
Abstract
Background
Natural abundance of carbon (C) and nitrogen (N) stable isotope ratios (δ13C and δ15N) has been used to indicate the state and cycle of ecosystem C and N. However, it is still unclear how C and N cycle of boreal forests respond to the N deposition.
Results
We conducted an 8-year continuous N addition field experiment in a Larix gmelinii forest in Greater Khingan Mountains, Northeast China. Four N treatments (0, 25, 50, 75 kg N ha−1 year−1) were built. The effects of N addition on the δ13C and δ15N of needle, branch, bark, and fine root of Larix gmelinii and soil were studied. The result of the balance between the N input and output flux showed that N addition significantly increased the δ15N in each organ of Larix gmelinii, but did not change the δ15N of soil. We also found that the N absorption by needles of Larix gmelinii could increase the needle photosynthesis rate and δ13C by increasing carboxylation, but N addition had no significant effect on the δ13C of soil and other organs. In addition, both the soil δ15N and δ13C increased with the soil depth.
Conclusions
Long-term N addition may lead to more open C and N cycles and further affect plant nutrient acquisition strategies in boreal forest ecosystems.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Background
Due to fossil fuel combustion and agricultural emissions, anthropogenic nitrogen (N) deposition has been increased dramatically in the past century and is expected to intensify further in the coming decades (Galloway et al. 2008; Liu et al. 2013). Excessive N input can lead to many negative effects, including increased N leaching to water, soil acidification, increased N gas losses from soil and changes in biodiversity (Gao et al. 2014; Chen et al. 2016; Lucander et al. 2021). Therefore, it is necessary to understand the impact direction, intensity and future trend of N deposition on ecosystem C and N cycle, and formulate effective C and N management measures under enhanced global N deposition.
The natural abundance of stable C and N isotope ratios (δ13C and δ15N) is an effective method to explore the C and N dynamics of different ecosystems (Liu et al. 2017; Rao et al. 2017). 15N natural abundance (δ15N) is the comprehensive result of the dynamic process of N cycle (Högberg 1997; Zhang et al. 2018). However, the fractionation of N isotopes in the process of N transformation, absorption, assimilation, distribution, and loss, leads to plants δ15N being different from the δ15N of N source (Bai et al. 2012). Therefore, δ15N of plants can reflect the δ15N of source N and the isotopic fractionation during N cycling processes (Robinson 2001; Kahmen et al. 2008). For example, during nitrification, when the chemical bond of the substrate breaks, the heavier 15N will be transformed into the product more slowly than the lighter 14N, so that the product will have a lower 15N/14N ratio than that of the remaining residual nitrate (Niu et al. 2016; Reis et al. 2016). Therefore, when N saturation causes increased nitrification, soil and plant will become more and more enriched in 15N (Wang et al. 2014). For ecosystem N pool, the N transformation rate is determined by the input and output rates (Högberg et al. 2014). If the increased N input leads to increased N transformation, stronger fractionation and loss of 15N-depleted N, it would affect enrichment of soil and subsequently plant tissue. However, if both the rates of N input and output changes with the increase of N deposition, the responses of plant and soil δ15N values would be complex. Results showed that N additions increased N losses, and the change in plant δ15N after N addition might be related to the balance between higher soil δ15N and 15N depleted N from atmospheric deposition (Kriszan et al. 2009; Gurmesa et al. 2017; Hobbie et al. 2017). Therefore, evaluating the changes in ecosystem δ15N under N addition treatments can provide valuable information on how N flux, assimilation and distribution respond to the increase in N input in forest ecosystems.
