
Abstract
Zooplankton community and its relationship to environmental variables were investigated seasonally between autumn 2006 and summer 2008 in the Gulf of Bandırma (the Sea of Marmara). In the Gulf, we identified nine Copepoda, three Cladocera, one Hydrozoa, one Cydippida, Appendicularia, Bivalvia, Chaetognatha, Cirripedia, Decapoda, Echinodermata, Gastropoda, Polychaeta, Pisces larvae, and Pisces eggs at three sampling stations. Shannon-Weaver diversity index and Pielou’s evenness value ranged between 0.00–2.72 and 0.00–1.00, respectively. The first two components of the redundancy analysis (RDA) explained 81.9% of the variance as the main factors (generally the longest arrows) affecting the spatial and temporal distributions of zooplankton in the Gulf of Bandırma, with dissolved oxygen (DO), nitrite+nitrate (NO3 + NO2-N) and phosphate (PO4-P). Liriope tetraphylla, Oithona nana, Paracalanus parvus, Penilia avirostris, Appendicularia and veliger larvae (Gastropoda) were positively related to DO and salinity, and negatively correlated with temperature, chlorophyll-a (Chl-a), Trophic State Index (Trix), PO4-P, and NO3 + NO2-N. According to Spearman’s rank correlation analysis, there was a strong positive correlation between the number of zooplankton individuals and temperature, and the number of zooplankton individuals negatively correlated with salinity and Chl-a. In order to determine similarities among seasons in the samples, the Bray-Curtis similarity index was calculated using species diversity and abundance values, while Euclidean distances were calculated using ecological parameters. Overall, the diversity of zooplankton in the Gulf of Bandırma was higher in the first sampling year and decreased due to the effect of mucilage in the second sampling year.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In marine ecosystems, zooplankton plays an important role as a primary consumer and a secondary producer. Zooplankton plays a key role in the pelagic food web by transferring photosynthetically produced energy to higher trophic levels. Due to its importance in the food chain, more zooplankton in an aquatic environment can lead to more fish populations feeding on zooplankton. Changes in environmental conditions affect the population structure and distribution of zooplankton, as well as other trophic levels (Wang et al. 2014).
With a surface area of 11,500 km2 and a maximum depth of 1390 m, the Sea of Marmara is a small basin between the continents of Asia and Europe, forming the “System of Turkish Straits” between Bosphorus and the Dardanelles (Beşiktepe et al. 1994). While brackish waters from the Black Sea, with a salinity of 17.6, flow into the Sea of Marmara with the overcurrent through the Bosphorus, waters from the Aegean Sea (the salinity is app. 38.5) are carried into the Sea of Marmara with the undercurrent through the Çanakkale. Between these two strata, which do not mix with each other, there is an intermediate layer of salinity approximately 25 m deep (Ullyott and Pektaş 1952). These deep salt waters have a low content of oxygen. Fauna and flora of the Sea of Marmara, which contains both brackish and typical sea water, are diverse.
Bandırma is a heavily populated area (> 120.000 people) impacted by anthropogenic and industrial pollution. According to Balkis et al. (2012), the phosphate content of the surface waters of Bandırma, located at the northeast Gulf of Bandırma and Kapıdağ Peninsula, are the highest in the Sea of Marmara. Domestic waste and the chemical fertilizer factory in this region are the primary causes of the increase in phosphate in the gulf (Balci et al. 2014). In addition, the nutrient contents of surface and bottom waters of Bandırma are quite rich. There are fertilizer and acid industry factories in the coastal areas of this gulf (Balci et al. 2014). Susurluk Stream, which originates from a combination of the Kara Stream and Manyas Lake (Including the Bird Paradise National Park, known for its host to many birds), is the most important provider of fresh water and nutrients in the gulf. There is also an important port in the Gulf of Bandırma. The economic importance of Bandırma city has increased in the Marmara Region due to intensive white meat and fertilizer production; however, this negatively affects the Gulf of Bandırma (Balci et al. 2014).
Many studies have been conducted on the zooplankton in the Sea of Marmara (Öğdül and Ergüven 1992; Benli et al. 2001; Tarkan et al. 2005; Svetlichny et al. 2006; Isinibilir et al. 2008; İşinibilir-Okyar et al. 2015). By contrast, in the Gulf of Bandırma, which is located to the south of the Sea of Marmara, there are few studies that are not comprehensive (Isinibilir 2010; Isinibilir et al. 2010).
In this study, the seasonal variations in zooplankton distribution, abundance and community composition in the Gulf of Bandırma (the Sea of Marmara) and the effects of several environmental factors on their distributions were analysed. In addition, this study is important as there are not enough comprehensive studies on zooplankton in the Gulf of Bandırma, fishing activities are carried out in this region, and the gulf is threatened with intense pollution. These data on zooplankton will contribute to the biodiversity in the Sea of Marmara.
