
Abstract
Thirty polymorphic simple sequence repeat (SSR) markers have been developed and characterized from RAPD product in Aspergillus niger in order to protect its natural resources. Polymorphisms of these RAPD-SSR markers were evaluated in a natural population of 25 strains collected from Shandong in China. The number of alleles (Na) per locus varied from 2 to 9. Observed (HO) and expected (HE) heterozygosities ranged from 0.56 to 0.96, and from 0.52 to 0.88, respectively. All the RAPD-SSR loci conformed to Hardy–Weinberg equilibrium after Bonferroni correction. They have showed sufficient level of polymorphisms to estimate the genetic diversity, population structure and species conservation in the A. niger. Five additional fungus species, Penicillium chrysogenum, Penicillium citrinum, Aspergillus flavus, Aspergillus ochratoxin and Aspergillus sulphureus were assessed for cross-species amplification. Three of the five species showed at least three polymorphic loci. In addition, eight loci were found to be polymorphic in at least one species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
INTRODUCTION
Aspergillus niger is a fungus and one of the most common species of the genus Aspergillus. Various strains of A. niger are cultured for the industrial production of many substances such as citric acid, gluconic acid and so on. Therefore, it is very important to collect and protect the natural resources of A. niger. In the long time, a good understanding of the genetic diversity, population structure and genetic differentiation of A. niger is required for the conservation and collection of this fungus. Microsatellites are tandemly repeated simple DNA sequences, which are widely dispersed throughout the genomes of eukaryotic and prokaryotic organisms (Eremenko et al., 2012; Zeng et al., 2013; Zhao et al., 2014; Xin et al., 2016; Zhdanova et al., 2016). Microsatellites are highly variable and most are thought to be selectively neutral, making them amendable to population genetic theory (Shaikhaev and Zhivotovsky, 2014). Microsatellites have become the marker of choice to study gene and genome evolution from the individual to populations or higher-level taxa. There are a lot of reports on molecular genetics of the genus Aspergillus performed using microsatellite markers in recent ten years (de Valk H.A. et al., 2007; Araujo et al., 2009; Christians and Watt, 2009; Araujo et al., 2010; Escribano et al., 2015; Kathuria et al., 2015). However, until now, only a small number of microsatellite sequences were available in GenBank for A. niger. Thus, screening for more polymorphic microsatellite markers in A. niger is very important for analyzing genome organization and evolution. In the present paper, we isolated 30 polymorphic RAPD-SSR markers derived from A. niger for the purpose of collecting and protecting its natural populations.
MATERIALS AND METHODS
Samples were collected from Dongying (DY), Yantai (YT) and Linyi (LY) in Shandong province, China in July, 2016. DNA extraction was performed as described by Sanchez et al. (2008) and Smirnova et al. (2011). The concentration was measured with a GENEQUANT Pro (Pharmacia Biotech Ltd., Cambridge, England) RNA/DNA spectrophotometer for absorption at 260 nm.
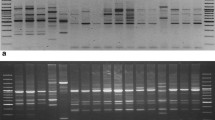
Randomly amplified polymorphism DNA (RAPD) method is a PCR-based DNA fingerprinting technique (Williams et al., 1990; Welsh et al., 1990). Genomic DNA was amplified with RAPD oligonucleotide primers. RAPD reactions were carried out in a 25-μL reaction mixture that included 20 pmol of RAPD primer, 100 μM of dNTPs, 10 mM Tris-HCl (pH 8.3), 50 mM KCl, 2.0 mM MgCl2, 1 unit of Taq polymerase (TaKaRa Corp.), and about 50 ng of template DNA. Amplification was performed on a Bioer Thermal Cycler (G-1000) (Bioer Corp.). PCR cycles were as follows: 5 min preamplification denaturation at 94°C, 45 cycles of 30 s at 94°C, 1 min annealing at 37°C, and 2 min extension at 72°C. As a final step, products were extended for 5 min at 72°C. Amplification products were then ligated into a T-vector that was used to transform into competent bacteria (Yokota and Oishi, 1990; Lunt et al., 1999). A total of 200 randomly selected clones were sequenced. Microsatellite sequences were screened using Tandem Repeats Finder (version: 2.02) (Benson, 1999). The criteria used in this software to identify microsatellites is as follows: 8 repeats for di-nucleotide repeat, 5 repeats for tri-nucleotide repeat, 4 repeats for tetra-nucleotide repeat.