Atmospheric air represents a constant C pool with relatively stable isotopic composition and concentration (Galewsky et al. 2016). Therefore, the change in 13C reflects plant physiology: the balance between CO2 supply (conduction) and CO2 demand (assimilation) (Ehleringer 1993). For example, the decrease in stomatal conductance and the restriction of photosynthetic enzyme activity will eventually lead to an increase in plant δ13C value (Jia et al. 2016; Lavergne et al. 2020). Several studies have suggested that foliar uptake of atmospheric reactive N could enhance the rates of photosynthesis and altered foliar δ13C through increased carboxylation (Siegwolf et al. 2001; Guerrieri et al. 2011), although some studies reported no changes in plant δ13C as a result of N enrichment (Wang et al. 2021). Carbon isotope exchange reaction enriches 13C in inorganic salts (such as carbonate), while the dynamic fractionation effect of photosynthesis in the organic cycle of C leads to the 13C-depletion in biogenic C (organic matter), leading to a change in C isotope composition during the exchange of CO2 between organisms and terrestrial ecosystems (Cao et al. 2012; Busch et al. 2020). In the process of microbial decomposition and mineralization of soil organic carbon (SOC) into CO2, the isotope effect is small (only 1–3‰), while strong fraction (10–70‰) occurs during methane production from SOC under anaerobic condition (Conrad et al. 2010). Compared with the fractionation of CO2 fixation during plant photosynthesis, the fractionation during SOC decomposition is almost negligible (Crow et al. 2006). Therefore, by measuring the change in C isotope ratio in each C pool, we can quantify the source, distribution and turnover of C in the “atmosphere–plant–soil” continuum.
Boreal forests account for about 15% of the earth’s total land area and 30% of the forest area, making them the second largest forest biota on land (Vitousek et al. 2002). The boreal forest plays an important role in regulating the global C and N balance and slowing down the rise of greenhouse gas concentration in the atmosphere, and it is also considered to be one of the ecosystems seriously limiting N (Randerson et al. 2006; Houghton 2007). The Greater Khingan Mountains is the only boreal forest zone of China. The widely distributed Larix gmelinii forest is one of the most sensitive vegetation types to global climate change. However, the impacts of N enrichment on the C and N cycle in this ecosystem need to be evaluated. Therefore, we carried out N addition field experiment in Larix gmelinii forest in Greater Khingan Mountains. In 2011, According to the current N deposition rate in northern China (25 kg N ha−1 year−1) (Liu et al. 2013), we established four N addition treatments (0, 25, 50, 75 kg N ha−1 year−1) to simulate the increase of atmospheric N deposition by 0, 1, 2, 3 times in the future. In this experiment, we measured the total C (TC), total N (TN), δ13C and δ15N of needle, branch, bark, and fine root of Larix gmelinii and soil under different N addition treatments. We also measured the photosynthetic rate and photosynthetic enzyme activity. The purpose of this study is to answer the following questions: (1) Whether N addition will change the composition of δ13C and δ15N in plants and soil? (2) What are the effects of different N addition levels on the changes of δ13C and δ15N composition in plants and soil? We hypothesized that (1) N addition will increase the loss of N through gaseous pathway and increase the soil δ15N value. (2) N addition will increase the absorption of N by Larix gmelinii and increase δ15N value in each organ. (3) N addition can increase the δ13C value of Larix gmelinii needles by increasing related processes of photosynthesis.
Methods
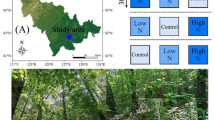
This study was conducted in a natural larch forest of Nanwenghe National Nature Reserve in the Greater Khingan Mountains, Northeast China (51° 05′–51° 39′ N, 125° 07′–125° 50′ E) (Fig. 1). The region has a typical temperate continental climate, with an average temperature of − 2.7 ℃, an annual precipitation of about 500 mm, a plant growth period of about 110 days and an annual sunshine time of 2500 h. The soil is dark brown sandy gravel soil with an average depth of 20 cm. The dominant tree species is Larix gmelinii, whose growth density is 2852 (± 99) trees/ha, and the average diameter at breast height (DBH) was 8.98 (± 0.32) cm, a stand age of 40 years and a canopy density of 0.76 ± 0.30. The vegetation under the forest includes Rhododendron dauricum and Rosa davurica.