Materials and methods
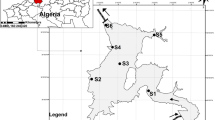
Zooplankton samples were collected seasonally at three stations (30 m depth to surface, vertically and horizontally) from the autumn (November) 2006 to the summer (August) 2008 in the Gulf of Bandırma using a WP2 plankton net with a 57 cm mouth diameter and 200 μm mesh size between 08:00 and 12:00 a.m. (Fig. 1). All zooplankton samples were preserved in 4% buffered formaline solution. Both horizontal and vertical sampling were used in the qualitative evaluation of the data (Table 1), while only vertical sampling was considered in the quantitative evaluation. Statistically, only quantitative data were included in the analyses.

The locations of the three stations in the Gulf of Bandırma
Samples were analysed into groups or species using a Bogorov counting chamber under a stereomicroscope (10-100X magnification). Two subsamples from each sample were taken from a storage bottle with 200 ml volume using a pipette (2 ml) for the quantitative analysis of zooplankton number of specimens. References used to identify the zooplankton can be found in previous work (Toklu Alıçlı and Sarihan 2016). Systematic classifications and nomenclatures of zooplankton species were made according to World Register of Marine Species (WoRMS 2020). In addition, temperature, salinity, dissolved oxygen (DO), Chl-a, nitrate+nitrite-N, phosphate-P and silicate-Si were obtained from the work of Balkis et al. (2012) which was carried out synchronously with this study. The average values of each ecological variable for each station were used in this study.
The biodiversity of the zooplankton community was analysed using the Pielou’s evenness index (Pielou 1966) for the equitability of species abundance, Shannon-Weaver diversity index (Shannon and Weaver 1964), and Simpson’s dominance index (Simpson 1949). The relationship between zooplankton diversity and the environmental variables were tested using detrended correspondence analysis (DCA) using CANOCO v4.0 for Windows (Ter Braak and Smilauer 1998). Before the analysis Kolmogorov-Smirnov test was applied to determine the normal distribution of the environmental data (Zar 2010). In the DCA analyses, the length of the gradients was 2.59 and 2.00 for the first and second axes, which were shorter than four, indicating that zooplankton diversity showed linear variation (Leps and Smilauer 2003) were suitable for the RDA. The significance of the relationships between the environmental variables and species data (rare species less than three occurrences were excluded) were tested using a Monte Carlo permutation test (p < 0.05) with automatic forward selection for the redundancy analysis (RDA) (Ter Braak 1986). Spearman’s rank correlation coefficients (Siegel 1956) were used to detect any correlations among the ecological variables and zooplankton abundance. Similarities between sampling stations were measured by Bray-Curtis similarity indices [log (x + 1)] and Euclidean distances (normalized) in Primer v6 software (Clarke and Warwick 2001).
Results
In the Gulf of Bandırma, 24 zooplankton taxa were identified between 2006 and 2008 at the three sampling stations (Table 1). During this study, the highest number of zooplankton taxa was found in the autumn of 2007 (13 taxa) and in summer 2008, the lowest number was found in winter 2008 (one taxon). The highest numbers of taxa were obtained from station 2 (St-2) in the spring and the autumn of 2007 while no taxa were found in St-2 and station 3 (St-3) in winter 2008.
When the abundance data were evaluated (Fig. 2), the highest number of individuals was obtained in spring 2008 (St-2, 1717 ind m−3) and no individuals was obtained in winter 2008 (St-2, St-3). The dominance of the copepod Acartia clausi is remarkable in the number of individuals in spring 2008 (1660 ind m−3). In winter 2008 no individuals were found in St-2 and 3, while only 4 ind m−3 were observed in station 1 (St-1). Acartia clausi was observed in all seasons except for autumn 2006 and summer 2007. No zooplankton taxa other than A. clausi were observed in winter 2008. When the number of individuals was evaluated, we found that Hydrozoa (Liriope tetraphylla, ~ 84% of total individuals) in summer 2007, Copepoda (A. clausi, ~ 95% of total individuals) in spring 2008, and Cladocera (Penilia avirostris, ~ 81% of total individuals) in summer 2008 significantly increased in all stations (Fig. 2). In addition, some species (Evadne spinifera, Penilia avirostris, Pleopis polyphmoides, Centropages typicus, Corycaeus spp. and Oncaea spp.) were just observed in one season, while some taxa (Corycaeus spp. in St-1 and Oncaea spp. in St-2) were observed in only one station (Table 1). Among these species, P. avirostris and P. polyphmoides which were observed in only one season and found in all stations, reached remarkable values in terms of abundance (314–732 ind m−3 in spring 2008, 35–83 ind m−3 in autumn 2007; respectively).