Microsatellite amplification was performed using a standard procedure (Liu et al., 2012). PCR was performed in a 25-μL reaction mixture that included 6 pmol of each primer set, 100 μM of dNTPs, 10 mM Tris-HCl (pH 8.3), 50 mM KCl, 1.5 mM MgCl2, about 1.2 unit of Taq polymerase (TaKaRa Corp.), and approximately 120 ng of template DNA. PCR cycles were performed as Liu et al. (2014) reported. RAPD-SSR polymorphism was screened using ABI 377 automated DNA sequencer.
RESULTS
A total of 126 microsatellites were found, of which 79 pairs of primers were designed and tested. A total of 30 RAPD-SSR markers were found to be polymorphic among 25 strains of A. niger collected from Shandong in China (Table 1). The number of alleles per locus ranged from 2 (Aniger09 and Aniger16) to 9 (Aniger04 and Aniger18), and the observed and expected heterozygosity ranged from 0.56 to 0.96, and from 0.52 to 0.88, respectively (Table 1). We used ARLEQUIN 3.11 and POPGENE 1.3.1 to calculate observed (HO) and expected (HE) heterozygosity and linkage disequilibrium respectively. Significance values for all multiple tests were corrected following Sequential Bonferroni procedure (Rice, 1989). All the loci conformed to Hardy-Weinberg equilibrium (HWE) as determined by using the Markov–Chain method implemented. Out of 435 possible pairwise comparisons between the 30 loci applied to A. niger, none showed significant linkage disequilibrium. Considerable differences were found among DY, YT and LY populations in the number of alleles, effective number of alleles, number of genotypes at all of these loci (Table 2). According to total number of alleles, total effective number of alleles, and total number of genotypes, the LY population showed the highest diversity among all the samples while the YT sample was the lowest. These new microsatellite markers have provided a helpful tool for genetic analyses and resources conservation of A. niger. The method is also very useful in rapid screening polymorphic microsatellite markers based on RAPD products from fungus resources.
Cross-species amplification was examined on five other fungus species using the conditions detailed for A. niger. All primer pairs were tested on 25 strains from each of the five species, Penicillium chrysogenum, Penicillium citrinum, Aspergillus flavus, Aspergillus ochratoxin and Aspergillus sulphureus. Three of the five species (Aspergillus flavus, Aspergillus ochratoxin and Aspergillus sulphureus) showed at least three polymorphic loci. In addition, eight loci were found to be polymorphic in at least one species. The results, summarized in Table 3, highlight the possibility of some microsatellites of A. niger being used in studies on several other species of the fungus.
DISCUSSION
The RAPD technique is a PCR-based DNA fingerprinting that can rapidly identify hundreds of polymorphisms and has been successfully applied to a wide range of organisms (Williams et al., 1990; Welsh et al., 1990). The RAPD polymorphisms, as AFLP, usually include SNPs, insertions and deletions (indels) and microsatellites (Bradeen and Simon, 1998; Liu et al., 2015). Therefore, polymorphic RAPD bands contain many polymorphisms caused by microsatellite sequence. Traditionally, microsatellite loci have been isolated from small insert size of genomic libraries of the species of interest, screening thousands of clones through colony hybridization with repeat containing probes (Rassmann et al., 1991). However, its disadvantage is to need southern hybridization, which is time-consuming. If sequenced randomly without hybridizing for microsatellite enrichment, the microsatellite-containing sequence yield is low. A fast and effective protocol was used in this study for fast isolation of sequences containing microsatellite repeats from RAPD bands of A. niger. This method provides a very effective means to amplify large numbers of DNA suitable size for direct cloning and sequencing. It dose not require library construction and hybridization screening, but have a relative high microsatellite yield. We sequenced a total of 200 clones from RAPD bands of A. niger, and 126 microsatellite-containing sequences were obtained. That is, more than 60% of clones contain microsatellites. Among the 126 microsatellites, 79 pairs of primers were designed and tested, with 30 of these giving polymorphic loci. The other 47 microsatellites had inappropriate flanking regions on one or both sides of the simple sequence repeats or possessed only a few repeats and thus having less potential for polymorphism.