Diagram of the experimental design in the Greater Khingan Mountains northeastern China. CK, LN, MN, HN represent the control, low N treatment, medium N treatment, and high N treatment, respectively
In 2011, we randomly set up three sample blocks (Fig. 1), and each sample block includes four 20 m × 20 m plots. The buffer between any two plots is greater than 10 m to avoid interference with nearby plots. We randomly set up four levels of N addition treatment, control (CK, 0 kg N ha−1 year−1), low N (LN, 25 kg N ha−1 year−1), medium N (MN, 50 kg N ha−1 year−1), and high N (HN, 75 kg N ha−1 year−1). The N deposition levels were chosen to simulate a future N deposition of 1–3 folds of the current N deposition level in the study region (25 kg N ha−1 year−1) (Liu et al. 2013). For each N addition plot, NH4NO3 was dissolved in 32 L water in the growing season (from May to September, a total of 5 times each year), and sprayed evenly on the forest floor of each plot using a prayer. To compensate water supply, the control plot was sprayed with the same amount of pure water. The natural abundance of the N fertilizer (NH4NO3) in this study was 0.21% ± 0.43‰.
Sampling method
The sample collection was carried out in mid July 2019. Three trees with similar height and DBH (diameter at breast height) in each plot were randomly selected. Current-year needle, branch, and bark of Larix gmelinii from the top most were cut with a pole pruner. The sampled tree species were marked with numbers. Under the marked trees in each plot, we randomly drilled three soil cores with a soil auger (5 cm in diameter and 20 cm in length), then picked out the root. To obtain soil samples, we randomly selected three points under each labeled tree species, and used a soil auger to get the soil (divided into Oa+e layer and mineral layer, in which the “O” layer is the surface layer of the soil, composed of forest litter, the “a” layer represents the humus layer, and the “e” layer represents the leaching layer). The samples of the same soil layer from each plot were evenly combined to obtain a representative soil sample in each plot. All samples were dried to constant weight at 65 °C and then ground into a fine powder using a Tecator sample mill (Subang, Shanghai, China) for analysis.
Photosynthetic parameter
Three Larix gmelinii trees with similar DBH and height were randomly selected in each plot, with a total of 36 trees (trees for sampling). The healthy needles were selected at different positions (from upper, middle, and lower canopy layers) of the tree crown, and the photosynthetic rate was measured in situ by a hand-held photosynthesis measurement system (CI-340, CID, USA). For each tree, the photosynthesis measurement was done three times in each layer to produce average photosynthetic rate of the whole plant. The photosynthetic rate of all leaves was measured from 9:00 to 11:00 every morning. Because the needles of Larix gmelinii are irregular, it is difficult to accurately determine its leaf area. It is inappropriate to use the photosynthesis per unit area of leaves to express the photosynthetic rate. Therefore, after each measurement of photosynthesis, the measured needles were removed with scissors and weighed to calculate the photosynthetic rate per unit mass. The following equation was used (Qiu 2011):
where Pn (FW) is the photosynthetic rate per unit fresh weight; Pn is the measured value displayed by photosynthetic instrument; S is the leaf chamber area; FW is the fresh weight of leaves in leaf chamber.
Chemical analysis
The C, 13C, N, and 15N contents of all samples were measured on an isotope ratio mass spectrometer (Isoprime 100, Isoprime Ltd.) coupled to an automatic, online elemental analyzer (Vario ISOTOPE cube). The reproducibility of the isotope determination was analyzed in multiple runs at several laboratories using an internal standard soil and plant sample from boreal forests. The accuracy of element content and isotope test is 0.01‰, and the repeated test error (± 2σ) is < 0.02% and < 0.05‰, respectively. Natural abundance δ15N and δ13C in samples were reported in the conventional delta (δ) notation, with units of per mil (‰). When measuring the 13C abundance of the soil, we acidified the soil with 1 N HCl at room temperature for 24 h to remove any inorganic C. The stable isotope ratio (δ15N and δ13C) is expressed as parts per thousand differences from a standard: δ15N/δ13C = (Rsample/Rstandard − 1) × 1000 (‰). Rsample represents the isotope ratio of 15N/14N or 13C/12C in the plant or soil sample, and Rstandard represents the standard ratio of 15N/14N or 13C/12C, usually expressed as 15N/14N of N2 in the atmosphere, Rstandard = 0.011237 is the 13C/12C ratio of international standard (PeeDee Belemnite—PDB).
