is a maze; an amazing niche. Cell Stem Cell 12(4):391–392
Hartwig J, Italiano J (2003) The birth of the platelet. J Thromb Haemost 1(7):1580–1586
Pallotta I, Lovett M, Rice W, Kaplan DL, Balduini A (2009) Bone marrow osteoblastic niche: a new model to study physiological regulation of megakaryopoiesis. PLoS One 4(12):e8359
Becker RP, De Bruyn PP (1976) The transmural passage of blood cells into myeloid sinusoids and the entry of platelets into the sinusoidal circulation; a scanning electron microscopic investigation. Am J Anat 145(2):183–205
Junt T et al (2007) Dynamic visualization of thrombopoiesis within bone marrow. Science 317(5845):1767–1770
Kowata S et al (2014) Platelet demand modulates the type of intravascular protrusion of megakaryocytes in bone marrow. Thromb Haemost 112(4):743–756
Heazlewood SY et al (2013) Megakaryocytes co-localise with hemopoietic stem cells and release cytokines that up-regulate stem cell proliferation. Stem Cell Res 11(2):782–792
Thon JN et al (2010) Cytoskeletal mechanics of proplatelet maturation and platelet release. J Cell Biol 191(4):861–874
Leven RM (1987) Megakaryocyte motility and platelet formation. Scanning Microsc 1(4):1701–1709
Tablin F, Castro M, Leven RM (1990) Blood platelet formation in vitro. The role of the cytoskeleton in megakaryocyte fragmentation. J Cell Sci 97((Pt 1)):59–70
Hamada T et al (1998) Transendothelial migration of megakaryocytes in response to stromal cell-derived factor 1 (SDF-1) enhances platelet formation. J Exp Med 188(3):539–548
Wang JF, Liu ZY, Groopman JE (1998) The alpha-chemokine receptor CXCR4 is expressed on the megakaryocytic lineage from progenitor to platelets and modulates migration and adhesion. Blood 92(3):756–764
Avecilla ST et al (2004) Chemokine-mediated interaction of hematopoietic progenitors with the bone marrow vascular niche is required for thrombopoiesis. Nat Med 10(1):64–71
Niswander LM, Fegan KH, Kingsley PD, McGrath KE, Palis J (2014) SDF-1 dynamically mediates megakaryocyte niche occupancy and thrombopoiesis at steady-state and following radiation injury. Blood 124(2):277–286
Pitchford SC, Lodie T, Rankin SM (2012) VEGFR1 stimulates a CXCR4-dependent translocation of megakaryocytes to the vascular niche, enhancing platelet production in mice. Blood 120(14):2787–2795
Pittenger MF et al (1999) Multilineage potential of adult human mesenchymal stem cells. Science 284(5411):143–147
Sacchetti B et al (2007) Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoietic microenvironment. Cell 131(2):324–336
Haynesworth SE, Baber MA, Caplan AI (1996) Cytokine expression by human marrow-derived mesenchymal progenitor cells in vitro: effects of dexamethasone and IL-1 alpha. J Cell Physiol 166(3):585–592
Majumdar MK, Thiede MA, Mosca JD, Moorman M, Gerson SL (1998) Phenotypic and functional comparison of cultures of marrow-derived mesenchymal stem cells (MSCs) and stromal cells. J Cell Physiol 176(1):57–66
Broudy VC, Lin NL, Kaushansky K (1995) Thrombopoietin (c-mpl ligand) acts synergistically with erythropoietin, stem cell factor, and interleukin-11 to enhance murine megakaryocyte colony growth and increases megakaryocyte ploidy in vitro. Blood 85(7):1719–1726
Navarro S et al (1991) Interleukin-6 and its receptor are expressed by human megakaryocytes: in vitro effects on proliferation and endoreplication. Blood 77(3):461–471
Ishibashi T et al (1989) Human interleukin 6 is a direct promoter of maturation of megakaryocytes in vitro. Proc Natl Acad Sci USA 86(15):5953–5957
Metcalf D, Hilton D, Nicola NA (1991) Leukemia inhibitory factor can potentiate murine megakaryocyte production in vitro. Blood 77(10):2150–2153
Cheng L, Qasba P, Vanguri P, Thiede MA (2000) Human mesenchymal stem cells support megakaryocyte and pro-platelet formation from CD34 + hematopoietic progenitor cells. JCP 184(1):58–69
Majumdar MK et al (2003) Characterization and functionality of cell surface molecules on human mesenchymal stem cells. J Biomed Sci 10(2):228–241
Angelopoulou M et al (2003) Cotransplantation of human mesenchymal stem cells enhances human myelopoiesis and megakaryocytopoiesis in NOD/SCID mice. Exp Hematol 31:413–420
Sugiyama T, Kohara H, Noda M, Nagasawa T (2006) Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity 25(6):977–988
Frey BM, Rafii S, Crystal RG, Moore MA (1998) Adenovirus long-term expression of thrombopoietin in vivo: a new model for myeloproliferative syndrome and osteomyelofibrosis. Schweiz Med Wochenschr 128(42):1587–1592