Microsatellite loci generally have ancient origins and show considerable evolutionary conservation, which suggests that microsatellite primers developed for any one locus may often be useful across a wide range of taxa (Liu et al., 2009). Cross-species amplification is a practical method to extend the utilization of microsatellite markers. Although cross-species amplification is convenient, it may lead to low or incomplete amplification due to as little as a single dinucleotide mismatch between the primer and the target DNA sequences. This problem can often be resolved by employing less stringent polymerase chain reaction (PCR) conditions such as lowering the annealing temperature or increasing the magnesium concentration in the reaction. In this study, 3, 6 and 3 microsatellite loci show polymorphic in Aspergillus flavus, Aspergillus ochratoxin and Aspergillus sulphureus, respectively. Nine microsatellite loci have amplification product and six are polymorphic in A. ochratoxin. It indicates A. niger may have a closer phylogenetic relationship with A. ochratoxin while farther with no polymorphic amplification of species of Penicillium chrysogenum and Penicillium citrinum.
In conclusion, the findings of this study prove the usefulness of RAPD bands as a valuable source for the identification of microsatellite from A. niger. The polymorphic microsatellite loci developed in this study for A. niger are anticipated for use in the further study of systematic and population genetics in this mold species. Cross-species amplification on five other species indicates that some microsatellite loci of A. niger are conservative in closely related species.
REFERENCES
Araujo, R., Amorim, A., and Gusmao, L., Genetic diversity of Aspergillus fumigates in indoor hospital environments, Med. Mycol., 2010, vol. 48, no. 6, pp. 832–838.
Araujo, R., Pina-Vaz, C., Rodrigues, A.G., Amorim, A., and Gusmao, L., Simple and highly discriminatory microsatellite-based multiplex PCR for Aspergillus fumigatus strain typing, Clin. Microbiol. Infect., 2009, vol. 15, no. 3, pp. 260–266.
Benson, G., Tandem repeats finder: A program to analyze DNA sequences, Nucleic Acids Res., 1999, vol. 27, no. 2, pp. 573–580.
Bradeen, J.M. and Simon, P.W., Conversion of an AFLP fragment linked to the carrot Y2 locus to a simple codominant PCR-based marker form, Theor. Appl. Genet., 1998, vol. 97, no. 5, pp. 960–967.
Christians, J.K. and Watt, C.A., Mononucleotide repeats represent an important source of polymorphic microsatellite markers in Aspergillus nidulans, Mol. Ecol. Resour., 2009, vol. 9, no. 2, pp. 572–578.
de Valk, H.A., Meis, J.F., and Klassen, C.H., Microsatellite based typing of Aspergillus fumigatus: Strengths, pitfalls and solutions, J. Microbiol. Methods, 2007, vol. 69, no. 2, pp. 268–272.
Eremenko, E.I., Ryazanova, A.G., Tsygankova, O.I., Tsygankova, E.A., Buravtseva, N.P., and Kuli-tchenko, A.N., Genotype diversity of Bacillus anthracis strains isolated from the Caucasus region, Mol. Genet., Microbiol. Virol., 2012, vol. 27, no. 2, pp. 74–78.
Escribano, P., Pelaez, T., Bouza, E., and Guinea, J., Microsatellite (STRAf) genotyping cannot differentiate between invasive and colonizing Aspergillus fumigatus isolates, J. Clin. Microbiol., 2015, vol. 53, no. 2, pp. 667–670.
Kathuria, S., Sharma, C., Singh, P.K., Agarwal, P., Agarwal, K., Hagen, F., Meis, J.F., and Chowdhary, A., Molecular epidemiology and in-vitro antifungal susceptibility of Aspergillus terreus species complex isolates in Delhi, India: Evidence of genetic diversity by amplified fragment length polymorphism and microsatellite typing, PLoS One, 2015, vol. 10, no. 3, p. e0118997.
Liu, J., Qu, J., Yang, C., Tang, D., Li, J., Lan, H., and Rong, T., Development of genome-wide insertion and deletion markers for maize, based on next-generation sequencing data, BMC Genomics, 2015, vol. 16, p. 601.
Liu, Y.G., Guo, Y.G., Hao, J., and Liu, L.X., Genetic diversity of swimming crab (Portunus trituberculatus) populations from Shandong peninsula as assessed by microsatellite markers, Biochem. Syst. Ecol., 2012, vol. 41, no. 1, pp. 91–97.
Liu, Y.G., Liu, L.X., and Xing, S.C., Development and characterization of thirteen polymorphic microsatellite loci in the Cynoglossus semilaevis, Conserv. Genet. Resour., 2014, vol. 6, no. 3, pp. 683–684.
Lunt, D.H., Hutchinson, W.F., and Carvalho, G.R., An efficient method for PCR-based isolation of microsatellite arrays (PIMA), Mol. Ecol., 1999, vol. 8, no. 5, pp. 891–893.