To determine the content of NH4+–N and NO3−–N in the soil, 5 g fresh soil samples were extracted with 100 mL 0.2 M KCl, oscillated 60 min in the oscillator, and then collected the supernatant. All analyses were performed on a Smartchem instrument 200 (Westco Scientific Instruments, Inc., Italy). Stomatal conductance was measured by hand-held photosynthesis measurement system (CI-340, CID, USA), chlorophyll content and Rubisco activity were measured by ultraviolet–visible spectrophotometer (SHIMADZU UV-1780, Japan).
Data analysis
The Kolmogorov–Smirnov test was used to detect whether the data conformed to the normal distribution, and data homogeneity was detected with Levene’s test. One-way analysis of variance (ANOVA) was used to analyze the effects of N addition on different components C %, δ13C, N %, and δ15N of the soil and Larix gmelinii, and the effects of N addition on needle photosynthesis related parameters. Pearson correlation analysis was used to analyze the correlation between different components C %, δ13C, N %, and δ15N of the soil and Larix gmelinii, and the correlation between photosynthetic related parameters and needle δ13C. All analyses were performed with SPSS 22.0 software package (SPSS, Inc., Chicago, Illinois, USA). Diagrams were drawn using Sigmaplot 13.0 software (Systat Software Inc., Chicago, IL, USA). Tukey’s post hoc test was used to test differences between treatments. Statistically significant differences were set at p < 0.05.
Results
Effects of N addition on the N content and δ15N of plant and soil
Compared with the control, the δ15N and N content of Oa+e layer showed a significant increase under MH and HN treatments (p < 0.05, Table 1). There was no significant difference in the δ15N between 0–10 cm and 10–20 cm soil, but MN and HN treatments significantly increased TN content in 0–10 cm soil (p < 0.05). HN treatment significantly increased NH4+–N and NO3−–N in 0–10 cm soil, while all the three levels of N addition had no significant effect on TN, NH4+–N, and NO3−–N in 10–20 cm soil (Table 1). Under different N addition treatments, the concentration of NH4+–N in Oa+e layer showed significant differences, and the concentration of NH4+–N under HN treatment was significantly higher than that of the control (p < 0.05, Table 1). Besides, MN treatment significantly increased NO3−–N in Oa+e layer (p < 0.05, Table 1).
The δ15N of each organ of Larix gmelinii increased with N addition. MN and HN treatments significantly increased δ15N of needle, bark, and fine root (p < 0.05), while HN treatment significantly increased the δ15N of branch and litter (p < 0.05, Fig. 2A). HN treatment significantly increased TN of needle, bark, and litter (p < 0.05), MN and HN treatments significantly increased TN of fine root (p < 0.05), while N addition had no significant effect on TN of branch (Fig. 2B). The soil δ15N and litter δ15N were significantly negatively correlated with the needle δ15N, while the needle δ15N was significantly positively correlated with the δ15N of Oa+e, bark, branch, litter, and fine root. There was a significant positive correlation between the δ15N of fine root and litter, branch, and bark (Fig. 5).

Effects of nitrogen addition on δ15N value and N concentration of different components of Larix gmelinii. A The δ15N value of different components of Larix gmelinii. B The N concentration of different components of Larix gmelinii. Error bars represent 1 SE of the mean. CK, LN, MN, HN represent the control, low N treatment, medium N treatment, and high N treatment, respectively
Effects of N addition on the C content and δ13C of plant and soil
Nitrogen addition did not significantly change TC and δ13C in Oa+e and 0–20 cm soil layers (Table 1). For Larix gmelinii, N addition significantly increased the δ13C of the needle (p < 0.05). The δ13C of branch and bark decreased with N addition, while the δ13C of litter and fine root increased with N addition (Fig. 3A). The N addition had no significant effect on the TC of organs of Larix gmelinii (Fig. 3B). The TC and TN of Oa+e were significantly positively correlated, and the TC of litter was significantly positively correlated with the δ13C of litter (Fig. 5).