Frey BM et al (1998) Adenovector-mediated expression of human thrombopoietin cDNA in immune-compromised mice: insights into the pathophysiology of osteomyelofibrosis. J Immunol 160(2):691–699
Yan XQ et al (1995) Chronic exposure to retroviral vector encoded MGDF (mpl-ligand) induces lineage-specific growth and differentiation of megakaryocytes in mice. Blood 86(11):4025–4033
Yan XQ et al (1996) A model of myelofibrosis and osteosclerosis in mice induced by overexpressing thrombopoietin (mpl ligand): reversal of disease by bone marrow transplantation. Blood 88(2):402–409
Villeval JL et al (1997) High thrombopoietin production by hematopoietic cells induces a fatal myeloproliferative syndrome in mice. Blood 90(11):4369–4383
Shivdasani RA et al (1995) Transcription factor NF-E2 is required for platelet formation independent of the actions of thrombopoietin/MGDF in megakaryocyte development. Cell 81(5):695–704
Shivdasani RA, Fujiwara Y, McDevitt MA, Orkin SH (1997) A lineage-selective knockout establishes the critical role of transcription factor GATA-1 in megakaryocyte growth and platelet development. EMBO J 16(13):3965–3973
Kacena MA et al (2004) Megakaryocyte-osteoblast interaction revealed in mice deficient in transcription factors GATA-1 and NF-E2. J Bone Miner Res 19(4):652–660
Suva LJ et al (2008) Platelet dysfunction and a high bone mass phenotype in a murine model of platelet-type von Willebrand disease. Am J Pathol 172(2):430–439
Wickenhauser C et al (1995) Detection and quantification of transforming growth factor beta (TGF-beta) and platelet-derived growth factor (PDGF) release by normal human megakaryocytes. Leukemia 9(2):310–315
Bord S et al (2005) Megakaryocytes modulate osteoblast synthesis of type-l collagen, osteoprotegerin, and RANKL. Bone 36(5):812–819
Bord S et al (2004) Synthesis of osteoprotegerin and RANKL by megakaryocytes is modulated by oestrogen. Br J Haematol 126(2):244–251
Bord S, Ireland DC, Beavan SR, Compston JE (2003) The effects of estrogen on osteoprotegerin, RANKL, and estrogen receptor expression in human osteoblasts. Bone 32(2):136–141
Pearse RN et al (2001) Multiple myeloma disrupts the TRANCE/osteoprotegerin cytokine axis to trigger bone destruction and promote tumor progression. Proc Natl Acad Sci USA 98(20):11581–11586
Chagraoui H et al (2003) Expression of osteoprotegerin mRNA and protein in murine megakaryocytes. Exp Hematol 31(11):1081–1088
Kacena MA et al (2006) Megakaryocyte-mediated inhibition of osteoclast development. Bone 39(5):991–999
Jiang S et al (1994) Cytokine production by primary bone marrow megakaryocytes. Blood 84(12):4151–4156
Soslau G, Morgan DA, Jaffe JS, Brodsky I, Wang Y (1997) Cytokine mRNA expression in human platelets and a megakaryocytic cell line and cytokine modulation of platelet function. Cytokine 9(6):405–411
Wickenhauser C et al (1995) Secretion of cytokines (interleukins-1 alpha, -3, and -6 and granulocyte-macrophage colony-stimulating factor) by normal human bone marrow megakaryocytes. Blood 85(3):685–691
Vannucchi AM et al (2002) Development of myelofibrosis in mice genetically impaired for GATA-1 expression (GATA-1(low) mice). Blood 100(4):1123–1132
Sipe JB et al (2004) Localization of bone morphogenetic proteins (BMPs)-2, -4, and -6 within megakaryocytes and platelets. Bone 35(6):1316–1322
Ciovacco WA et al (2009) The role of gap junctions in megakaryocyte-mediated osteoblast proliferation and differentiation. Bone 44(1):80–86
Lemieux JM, Horowitz MC, Kacena MA (2010) Involvement of integrins alpha(3)beta(1) and alpha(5)beta(1) and glycoprotein IIb in megakaryocyte-induced osteoblast proliferation. J Cell Biochem 109(5):927–932
Cheng YH et al (2013) Pyk2 regulates megakaryocyte-induced increases in osteoblast number and bone formation. J Bone Miner Res 28(6):1434–1445
Cheng YH et al (2014) Signaling pathways involved in megakaryocyte-mediated proliferation of osteoblast lineage cells. J Cell Physiol 230(3):578–586
Ciovacco WA, Cheng YH, Horowitz MC, Kacena MA (2010) Immature and mature megakaryocytes enhance osteoblast proliferation and inhibit osteoclast formation. J Cell Biochem 109(4):774–781
Chagraoui H et al (2002) Prominent role of TGF-beta 1 in thrombopoietin-induced myelofibrosis in mice. Blood 100(10):3495–3503
Chagraoui H et al (2003) Stimulation of osteoprotegerin production is responsible for osteosclerosis in mice overexpressing TPO. Blood 101(8):2983–2989
Dominici M et al (2009) Restoration and reversible expansion of the osteoblastic hematopoietic stem cell niche after marrow radioablation. Blood 114(11):2333–2343