Seasonal variations in the abundance of zooplankton taxa at three stations in the Gulf of Bandırma
The Shannon-Weaver diversity index (H′) ranged from 0.00 (winter 2008) to 2.72 (spring 2007), the Pielou’s evenness index (J’) from 0.00 (winter 2008) to 1.00 (winter 2007) and the Simpson dominance index (λ) from 0.18 (spring 2007) to 1.00 (winter 2008) (Fig. 3). The highest dominance (λ) and the lowest diversity indices (H′ and J’) were obtained at a sampling station in winter 2008 due to the only recorded one taxon (Acartia clausi, 4 ind m−3 in St-1). Nine zooplankton taxa found in St-2 in spring 2007 showed a balanced distribution in terms of the number of individuals, and the H′ increased in this season compared with other periods and stations. In addition, the dominance index value was the lowest in this period because the dominance of Pleurobrachia pileus was below 30%. In the winter of 2007, the index was the highest (1.00) in St-3 because equal numbers of individuals were found for Paracalanus parvus, Pleurobrachia pileus and Bivalvia larvae.

Seasonal zooplankton species diversity (H′), dominance (λ) and evenness (J′) indices in the stations in Gulf of Bandırma
In the Gulf of Bandırma, water temperature ranged between 8.07 and 22.00 °C, salinity between 26.00 and 31.03, DO between 5.15 and 13.41 mg L−1, Chl-a values between 0.52 and 9.21 μg L−1. The amount of nitrate+nitrite-N (NO3 + NO2-N) ranged between 0.55 and 3.12 μg-at N L−1, phosphate-P (PO4-P) concentrations between 0.43 and 5.47 μg-at P L−1, amount of silicate-Si between 1.04 and 7.53 μg-at Si L−1 (Table 2; Balkis et al. 2012). Spearman’s rank-correlation analysis (rs) was performed with biotic and abiotic environmental factors (Table 3). Accordingly, the abundance of zooplankton was positively correlated with temperature (p < 0.01) and negatively correlated with salinity and Chl-a (p < 0.05). Even though we found a positive relationship (p < 0.05) between the number of taxa and abundance, this trend was not valid for the overall study. Moreover, a positive relationship (p < 0.05) was detected between J’ and DO and SiO4-Si.
In the DCA analyses, the length of the gradients was 2.59 and 2.00 for the first and second axes, which were shorter than 4, indicating that zooplankton diversity showed linear variation and was suitable for the RDA (Leps and Smilauer 2003). In the RDA, the eigenvalues of the first two axes were λ1 = 0.158 and λ2 = 0.114 and accounted for 50.5% of the species-environment relation, indicating a strong taxon-environment correlation (91.9% and 81.9%). In the Monte Carlo permutation test (p < 0.05) with automatic forward selection for the redundancy analysis (RDA) SiO4-Si was omitted with respect to its minor effect on zooplankton diversity. The arrows of DO, NO3 + NO2-N and PO4-P were the longest on the RDA diagram which might be evaluated as the most important variable regarding zooplankton diversity. On the RDA diagram (Fig. 4), axis 1 was positively correlated with Trix, Chl-a, temperature, NO3 + NO2-N and PO4-P, and was negatively correlated with DO, Secchi disk depth and salinity. Two axes showed a positive correlation with only Chl-a, temperature and PO4-P.

RDA showing the distribution of stations, zooplankton assemblages in relation to the selected environmental variables in the Gulf of Bandırma
The RDA axes scores distinguished two zooplankton assemblages generated by axis 2, including Liriope tetraphylla, Oithona nana, Paracalanus parvus, Appendicularia and Trochophora/Veliger larvae (Gastropoda and Bivalvia), and additional groups with other species. The first group that was represented by six taxa and related to the three sampling stations positively correlated with DO, Secchi disk depth and salinity. The second zooplankton group of 13 taxa was positively associated with Chl-a, temperature, NO3 + NO2-N, PO4-P, Trix and Chl-a. Additionally, the ordination analysis revealed a significant inverse relationship of PO4-P, Trix, temperature and Chl-a with DO, and salinity. According to the RDA analysis, there was a strong negative association between O. nana and salinity. Liriope tetraphylla was found to be positively related to DO and Secchi disk depth. Temperature, Chl-a, Trix and PO4-P positively correlated with Penilia avirostris while showed negative correlation with O. nana, P. parvus and Appendicularia. The close relationship between summer 2008 sampling season and temperature, Chl-a, Trix and PO4-P is clearly seen in the RDA diagram. The results of the Bray-Curtis similarity index, which was applied to determine the similarity in the number of zooplankton taxa and their abundances among seasons and stations, are given in Fig. 5a and b. When the results were evaluated seasonally, the similarity of the sampling periods was 11.8%, with the exception of the winter of 2008.