Rassmann, K., Schlotterer, C., and Tautz, D., Isolation of simple sequence loci for use in polymerase chain reaction-based DNA fingerprinting, Electrophores, 1991, vol. 12, pp. 113–118.
Rice, W.R., Analyzing tables of statistical tests, Evolution, 1989, vol. 43, no. 1, pp. 223–225.
Sanchez, B., Rodriguez, M., Casado, E.M., Martin, A., and Cordoba, J.J., Development of an efficient fungal DNA extraction method to be used in random amplified polymorphic DNA-PCR analysis to differentiate cyclopiazonic acid mold producers, J. Food Prot., 2008, vol. 71, no. 12, pp. 2497–2503.
Schneider, S., Roessli, D., and Excoffier, L., ARLEQUIN: A Software for Population Genetics Data Analysis, Geneva: Genetics and Biometry Laboratory, Univ. of Geneva, 2000.
Shaikhaev, E.G. and Zhivotovsky, L.A., The evolution of microsatellite loci in salmonid fishes, Russ. J. Genet., 2014, vol. 50, no. 8, pp. 853–859.
Smirnova, N.I., Zadnova, S.P., Shashkova, A.V., and Kutyrev, V.V., Genome variability in the altered variants of Vibrio cholerae biovar El Tor isolated in Russia, Mol. Genet., Microbiol. Virol., 2011, vol. 26, no. 3, pp. 102–110.
Welsh, J. and McClelland, M., Fingerprinting genomes using PCR with arbitrary primers, Nucleic Acids Res., 1990, vol. 18, no. 24, pp. 7213–7218.
Williams, J.G.K., Kubelik, A.R., Livak, K.J., et al., DNA polymorphisms amplified by arbitrary primers are useful as genetic markers, Nucleic Acids Res., 1990, vol. 18, pp. 6531–6535.
Xin, M.M., Zhang, S.H., Wang, D.Q., Li, C.J., Yue, H.M., and Wei, Q.W., Twenty-four novel microsatellites for the endangered Chinese sturgeon (Acipenser sinensis Gray, 1835), J. Appl. Ichthyol., 2016, vol. 32, no. 3, pp. 405–408.
Yeh, F.C., Yang, R.C., and Boyle, T., POPGENE version 1.3.1. Microsoft Window-Bases Freeware for Population Genetic Analysis, Univ. of Alberta and the Centre for Int. Forestry Research, 1990. http://www.ualberta.ca/~fyeh/.
Yokota, H. and Oishi, M., Differential cloning of genomic DNA: Cloning of DNA with an altered primary structure by in-gel competitive reassociation, Proc. Natl. Acad. Sci. U. S. A., 1990, vol. 87, no. 16, pp. 6398–6402.
Zeng, Q., Ye, H., Ludwig, A., Wang, Z., Zhang, Y., and Peng, Z., Microsatellite development for the endangered Yangtze sturgeon (Acipenser dabryanus Dumeril, 1869) using 454 sequencing, J. Appl. Ichthyol., 2013, vol. 29, no. 6, pp. 1219–1221.
Zhao, J., Zhao, K., and Peng, Z., Development and characterization of eleven microsatellite markers for an endangered cavefish (Triplophysa rosa Chen and Yang, 2005) using 454 sequencing, J. Appl. Ichthyol., 2014, vol. 30, no. 5, pp. 1056–1058.
Zhdanova, S.N., Ogarkov, O.B., Alexeeva, G.I., Vinokurova, M.K., Sinkov, V.V., Astaf’ev, V.A., Savilov, E.D. and Kravchenko, A.F., Genetic diversity of Mycobacterium tuberculosis isolates in the Republic of Sakha (Yakutia), Russia, Mol. Genet., Microbiol. Virol., 2016, vol. 31, no. 2, pp. 51–57.
ACKNOWLEDGMENTS
This work was supported by grants from the High-level talent introduction project of Xinjiang Uygur Autonomous Region (201412) and the project of domestic visiting scholars for young College Backbone Teachers in Shandong province (SD2014).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
The authors declare that they have no conflict of interest. This article does not contain any studies involving animals or human participants performed by any of the authors.
Additional information
The article is published in the original.
About this article

Cite this article
Yun-Guo Liu, Han, QD., Li, YY. et al. Isolation and Characterization of Thirty Polymorphic Microsatellite Markers from RAPD Product in Aspergillus niger and a Test of Cross-Species Amplification. Mol. Genet. Microbiol. Virol. 33, 254–260 (2018). https://doi.org/10.3103/S0891416818040043
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3103/S0891416818040043