Olson TS et al (2013) Megakaryocytes promote murine osteoblastic HSC niche expansion and stem cell engraftment after radioablative conditioning. Blood 121(26):5238–5249
Kacena MA, Gundberg CM, Nelson T, Horowitz MC (2005) Loss of the transcription factor p45 NF-E2 results in a developmental arrest of megakaryocyte differentiation and the onset of a high bone mass phenotype. Bone 36(2):215–223
Shivdasani RA, Fielder P, Keller GA, Orkin SH, de Sauvage FJ (1997) Regulation of the serum concentration of thrombopoietin in thrombocytopenic NF-E2 knockout mice. Blood 90(5):1821–1827
Meijome TE et al (2014) GATA-1 Deficiency rescues trabecular but not cortical bone in opg deficient mice. J Cell Physiol 230(4):783–790
Naveiras O et al (2009) Bone-marrow adipocytes as negative regulators of the haematopoietic microenvironment. Nature 460(7252):259–263
Kato H et al (2006) Adiponectin acts as an endogenous antithrombotic factor. Arterioscler Thromb Vasc Biol 26(1):224–230
Sun S et al (2013) Expression of plasma membrane receptor genes during megakaryocyte development. Physiol Genomics 45(6):217–227
Nakata M, Yada T, Soejima N, Maruyama I (1999) Leptin promotes aggregation of human platelets via the long form of its receptor. Diabetes 48(2):426–429
Nakata M, Maruyama I, Yada T (2005) Leptin potentiates ADP-induced [Ca(2 +)](i) increase via JAK2 and tyrosine kinases in a megakaryoblast cell line. Diabetes Res Clin Pract 70(3):209–216
Gerrits AJ et al (2012) Induction of insulin resistance by the adipokines resistin, leptin, plasminogen activator inhibitor-1 and retinol binding protein 4 in human megakaryocytes. Haematologica 97(8):1149–1157
Hooper AT et al (2009) Engraftment and reconstitution of hematopoiesis is dependent on VEGFR2-mediated regeneration of sinusoidal endothelial cells. Cell Stem Cell 4(3):263–274
Rafii S et al (1995) Human bone marrow microvascular endothelial cells support long-term proliferation and differentiation of myeloid and megakaryocytic progenitors. Blood 86(9):3353–3363
Rafii S, Mohle R, Shapiro F, Frey BM, Moore MA (1997) Regulation of hematopoiesis by microvascular endothelium. Leuk Lymphoma 27(5–6):375–386
Irie S, Tavassoli M (1986) Purification and characterization of rat bone marrow endothelial cells. Exp Hematol 14(10):912–918
Fei RG, Penn PE, Wolf NS (1990) A method to establish pure fibroblast and endothelial cell colony cultures from murine bone marrow. Exp Hematol 18(8):953–957
Masek LC, Sweetenham JW (1994) Isolation and culture of endothelial cells from human bone marrow. Br J Haematol 88(4):855–865
Delia D et al (1993) CD34 expression is regulated reciprocally with adhesion molecules in vascular endothelial cells in vitro. Blood 81(4):1001–1008
Choi K, Kennedy M, Kazarov A, Papadimitriou JC, Keller G (1998) A common precursor for hematopoietic and endothelial cells. Development 125(4):725–732
Möhle R, Green D, Moore MA, Nachman RL, Rafii S (1997) Constitutive production and thrombin-induced release of vascular endothelial growth factor by human megakaryocytes and platelets. Proc Natl Acad Sci USA 94(2):663–668
Bobik R, Hong Y, Breier G, Martin JF, Erusalimsky JD (1998) Thrombopoietin stimulates VEGF release from c-Mpl-expressing cell lines and haematopoietic progenitors. FEBS Lett 423(1):10–14
Kwon SM et al (2014) Cross Talk with Hematopoietic cells regulates the endothelial progenitor cell differentiation of cd34 positive cells. PLoS One 9(8):e106310
Kopp HG et al (2006) Thrombospondins deployed by thrombopoietic cells determine angiogenic switch and extent of revascularization. J Clin Invest 116(12):3277–3291
Kong Y et al (2014) Association between an impaired bone marrow vascular microenvironment and prolonged isolated thrombocytopenia after allogeneic hematopoietic stem cell transplantation. Biol Blood Marrow Transplant 20(8):1190–1197
Yamazaki S et al (2011) Nonmyelinating Schwann cells maintain hematopoietic stem cell hibernation in the bone marrow niche. Cell 147(5):1146–1158
Suzuki C et al (1989) Continuous perfusion with interleukin 6 (IL-6) enhances production of hematopoietic stem cells (CFU-S). Biochem Biophys Res Commun 159(3):933–938
Kirouac DC et al (2010) Dynamic interaction networks in a hierarchically organized tissue. Mol Syst Biol 6:417
Zhao M et al (2014) Megakaryocytes maintain homeostatic quiescence and promote post-injury regeneration of hematopoietic stem cells. Nat Med 20(11):1321–1326
Bruns I et al (2014) Megakaryocytes regulate hematopoietic stem cell quiescence through CXCL4 secretion. Nat Med 20(11):1315–1320