Bray-Curtis similarity (a, b) and Euclidean distance (c, d) dendogram of the seasons and stations in the Gulf of Bandırma
The number of individuals in winter 2008 was 4 ind m−3, which differed from all other seasons. When the seasons with 5% similarity were evaluated, three different groups and two different seasons were observed. In two of the groups, consecutive seasons formed different groups. The similarity between autumn 2006 and summer 2007 was 67%, the highest similarity recorded. In terms of stations, St-1 and St-2 were 87.2% similar, whereas the similarity across all stations was 80.2%. Similarly, when the RDA analysis was taken into consideration, stations had similar distributions, and there was no significant difference between the stations (Fig. 4).
The dendrogram of the Euclidean distances (ED) used to determine the similarity of ecological variables is given in Fig. 5c and d. According to this dendrogram, the similarity of all seasons (except for winter 2008) was 7.4 ED. While phosphate values were higher in summer 2008 than in the other seasons, silicate and DO values were lower. Because of this, the summer of 2008 was separated from the other seasons. When the seasons were evaluated with similarity at 4 ED, two groups and four different seasons were observed. The similarity between winter 2007 and autumn 2007 was observed at 2.5 distance. The highest similarity was recorded between these seasons. When the ED was evaluated in terms of stations, St-2 and St-3 were similar in 13.2 ED. St-1 was found to be similar to other stations at a distance of 14.1. According to these results, there is no clear similarity between the Bray-Curtis similarity and ED dendrograms (by season or station).
Discussion
The relationship between environmental factors and zooplankton diversity in marine ecosystems due to their rapid response to the changes allows us to understand the natural processes that occur in marine ecosystems caused by such as climate change, pollution (Chiba et al. 2018). In this study, which deals with the diversity of zooplankton and their relationship with ecological variables in the Gulf of Bandırma located in the east of the Kapıdağ Peninsula, a total of 24 zooplankton taxa were determined. In a previous study conducted by Isinibilir (2010) in the same gulf, 39 zooplankton species were reported, but a species list was not presented. In another study (Toklu-Alıçlı et al. 2014) carried out in the Gulf of Erdek, which is located in the west of the peninsula, 26 taxa were reported of which 17 were identified on the basis of genus or species and nine on the basis of group. While all taxa recorded in the present study were also reported from the Gulf of Erdek, Pseudocalanus elongates (Brady, 1865), Anomalocera patersonii Templeton, 1837, and Euterpina acutifrons (Dana, 1847) were not found in the Gulf of Bandırma. In a review article (Yilmaz and Isinibilir 2016), 129 species of Copepoda have been reported in the Sea of Marmara from the 1950s to nowadays, nine of which were detected in this study. The reason why fewer species were found in our study is that the previous study is a checklist of Copepoda in the Sea of Marmara. Additionally, Copepoda represented the majority of the species (9 species) in the Gulf of Bandırma. Unal et al. (2000) found 111 Copepoda taxa, Benli et al. (2001) found 30 Copepoda taxa and Toklu-Alıçlı et al. (2014) found 12 Copepoda taxa in the Sea of Marmara. In another study (Toklu-Alıçlı et al. 2014) conducted in the south of the Sea of Marmara, the highest number of taxa (21 taxa) was found in the spring of 2008 in the Gulf of Erdek. During the winter of 2008, in which the mucilage event in the Sea of Marmara was observed (Balkis et al. 2011; Tüfekci et al. 2010), a single species (Acartia clausi) was found only in St-1. There were six Cladocera (Yilmaz and Isinibilir 2016), 62 Hydrozoa and four Ctenophora (Isinibilir and Yilmaz 2016) taxa reported for the Sea of Marmara. In this study, the Cladocera were represented by three species, while Hydrozoa and Ctenophora were represented by one species each.
The Marmara Sea is a very small basin with a connection to the Mediterranean and Black Sea via the Turkish Straits System (TSS). In a review article taking into account the Copepoda diversity in the Mediterranean (Aker 2015), the presence of 285 taxa in the Adriatic Sea, 24 taxa in the Ionian and Sicilian-Libyan Region, 319 taxa in the Levantine Sea and 202 taxa in the Aegean Sea were recorded. The pelagic ecosystem of the Marmara has some basic common characteristics with the Northern Aegean Sea because of the connections through the TSS (Altuğ et al. 2011). Acartia clausi has been reported as an indicator of pollution in the Bay of Biscay and the Aegean Sea (Uriarte and Villate 2005; Sever 2009). Also, Sever (2009) was found that A. clausi was dominant in the spring and winter seasons in the Aegean Sea. In a study by Najdek (1997) conducted during mucilage formation in the Adriatic Sea, A. clausi was found to be the primary dominant Copepoda species. Acartia clausi was abundantly observed in the region where the study was conducted by Isinibilir (2010). Likewise, İşinibilir-Okyar et al. (2015) acquired similar results in the northeastern Sea of Marmara during the mucilage event in 2008, showing that mucilage formation had a significant effect on zooplankton abundance and community structure. Similarly, in present study, A. clausi was the dominant species (1660 ind m−3) in the spring of 2008, following the mucilage event in the Sea of Marmara. Acartia clausi was observed in all seasons except for in the autumn of 2006 and summer of 2007 in the present study. Similarly, no zooplankton group or species were encountered except for two copepod species and a meroplankton taxon in the Gulf of Erdek (Toklu-Alıçlı et al. 2014). These findings may be related to the mucilage event, which occurred in the Sea of Marmara between October 2007 and June 2008, causing environmental problems which impacted fishery activities as result of excess nutrient loading into coastal areas.