Tew JG et al (1992) Germinal centers and antibody production in bone marrow. Immunol Rev 126:99–112
Kallies A et al (2004) Plasma cell ontogeny defined by quantitative changes in blimp-1 expression. J Exp Med 200(8):967–977
Sze DM, Toellner KM, García de Vinuesa C, Taylor DR, MacLennan IC (2000) Intrinsic constraint on plasmablast growth and extrinsic limits of plasma cell survival. J Exp Med 192(6):813–821
Tokoyoda K, Egawa T, Sugiyama T, Choi BI, Nagasawa T (2004) Cellular niches controlling B lymphocyte behavior within bone marrow during development. Immunity 20(6):707–718
Belnoue E et al (2008) APRIL is critical for plasmablast survival in the bone marrow and poorly expressed by early-life bone marrow stromal cells. Blood 111(5):2755–2764
Winter O et al (2010) Megakaryocytes constitute a functional component of a plasma cell niche in the bone marrow. Blood 116(11):1867–1875
Landoni VI (2004) Macrophage derived signalling regulates negatively the megakaryocyte compartment. Cell Mol Biol (Noisy-le-grand) 50:OL667–OL675
D’Atri LP et al (2011) Paracrine regulation of megakaryo/thrombopoiesis by macrophages. Exp Hematol 39(7):763–772
Clark BR, Keating A (1995) Biology of bone marrow stroma. Ann N Y Acad Sci 770:70–78
Moore KA (2004) Recent advances in defining the hematopoietic stem cell niche. Curr Opin Hematol 11(2):107–111
Coppinger JA, Maguire PB (2007) Insights into the platelet releasate. Curr Pharm Des 13(26):2640–2646
Zufferey A et al (2014) Characterization of the platelet granule proteome: evidence of the presence of MHC1 in alpha-granules. J Proteomics 101:130–140
Italiano JE et al (2008) Angiogenesis is regulated by a novel mechanism: pro- and antiangiogenic proteins are organized into separate platelet alpha granules and differentially released. Blood 111(3):1227–1233
Sehgal S, Storrie B (2007) Evidence that differential packaging of the major platelet granule proteins von Willebrand factor and fibrinogen can support their differential release. J Thromb Haemost 5(10):2009–2016
Villeneuve J et al (2009) Tissue inhibitors of matrix metalloproteinases in platelets and megakaryocytes: a novel organization for these secreted proteins. Exp Hematol 37(7):849–856
Balduini A et al (2008) Adhesive receptors, extracellular proteins and myosin IIA orchestrate proplatelet formation by human megakaryocytes. J Thromb Haemost 6(11):1900–1907
Lane WJ et al (2000) Stromal-derived factor 1-induced megakaryocyte migration and platelet production is dependent on matrix metalloproteinases. Blood 96(13):4152–4159
Schachtner H et al (2013) Megakaryocytes assemble podosomes that degrade matrix and protrude through basement membrane. Blood 121(13):2542–2552
Schick PK, Wojensk CM, Bennett V, Denisova L (1996) Fibronectin isoforms in megakaryocytes. Stem Cells 14(Suppl 1):212–219
Nigatu A et al (2006) Megakaryocytic cells synthesize and platelets secrete alpha5-laminins, and the endothelial laminin isoform laminin 10 (alpha5beta1gamma1) strongly promotes adhesion but not activation of platelets. Thromb Haemost 95(1):85–93
Bentley SA, Alabaster O, Foidart JM (1981) Collagen heterogeneity in normal human bone marrow. Br J Haematol 48(2):287–291
Reilly JT, Nash JR, Mackie MJ, McVerry BA (1985) Immuno-enzymatic detection of fibronectin in normal and pathological haematopoietic tissue. Br J Haematol 59(3):497–504
Cattoretti G, Schiró R, Orazi A, Soligo D, Colombo MP (1993) Bone marrow stroma in humans: anti-nerve growth factor receptor antibodies selectively stain reticular cells in vivo and in vitro. Blood 81(7):1726–1738
Kuter DJ, Bain B, Mufti G, Bagg A, Hasserjian RP (2007) Bone marrow fibrosis: pathophysiology and clinical significance of increased bone marrow stromal fibres. Br J Haematol 139(3):351–362
Bauermeister DE (1971) Quantitation of bone marrow reticulin–a normal range. Am J Clin Pathol 56(1):24–31
Thiele J et al (2005) European consensus on grading bone marrow fibrosis and assessment of cellularity. Haematologica 90(8):1128–1132
Eliades A et al (2011) Control of megakaryocyte expansion and bone marrow fibrosis by lysyl oxidase. J Biol Chem 286(31):27630–27638
Kagan HM, Li W (2003) Lysyl oxidase: properties, specificity, and biological roles inside and outside of the cell. J Cell Biochem 88(4):660–672
Balduini CL, Pecci A, Noris P (2013) Diagnosis and management of inherited thrombocytopenias. Semin Thromb Hemost 39(2):161–171
Balduini CL, Savoia A, Seri M (2013) Inherited thrombocytopenias frequently diagnosed in adults. J Thromb Haemost 11(6):1006–1019