Paracalanus parvus, which is defined as one of the main components of coastal zooplankton for the Mediterranean and Black Sea by Kasapidis et al. 2018, has also been reported in the Sea of Marmara. This species was found by Tarkan et al. (2005) in the spring of 2005 and Svetlichny et al. (2006) in October 2005. In this study, P. parvus, was observed in every season except for winter 2008. Additionally, Corycaeus spp. in autumn of 2007, Centropages typicus in spring 2008, and Oncaea spp. in autumn 2006 were recorded. Cladocera species Pleopis polyphemoides (autumn 2007), Evadne spinifera (spring 2008), and Penilia avirostris (summer 2008) were detected in the gulf. Appendicularia were recorded in all seasons throughout the study except for in the winter of 2008, while Cirripedia larvae were recorded only in the summer of 2008. Benli et al. (2001) determined that Centropages typicus were abundant (22%) in winter, and Öğdül and Ergüven (1992) and Tarkan et al. (2005) never encountered this species in their study in the Istanbul Strait. In this study, C. typicus only consisted of 0.09% of the zooplankton in spring 2008. In addition, Liriope tetraphylla, which was reported as a new record for the Sea of Marmara in 2005 (Isinibilir et al. 2010), was found in six seasons (autumn 2006, winter 2007, spring 2007, summer 2007, autumn 2007 and summer 2008) in this study.
According to the number of individuals in the stations, individuals belonging to Cladocera reached the highest levels during summer 2008 and the maximum number of individuals were found in St-2. Penilia avirostris (732 ind m−3) was observed to cause this increase. In the first sampling year of this study, P. avirostris has never been seen (the mucilage may be a factor). However, in the second sampling year, this species was found only in summer 2008 and it was dominant (76.5–87.8% in the stations). This situation may be due to the abundance of organisms that can be fed for grazing after the mucilage event. Also, it is known as a key component of mesozooplankton in coastal waters during stratification and important in the marine ecosystem because of its high filtering capacity and high reproductive rate and make a significant contribution to the energy and matter cycles (Atienza et al. 2006). When evaluating the studies about P. avirostris in the Sea of Marmara, Isinibilir (2010) stated that the species was dominant in summer in the Gulf of Bandırma. Atienza et al. (2008) found that P. avirostris was abundant (2500–3000 ind m−3) in summer in the Catalan Sea, whereas İşinibilir-Okyar et al. (2015) encountered P. avirostris from July to September, stating that the species is also dominant in September.
The study conducted in the northeastern area of the Sea of Marmara in January 2004 and December 2007 stateed that L. tetraphylla bloomed in 2006 and 2007, and therefore, zooplankton abundances were negatively impacted (Yilmaz 2015). In a study conducted in the Gulf of Bandırma, Isinibilir et al. (2010) encountered L. tetraphylla in October 2006 and March 2007. Yilmaz (2015) stated that L. tetraphylla was a very abundant taxon in 2006 and 2007 in the Turkish Straits System. The dominant summer–autumn species P. avirostris disappeared in autumn 2006 and diminished by approximately 30-fold in 2007 when compared to years without L. tetraphylla. These results indicate how the existence of L. tetraphylla negatively affects zooplankton structure. In this study, L. tetraphylla was observed during the same years. While the species was observed in every station in autumn 2006 and summer 2007, it constituted 84.4% of the zooplankton in summer 2007.
Statistically, the number of zooplankton individuals was positively correlated to temperature (p < 0.01) and negatively correlated to salinity and Chl-a (p < 0.05) in the Gulf of Bandırma. There are studies indicating that the factor affecting seasonal changes in zooplankton populations is temperature (Siokou-Frangou et al. 1998; Villate 1994). According to the RDA, DO, NO3 + NO2-N and PO4-P related to temperature showed the longest arrow on the diagram which might be evaluated as the main factor affecting the spatial and temporal distributions of zooplankton in the Gulf of Bandırma. This relationship network is similar to that which was found in the CCA analysis of Toklu-Alıçlı et al. (2014). However, Toklu-Alıçlı et al. (2014) reported that the number of zooplankton individuals was positively correlated with salinity in the Gulf of Erdek. In addition, they found a negative relationship between the number of individuals and Chl-a, as in this study. This relationship is explained by the fact that zooplankton graze on small phytoplankton (Terbiyik Kurt and Polat 2015). In contrast, no relationship was found between nutrients and the number of zooplankton species and their abundances. According to the RDA analysis, there was a strong negative association of Chl-a with Oithona nana and Paracalanus parvus. Such a relationship may be due to O. nana grazing on small phytoplankton (heavily on diatoms) (Hafferssas and Seridji 2010). Liriope tetraphylla was found to be positively related to DO and Secchi disk depth.
Although the number of species increased in many seasons, the number of individuals was low, and in some seasons, even if the number of species was low, the dominant species increased to high abundances. It must be considered that during the mucilage event in winter 2008, the number of taxa and individuals was very low, possibly leading to an incorrect evaluation. Moreover, due to the resting period and cyst formation in coastal zooplankton, differences between stations, seasons and years can be encountered (Belmonte and Rubino 2019). Therefore, as emphasized in conservation biology studies, predominant species (often referred to as “umbrella species”), could help protect other species living in the same ecosystem so that they can exist in the environment with other species and increase the number of species in the ecosystem (Weng et al. 2015). The positive relationship (p < 0.05) between J’ and DO can be explained by the high J’ values in spring and autumn and the increase in DO during these periods. There was no significant relationship detected between silicate and the number of zooplankton species or individuals, but there was a positive relationship between silicate and J’.
Conclusions
In conclusion, this is a comprehensive study on zooplankton in the Gulf of Bandırma. Zooplankton species and abundances were evaluated together with environmental variables, and it was determined that the factor of temperature had an effect on zooplankton. Also, chlorophyll-a and salinity influenced the spatial and temporal distributions of zooplankton in the Gulf of Bandırma. In addition, Acartia clausi was the dominant species in the spring 2008, while Penilia avirostris was dominant in the summer 2008. It was observed that the diversity was higher in the first sampling year of the study and decreased due to the effect of mucilage in the second sampling year. The seasonal succession and distribution of zooplankton in the marine environment depends on the biotic and abiotic dynamics resulting in temporal and spatial changes in the plankton. Long-term monitoring and frequent sampling is needed to better understand the response of plankton to these changes. As a result of the study, it can be said that the diversity was higher in the first sampling year, and it decreased due to the effect of mucilage in the second sampling year in the Gulf of Bandırma.
Data availability
Not applicable.
References
Aker HV (2015) Zooplankton of the Aegean Sea. In: Katağan T, Tokaç A, Beşiktepe Ş, Öztürk B (eds) The Aegean Sea marine biodiversity, fisheries, conservation and governance. Turkish Marine Research Foundation (TUDAV), Istanbul, pp 176–187
Altuğ G, Aktan Y, Oral M, Topaloğlu B, Dede A, Keskin Ç, İşinibilir M, Çardak M, Çiftçi PS (2011) Biodiversity of the northern Aegean Sea and southern part of the Sea of Marmara, Turkey. Mar Biodivers Rec 4:e65. https://doi.org/10.1017/S1755267211000662
Atienza D, Saiz E, Calbet A (2006) Feeding ecology of the marine cladoceran Penilia avirostris: natural diet, prey selectivity and daily ration. Mar Ecol Prog Ser 315:211–220. https://doi.org/10.3354/meps315211
Atienza D, Saiz E, Skovgaard A, Trepat I, Calbet A (2008) Life history and population dynamics of the marine cladoceran Penilia avirostris (Branchiopoda: Cladocera) in the Catalan Sea (NW Mediterranean). J Plankton Res 30(4):345–357. https://doi.org/10.1093/plankt/fbm109
Balci M, Balkis N, Durmus T, Sivri N (2014) Seasonal variations of nutrients and chlorophyll-a in the coastal waters of the Kapidag peninsula (Marmara Sea). Fresenius Environ Bull 23(12b):3391–3399
Balkis N, Atabay H, Türetgen I, Albayrak S, Balkis H, Tüfekçi V (2011) Role of single-celled organisms in mucilage formation on the shores of Büyükada Island (the Marmara Sea). J Mar Biol Assoc UK 91:771–781. https://doi.org/10.1017/S0025315410000081
Balkis N, Toklu-Alicli B, Balci M (2012) Evaluation of ecological quality status with the trophic index (TRIX) values in the coastal waters of the gulfs of Erdek and Bandırma in the Marmara Sea. In: Voudouris K (ed) Ecological water quality - water treatment and reuse. InTech, Rijeka, Croatia, pp 1–22. https://doi.org/10.5772/33698