Pecci A et al (2009) Megakaryocytes of patients with MYH9-related thrombocytopenia present an altered proplatelet formation. Thromb Haemost 102(1):90–96
Chen Z et al (2007) The May-Hegglin anomaly gene MYH9 is a negative regulator of platelet biogenesis modulated by the Rho-ROCK pathway. Blood 110(1):171–179
Balduini A et al (2009) Proplatelet formation in heterozygous Bernard-Soulier syndrome type Bolzano. J Thromb Haemost 7(3):478–484
Balduini A, Malara A, Balduini CL, Noris P (2011) Megakaryocytes derived from patients with the classical form of Bernard-Soulier syndrome show no ability to extend proplatelets in vitro. Platelets 22(4):308–311
Bury L, Malara A, Gresele P, Balduini A (2012) Outside-in signalling generated by a constitutively activated integrin αIIbβ3 impairs proplatelet formation in human megakaryocytes. PLoS One 7(4):e34449
Kunishima S et al (2013) ACTN1 mutations cause congenital macrothrombocytopenia. Am J Hum Genet 92(3):431–438
Guéguen P et al (2013) A missense mutation in the alpha-actinin 1 gene (ACTN1) is the cause of autosomal dominant macrothrombocytopenia in a large French family. PLoS One 8(9):e74728
Bluteau D et al (2014) Thrombocytopenia-associated mutations in the ANKRD26 regulatory region induce MAPK hyperactivation. J Clin Invest 124(2):580–591
Necchi V et al (2013) Ubiquitin/proteasome-rich particulate cytoplasmic structures (PaCSs) in the platelets and megakaryocytes of ANKRD26-related thrombo-cytopenia. Thromb Haemost 109(2):263–271
Bottega R et al (2013) Correlation between platelet phenotype and NBEAL2 genotype in patients with congenital thrombocytopenia and α-granule deficiency. Haematologica 98(6):868–874
Nurden AT, Nurden P (2007) The gray platelet syndrome: clinical spectrum of the disease. Blood Rev 21(1):21–36
Breton-Gorius J, Vainchenker W, Nurden A, Levy-Toledano S, Caen J (1981) Defective alpha-granule production in megakaryocytes from gray platelet syndrome: ultrastructural studies of bone marrow cells and megakaryocytes growing in culture from blood precursors. Am J Pathol 102(1):10–19
Guerrero JA, et al. (2014) Gray Platelet Syndrome: Pro-inflammatory megakaryocytes and α-granule loss cause myelofibrosis and confer resistance to cancer metastasis in mice. Blood
Barosi G, Lupo L, Rosti V (2012) Management of myeloproliferative neoplasms: from academic guidelines to clinical practice. Curr Hematol Malig Rep 7(1):50–56
Malinge S et al (2008) Activating mutations in human acute megakaryoblastic leukemia. Blood 112(10):4220–4226
Thiele J et al. (2008) Primary myelofibrosis. (Swerdlow SH, Campo E, Harris NL, Jaffee ES, Pileri SA, Stein H, Thiele J, Vardiman JW, ed., Lyon: IARC Press), pp 44–47
Michiels JJ (1997) Diagnostic criteria of the myeloproliferative disorders (MPD): essential thrombocythaemia, polycythaemia vera and chronic megakaryocytic granulocytic metaplasia. Neth J Med 51(2):57–64
Michiels JJ, Berneman Z, Schroyens W, De Raeve H (2014) Changing Concepts of diagnostic criteria of myeloproliferative disorders and the molecular etiology and classification of myeloproliferative neoplasms: from dameshek 1950 to vainchenker 2005 and beyond. Acta Haematol 133(1):36–51
Alvarez-Larrán A et al (2014) WHO-histological criteria for myeloproliferative neoplasms: reproducibility, diagnostic accuracy and correlation with gene mutations and clinical outcomes. Br J Haematol 166(6):911–919
Balduini A et al (2011) In vitro megakaryocyte differentiation and proplatelet formation in Ph-negative classical myeloproliferative neoplasms: distinct patterns in the different clinical phenotypes. PLoS One 6(6):e21015
Tefferi A et al (2014) CALR vs JAK2 vs MPL-mutated or triple-negative myelofibrosis: clinical, cytogenetic and molecular comparisons. Leukemia 28(7):1472–1477
Barosi G (2014) Essential thrombocythemia vs. early/prefibrotic myelofibrosis: why does it matter. Best Pract Res Clin Haematol 27(2):129–140
Kralovics R et al (2005) A gain-of-function mutation of JAK2 in myeloproliferative disorders. N Engl J Med 352(17):1779–1790
Baxter EJ et al (2005) Acquired mutation of the tyrosine kinase JAK2 in human myeloproliferative disorders. Lancet 365(9464):1054–1061
Kaushansky K (2009) Molecular mechanisms of thrombopoietin signaling. J Thromb Haemost 7(Suppl 1):235–238
Hitchcock IS, Kaushansky K (2014) Thrombopoietin from beginning to end. Br J Haematol 165(2):259–268
Seita J et al (2007) Lnk negatively regulates self-renewal of hematopoietic stem cells by modifying thrombopoietin-mediated signal transduction. Proc Natl Acad Sci USA 104(7):2349–2354