Belmonte G, Rubino F (2019) Resting cysts from coastal marine plankton. In: Hawkins SJ, Allcock AL, Bates AE, Firth LB, Smith IP, Swearer SE, Todd PA (eds) Oceanography and marine biology: an annual review (57). CRC Press, Florida, pp 1–88
Benli H, Tarkan A, Sever T (2001) Comparison of the mesozooplankton composition the southwestern Black Sea, sea of Marmara and eastern Aegean Sea. Turkish J Mar Sci 7(3):163–179
Beşiktepe ŞT, Sur HI, Özsoy E, Abdul Latif M, Oǧuz T, Ünlüata Ü (1994) The circulation and hydrography of the Marmara Sea. Prog Oceanogr 34:285–334. https://doi.org/10.1016/0079-6611(94)90018-3
Chiba S, Batten S, Martin CS, Ivory S, Miloslavich P, Weatherdon LV (2018) Zooplankton monitoring to contribute towards addressing global biodiversity conservation challenges. J Plankton Res 40(5):509–518. https://doi.org/10.1093/plankt/fby030
Clarke KR, Warwick RM (2001) Change in marine communities: an approach to statistical analysis and interpretation. PRIMER-E, Plymouth-UK
Hafferssas A, Seridji R (2010) Relationships between the hydrodynamics and changes in copepod structure on the Algerian coast. Zool Stud 49(3):353–366
Isinibilir M (2010) Spatial and temporal variation of zooplankton in two bays in the southern sea of Marmara. Crustaceana 83(2):233–244. https://doi.org/10.1163/001121609X12512848343568
Isinibilir M, Yilmaz IN (2016) Jellyfish species of the sea of Marmara. In: Özsoy E, Çağatay MN, Balkıs N, Balkıs N, Öztürk B (eds) The sea of Marmara marine biodiversity, fisheries, conservation and governance. Turkish Marine Research Foundation (TUDAV), Istanbul, pp 390–400
Isinibilir M, Kideys AE, Tarkan AN, Yilmaz IN (2008) Annual cycle of zooplankton abundance and species composition in Izmit Bay (the northeastern Marmara Sea). Estuar Coast Shelf S 78(4):739–747. https://doi.org/10.1016/j.ecss.2008.02.013
Isinibilir M, Yilmaz IN, Piraino S (2010) New contributions to the jellyfish fauna of the Marmara Sea. Ital J Zool 77:179–185. https://doi.org/10.1080/11250000902895766
İşinibilir-Okyar M, Üstün F, Orun DA (2015) Changes in abundance and community structure of the zooplankton population during the 2008 mucilage event in the northeastern Marmara Sea. Turk J Zool 39:28–38. https://doi.org/10.3906/zoo-1308-11
Kasapidis P, Siokou I, Khelifi-Touhami M et al (2018) Revising the taxonomic status and distribution of the Paracalanus parvus species complex (Copepoda, Calanoida) in the Mediterranean and black seas through an integrated analysis of morphology and molecular taxonomy. J Plankton Res 40(5):595–605. https://doi.org/10.1093/plankt/fby036
Leps J, Smilauer P (2003) Multivariate analysis of ecological data using CANOCO, multivariate analysis of ecological data using CANOCO. Cambridge University Press, New York. https://doi.org/10.1017/cbo9780511615146
Najdek M (1997) Unusual changes of zooplankton fatty acid composition in the northern Adriatic during the 1991 mucilage event. Mar Ecol Prog Ser 159:143–150. https://doi.org/10.3354/meps159143
Öğdül RG, Ergüven H (1992) A study on the abundance and composition of zooplankton groups in the Bay of Beykoz (Bosphorus). Istanbul Univ J Aquat Prod 6:105–120 (in Turkish)
Pielou EC (1966) The measurement of diversity in different types of biological collections. J Theor Biol 13:131–144. https://doi.org/10.1016/0022-5193(66)90013-0
Sever TM (2009) Pelagic Copepoda fauna of the Aegean Sea and the distribution of the common species. Ege J Fish Aquat Sci 26(3):203–209
Shannon CE, Weaver W (1964) The mathematical theory of communication. University of Illinois Press, Urbana, USA
Siegel S (1956) Nonparametric statistics for the behavioral sciences. McGraw-Hill, New York-USA
Simpson EH (1949) Measurement of diversity. Nature 163:688. https://doi.org/10.1038/163688a0
Siokou-Frangou I, Papathanassiou E, Lepretre A, Frontier S (1998) Zooplankton assemblages and influence of environmental parameters on them in a Mediterranean coastal area. J Plankton Res 20:847–870. https://doi.org/10.1093/plankt/20.5.847
Svetlichny LS, Hubareva ES, Kideys AE, İsinibilir M, Shmeleva A (2006) Zooplankton community state in the northeastern Marmara Sea during early autumn with comments on mass mortality of the Black Sea species due to the salinity gradient. J Black Sea/Medit Environ 12(3):213–231