James C et al (2008) The hematopoietic stem cell compartment of JAK2V617F-positive myeloproliferative disorders is a reflection of disease heterogeneity. Blood 112(6):2429–2438
Anand S et al (2011) Effects of the JAK2 mutation on the hematopoietic stem and progenitor compartment in human myeloproliferative neoplasms. Blood 118(1):177–181
Tiedt R et al (2008) Ratio of mutant JAK2-V617F to wild-type Jak2 determines the MPD phenotypes in transgenic mice. Blood 111(8):3931–3940
Sangkhae V, Etheridge SL, Kaushansky K, Hitchcock IS (2014) The thrombopoietin receptor, MPL, is critical for development of a JAK2V617F-induced myeloproliferative neoplasm. Blood 124(26):3956–3963
Lasho TL, Pardanani A, Tefferi A (2010) LNK mutations in JAK2 mutation-negative erythrocytosis. N Engl J Med 363(12):1189–1190
Oh ST et al (2010) Novel mutations in the inhibitory adaptor protein LNK drive JAK-STAT signaling in patients with myeloproliferative neoplasms. Blood 116(6):988–992
Pardanani A et al (2010) LNK mutation studies in blast-phase myeloproliferative neoplasms, and in chronic-phase disease with TET2, IDH, JAK2 or MPL mutations. Leukemia 24(10):1713–1718
Lasho TL, Tefferi A, Finke C, Pardanani A (2011) Clonal hierarchy and allelic mutation segregation in a myelofibrosis patient with two distinct LNK mutations. Leukemia 25(6):1056–1058
Velazquez L et al (2002) Cytokine signaling and hematopoietic homeostasis are disrupted in Lnk-deficient mice. J Exp Med 195(12):1599–1611
Takaki S, Morita H, Tezuka Y, Takatsu K (2002) Enhanced hematopoiesis by hematopoietic progenitor cells lacking intracellular adaptor protein. Lnk. J Exp Med 195(2):151–160
Tong W, Lodish HF (2004) Lnk inhibits Tpo-mpl signaling and Tpo-mediated megakaryocytopoiesis. J Exp Med 200(5):569–580
Bersenev A et al (2010) Lnk constrains myeloproliferative diseases in mice. J Clin Invest 120(6):2058–2069
Pardanani AD et al (2006) MPL515 mutations in myeloproliferative and other myeloid disorders: a study of 1182 patients. Blood 108(10):3472–3476
Pikman Y et al (2006) MPLW515L is a novel somatic activating mutation in myelofibrosis with myeloid metaplasia. PLoS Med 3(7):e270
Chaligné R et al (2008) New mutations of MPL in primitive myelofibrosis: only the MPL W515 mutations promote a G1/S-phase transition. Leukemia 22(8):1557–1566
Vannucchi AM et al (2008) Characteristics and clinical correlates of MPL 515 W > L/K mutation in essential thrombocythemia. Blood 112(3):844–847
Beer PA et al (2008) MPL mutations in myeloproliferative disorders: analysis of the PT-1 cohort. Blood 112(1):141–149
Guglielmelli P et al (2007) Anaemia characterises patients with myelofibrosis harbouring Mpl mutation. Br J Haematol 137(3):244–247
Chaligné R et al (2007) Evidence for MPL W515L/K mutations in hematopoietic stem cells in primitive myelofibrosis. Blood 110(10):3735–3743
Li Y et al (1996) Proto-oncogene c-mpl is involved in spontaneous megakaryocytopoiesis in myeloproliferative disorders. Br J Haematol 92(1):60–66
Ulich TR et al (1996) Systemic hematologic effects of PEG-rHuMGDF-induced megakaryocyte hyperplasia in mice. Blood 87(12):5006–5015
Yanagida M et al (1997) The role of transforming growth factor-beta in PEG-rHuMGDF-induced reversible myelofibrosis in rats. Br J Haematol 99(4):739–745
Kuter DJ et al (2009) Evaluation of bone marrow reticulin formation in chronic immune thrombocytopenia patients treated with romiplostim. Blood 114(18):3748–3756
Kuter DJ (2009) Thrombopoietin and thrombopoietin mimetics in the treatment of thrombocytopenia. Annu Rev Med 60:193–206
Klampfl T et al (2013) Somatic mutations of calreticulin in myeloproliferative neoplasms. N Engl J Med 369(25):2379–2390
Nangalia J et al (2013) Somatic CALR mutations in myeloproliferative neoplasms with nonmutated JAK2. N Engl J Med 369(25):2391–2405
Broséus J, Park JH, Carillo S, Hermouet S, Girodon F (2014) Presence of calreticulin mutations in JAK2-negative polycythemia vera. Blood
Smith MJ, Koch GL (1989) Multiple zones in the sequence of calreticulin (CRP55, calregulin, HACBP), a major calcium binding ER/SR protein. EMBO J 8(12):3581–3586
Di Buduo CA et al (2014) The importance of calcium in the regulation of megakaryocyte function. Haematologica 99(4):769–778
Vannucchi AM et al (2014) Calreticulin mutation-specific immunostaining in myeloproliferative neoplasms: pathogenetic insight and diagnostic value. Leukemia 28(9):1811–1818
Llewelyn Roderick H, Llewellyn DH, Campbell AK, Kendall JM (1998) Role of calreticulin in regulating intracellular Ca2 + storage and capacitative Ca2 + entry in HeLa cells. Cell Calcium 24(4):253–262