Tarkan AN, Isinibilir M, Tarkan AS (2005) Seasonal variations of the zooplankton composition and abundance in the Istanbul Strait. Pak J Biol Sci 8:1327–1336. https://doi.org/10.3923/pjbs.2005.1327.1336
Ter Braak CJF (1986) Canonical correspondence analysis: a new eigenvector technique for multivariate direct gradient analysis. Ecology 67:1167–1179. https://doi.org/10.2307/1938672
Ter Braak CJF, Smilauer P (1998) CANOCO reference manual and user’s guide to Canoco for windows: software for canonical community ordination (version 4). Centre for Biometry, Wageningen, the Netherlands
Terbiyik Kurt T, Polat S (2015) Zooplankton abundance, biomass, and size structure in the coastal waters of the northeastern Mediterranean Sea. Turk J Zool 39:378–387. https://doi.org/10.3906/zoo-1311-14
Toklu Alıçlı B, Sarihan E (2016) Seasonal changes of zooplankton species and groups composition in Iskenderun Bay (north east Levantine, Mediterranean Sea). Pak J Zool 48(5):1395–1405
Toklu-Alıçlı B, Balkis N, Balci M (2014) Seasonal distribution of zooplankton in the Gulf of Erdek (the Marmara Sea) and the impact of ecological variables. Fresenius Environ Bull 23(12):3013–3021
Tüfekci V, Balkis N, Polat Beken C, Ediger D, Mantikci M (2010) Phytoplankton composition and environmental conditions of a mucilage event in the sea of Marmara. Turk J Biol 34(2):199–210. https://doi.org/10.3906/biy-0812-1
Ullyott P, Pektaş H (1952) Çanakkale Boğazı’ndaki yıllık temperatür ve tuzluluk değişmeleri hakkında ilk araştırmalar. Hidrobiyoloji Mecmuası Serie A:19–33 (in Turkish)
Unal E, Shmeleva A, Zagorodnyaya J, Kideys AE (2000) Zooplankton structure and copepod species of the sea of Marmara in spring 1998. In: Öztürk B, Kadıoğlu M, Öztürk H (eds) Marmara Sea 2000 Symposium. Turkish Marine Research Foundation (TUDAV), Istanbul-Turkey, pp 450–460
Uriarte I, Villate F (2005) Differences in the abundance and distribution of copepods in two estuaries of the Basque coast (Bay of Biscay) in relation to pollution. J Plankton Res 27(9):863–874. https://doi.org/10.1093/plankt/fbi059
Villate F (1994) Temporal variability of the spatial distribution of the zooplankton community in a coastal embayment of the Basque Country in relation to physical phenomena. Hydrobiologia 288:79–95. https://doi.org/10.1007/BF00007128
Wang Y, Zhang W, Lin Y, Cao W, Zheng L, Yang J (2014) Phosphorus, nitrogen and chlorophyll-a are significant factors controlling ciliate communities in summer in the northern Beibu gulf, South China Sea. PLoS One 9(7):e101121. https://doi.org/10.1371/journal.pone.0101121
Weng KC, Pedersen MW, Del Raye GA, Caselle JE, Gray AE (2015) Umbrella species in marine systems: using the endangered humphead wrasse to conserve coral reefs. Endanger Spec Res 27:251–263. https://doi.org/10.3354/esr00663
WoRMS. (2020). Animalia. http://www.marinespecies.org. Accessed 30 May 2020
Yilmaz IN (2015) Collapse of zooplankton stocks during Liriope tetraphylla (hydromedusa) blooms and dense mucilaginous aggregations in a thermohaline stratified basin. Mar Ecol 36:595–610. https://doi.org/10.1111/maec.12166
Yilmaz IN, Isinibilir M (2016) Zooplankton of the Sea of Marmara. In: Özsoy E, Çağatay MN, Balkıs N, Balkıs N, Öztürk B (eds) The sea of Marmara marine biodiversity, fisheries, conservation and governance. Turkish Marine Research Foundation (TUDAV), Istanbul, pp 376–389
Zar J (2010) Biostatistical analysis. Pearson Prentice-Hall, Upper Saddle River, New Jersey, USA
Acknowledgments
The authors are grateful to Prof. Dr. Hüsamettin BALKIS from Istanbul University, Faculty of Science for their valuable assistance. This work was supported by the Research Fund of Istanbul University, Project Number 541.
Funding
This work was supported by the Research Fund of Istanbul University, Project Number 541.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Code availability
Not applicable.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Toklu-Alicli, B., Balkis-Ozdelice, N., Durmus, T. et al. Relationship between environmental factors and zooplankton diversity in the Gulf of Bandırma (the Sea of Marmara). Biologia 76, 1727–1736 (2021). https://doi.org/10.2478/s11756-020-00668-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.2478/s11756-020-00668-8