Ma J, Pan Z (2003) Retrograde activation of store-operated calcium channel. Cell Calcium 33(5–6):375–384
Grosse J et al (2007) An EF hand mutation in Stim1 causes premature platelet activation and bleeding in mice. J Clin Invest 117(11):3540–3550
Dragoni S et al (2014) Enhanced expression of Stim, Orai, and TRPC transcripts and proteins in endothelial progenitor cells isolated from patients with primary myelofibrosis. PLoS ONE 9(3):e91099
Rampal R et al (2014) Integrated genomic analysis illustrates the central role of JAK-STAT pathway activation in myeloproliferative neoplasm pathogenesis. Blood 123(22):e123–e133
Barnard DR et al (2007) Comparison of childhood myelodysplastic syndrome, AML FAB M6 or M7, CCG 2891: report from the Children’s Oncology Group. Pediatr Blood Cancer 49(1):17–22
Malinge S, Izraeli S, Crispino JD (2009) Insights into the manifestations, outcomes, and mechanisms of leukemogenesis in Down syndrome. Blood 113(12):2619–2628
Wickrema A, Crispino JD (2007) Erythroid and megakaryocytic transformation. Oncogene 26(47):6803–6815
Fröhling S et al (2007) Identification of driver and passenger mutations of FLT3 by high-throughput DNA sequence analysis and functional assessment of candidate alleles. Cancer Cell 12(6):501–513
Fröhling S, Scholl C, Gilliland DG, Levine RL (2005) Genetics of myeloid malignancies: pathogenetic and clinical implications. J Clin Oncol 23(26):6285–6295
Khan I, Malinge S, Crispino J (2011) Myeloid leukemia in Down syndrome. Crit Rev Oncog 16(1–2):25–36
Walters DK et al (2006) Activating alleles of JAK3 in acute megakaryoblastic leukemia. Cancer Cell 10(1):65–75
Wechsler J et al (2002) Acquired mutations in GATA1 in the megakaryoblastic leukemia of Down syndrome. Nat Genet 32(1):148–152
Li Z et al (2005) Developmental stage-selective effect of somatically mutated leukemogenic transcription factor GATA1. Nat Genet 37(6):613–619
Kuhl C et al (2005) GATA1-mediated megakaryocyte differentiation and growth control can be uncoupled and mapped to different domains in GATA1. Mol Cell Biol 25(19):8592–8606
Shivdasani R (2002) An animal model for myelofibrosis. Blood 100(4):1109
Vyas P, Ault K, Jackson CW, Orkin SH, Shivdasani RA (1999) Consequences of GATA-1 deficiency in megakaryocytes and platelets. Blood 93(9):2867–2875
Vannucchi AM et al (2005) A pathobiologic pathway linking thrombopoietin, GATA-1, and TGF-beta1 in the development of myelofibrosis. Blood 105(9):3493–3501
Hollanda LM et al (2006) An inherited mutation leading to production of only the short isoform of GATA-1 is associated with impaired erythropoiesis. Nat Genet 38(7):807–812
Hoeller S et al (2014) Morphologic and GATA1 sequencing analysis of hematopoiesis in fetuses with trisomy 21. Hum Pathol 45(5):1003–1009
Stankiewicz MJ, Crispino JD (2013) AKT collaborates with ERG and Gata1 s to dysregulate megakaryopoiesis and promote AMKL. Leukemia 27(6):1339–1347
Balduini A et al (2012) Constitutively released adenosine diphosphate regulates proplatelet formation by human megakaryocytes. Haematologica 97(11):1657–1665
Currao M, Balduini CL, Balduini A (2013) High doses of romiplostim induce proliferation and reduce proplatelet formation by human megakaryocytes. PLoS One 8(1):e54723
Bonner JC (2004) Regulation of PDGF and its receptors in fibrotic diseases. Cytokine Growth Factor Rev 15(4):255–273
Pohlers D et al (2009) TGF-beta and fibrosis in different organs-molecular pathway imprints. Biochim Biophys Acta 1792(8):746–756
Blobe GC, Schiemann WP, Lodish HF (2000) Role of transforming growth factor beta in human disease. N Engl J Med 342(18):1350–1358
Zingariello M et al (2013) Characterization of the TGF-β1 signaling abnormalities in the Gata1low mouse model of myelofibrosis. Blood 121(17):3345–3363
Ciurea SO et al (2007) Pivotal contributions of megakaryocytes to the biology of idiopathic myelofibrosis. Blood 110(3):986–993
Badalucco S et al (2013) Involvement of TGFβ1 in autocrine regulation of proplatelet formation in healthy subjects and patients with primary myelofibrosis. Haematologica 98(4):514–517
Ponce CC, de Lourdes F, Chauffaille M, Ihara SS, Silva MR (2012) The relationship of the active and latent forms of TGF-β1 with marrow fibrosis in essential thrombocythemia and primary myelofibrosis. Med Oncol 29(4):2337–2344
Terui T et al (1990) The production of transforming growth factor-beta in acute megakaryoblastic leukemia and its possible implications in myelofibrosis. Blood 75(7):1540–1548
Heldin CH, Westermark B (1999) Mechanism of action and in vivo role of platelet-derived growth factor. Physiol Rev 79(4):1283–1316
Biernacka A, Dobaczewski M, Frangogiannis NG (2011) TGF-β signaling in fibrosis. Growth Factors 29(5):196–202
Katoh O, Kimura A, Itoh T, Kuramoto A (1990) Platelet derived growth factor messenger RNA is increased in bone marrow megakaryocytes in patients with myeloproliferative disorders. Am J Hematol 35(3):145–150
Bock O et al (2005) Aberrant expression of platelet-derived growth factor (PDGF) and PDGF receptor-alpha is associated with advanced bone marrow fibrosis in idiopathic myelofibrosis. Haematologica 90(1):133–134
Niino D, Tsuchiya T, Tomonaga M, Miyazaki Y, Ohshima K (2013) Clinicopathological features of acute megakaryoblastic leukaemia: relationship between fibrosis and platelet-derived growth factor. Pathol Int 63(3):141–149
Fava RA et al (1990) Synthesis of transforming growth factor-beta 1 by megakaryocytes and its localization to megakaryocyte and platelet alpha-granules. Blood 76(10):1946–1955
Janssens K, ten Dijke P, Janssens S, Van Hul W (2005) Transforming growth factor-beta1 to the bone. Endocr Rev 26(6):743–774
Yang M, Chesterman CN, Chong BH (1995) Recombinant PDGF enhances megakaryocytopoiesis in vitro. Br J Haematol 91(2):285–289
Su RJ et al (2001) Platelet-derived growth factor enhances ex vivo expansion of megakaryocytic progenitors from human cord blood. Bone Marrow Transplant 27(10):1075–1080
Tibbles HE, Navara CS, Hupke MA, Vassilev AO, Uckun FM (2002) Thrombopoietin induces p-selectin expression on platelets and subsequent platelet/leukocyte interactions. Biochem Biophys Res Commun 292(4):987–991
Malara A et al (2011) Megakaryocyte-matrix interaction within bone marrow: new roles for fibronectin and factor XIII-A. Blood 117(8):2476–2483
Abbonante V et al (2013) Discoidin domain receptor 1 protein is a novel modulator of megakaryocyte-collagen interactions. J Biol Chem 288(23):16738–16746
Chang Y et al (2007) Proplatelet formation is regulated by the Rho/ROCK pathway. Blood 109(10):4229–4236
Malara A et al (2011) Extracellular matrix structure and nano-mechanics determine megakaryocyte function. Blood 118(16):4449–4453
Shin JW, Swift J, Spinler KR, Discher DE (2011) Myosin-II inhibition and soft 2D matrix maximize multinucleation and cellular projections typical of platelet-producing megakaryocytes. Proc Natl Acad Sci USA 108(28):11458–11463
Richardson JL, Shivdasani RA, Boers C, Hartwig JH, Italiano JE (2005) Mechanisms of organelle transport and capture along proplatelets during platelet production. Blood 106(13):4066–4075
Pallotta I, Lovett M, Kaplan DL, Balduini A (2011) Three-dimensional system for the in vitro study of megakaryocytes and functional platelet production using silk-based vascular tubes. Tissue Eng Part C Methods 17(12):1223–1232
Omenetto FG, Kaplan DL (2010) New opportunities for an ancient material. Science 329(5991):528–531
Zhang J et al (2012) Stabilization of vaccines and antibiotics in silk and eliminating the cold chain. Proc Natl Acad Sci USA 109(30):11981–11986
Matsunaga T et al (2006) Ex vivo large-scale generation of human platelets from cord blood CD34 + cells. Stem Cells 24(12):2877–2887
de Barros AP et al (2010) Osteoblasts and bone marrow mesenchymal stromal cells control hematopoietic stem cell migration and proliferation in 3D in vitro model. PLoS One 5(2):e9093
Sullenbarger B, Bahng JH, Gruner R, Kotov N, Lasky LC (2009) Prolonged continuous in vitro human platelet production using three-dimensional scaffolds. Exp Hematol 37(1):101–110
Torisawa YS et al (2014) Bone marrow-on-a-chip replicates hematopoietic niche physiology in vitro. Nat Methods 11(6):663–669
Thon JN et al (2014) Platelet bioreactor-on-a-chip. Blood [Epub ahead of print]
Acknowledgments
This review was made possible through research support by Cariplo Foundation (2010-0807), Italian Ministry of Health (grant RF-2009-1550218), Italian Ministry of University and Research FIRB (RBFR1299KO) and a grant from Associazione Italiana per la Ricerca sul Cancro (AIRC, Milano) “Special Program Molecular Clinical Oncology 5 × 1000” to AGIMM (AIRC-Gruppo Italiano Malattie Mieloproliferative) and US National Institutes of Health (grant EB016041-01).
Conflict of interests
The authors declare no conflicts of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Malara, A., Abbonante, V., Di Buduo, C.A. et al. The secret life of a megakaryocyte: emerging roles in bone marrow homeostasis control. Cell. Mol. Life Sci. 72, 1517–1536 (2015). https://doi.org/10.1007/s00018-014-1813-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00018-014-1813-y