Effects of nitrogen addition on δ13C value and C concentration of different components of Larix gmelinii. A The δ13C value of different components of Larix gmelinii. B The C concentration of different components of Larix gmelinii. Error bars represent 1 SE of the mean. CK, LN, MN, HN represent the control, low N treatment, medium N treatment, and high N treatment, respectively
Effects of N addition on the photosynthetic rate of Larix gmelinii
MN treatment significantly increased the photosynthetic rate of Larix gmelinii (Fig. 4A). HN treatment significantly increased the chlorophyll content (Fig. 4C). LN and MN treatments significantly increased the activity of Rubisco enzyme (Fig. 4D). Nitrogen addition had no significant effect on the stomatal conductance of the needle (Fig. 4B). The photosynthetic rate was significantly positively correlated with Rubisco enzyme, chlorophyll, and needle δ13C (p < 0.05), and chlorophyll and stomatal conductance were significantly positively correlated with the needle δ13C (p < 0.05, Table 2).

Effects of nitrogen addition on photosynthetic related indexes of Larix gmelinii. Error bars represent 1 SE of the mean. A The photosynthetic rate of Larix gmelinii. B The stomatal conductance of Larix gmelinii. C The chlorophyll content of Larix gmelinii. D The Rubisco activity of Larix gmelinii. CK, LN, MN, HN represent the control, low N treatment, medium N treatment, and high N treatment, respectively
Discussion
Effects of N addition on the N content and δ15N of plant and soil
The input process (e.g., N deposition and biological N fixation) and output process (e.g., denitrification, leaching and plant absorption) of N in soil determine the size of N pool (Dawson et al. 2002; Fernandez-Alonso et al. 2018; Rivero-Villar et al. 2021). In this study, addition of N with δ15N close to that of atmospheric N2 (≈ 0), significantly increased the δ15N of Larix gmelinii (Fig. 2). Our results showed that the N addition significantly increased the soil TN (Table 1), but had no significant effect on the δ15N of soil mineral horizon, which is inconsistent with previous studies (Gurmesa et al. 2017; Wang et al. 2021) and our first hypothesis. The δ15N of soil is regulated by N input and N output fluxes (Liu et al. 2017). Based on the mass balance model of Brenner et al. (2001), under the condition of constant N output flux, N addition reduces the δ15N of soil. Therefore, the lack of significant changes in soil δ15N after long-term N addition is likely to be the result of the balance between input and output flux. This is probably due to the loss of 15N-depleted N through denitrification and leaching. Under such condition, the residual soil (and plant) would be 15N-enriched. MN and HN treatments significantly increased NH4+–N and NO3––N, and increased soil mineralization rate (Table 1), which might lead to increased denitrification and leaching. Gaseous losses of 15N-depleted N by denitrification would enrich 15N in soil N (Houlton et al. 2006). It has been proved that NH3 volatilization process will cause 40–60‰ isotope fractionation effect (Robinson 2001; Liu et al. 2017). MN and HN treatments significantly increased the δ15N of Oa+e, which might be because we added a relatively 15 N-enriched N to the Oa+e that was relatively 15N-depleted. In addition, soil organic matter rich in 14N was decomposed during soil microbial decomposition, resulting in soil 15N enrichment (Bostrm et al. 2007). 15N enriched microorganisms and microbial residues are produced in the process of microbial assimilation for organic matter, which will also lead to the gradual enrichment of 15N in soil organic matter over time (Dijkstra et al. 2010).
Nitrogen addition significantly increased the δ15N in plant compartments (Fig. 2), and the δ15N in needles was positively correlated with branch, bark, fine root, litter, and humus (Fig. 5), which was consistent with our second hypothesis. The increased plant δ15N as a result of N addition could be the imprint of the added fertilizer N which was 15N-enriched compared to the plant tissues (Gurmesa et al. 2017). Our results also showed that the increase of δ15N of Larix gmelinii was directly related to the soil δ15N (Fig. 5). In the process of N cycle, N addition leads to an increase in plant N utilization rate, and then increases the δ15N of each organ of Larix gmelinii due to isotope fractionation (Sheng et al. 2019). In addition, when the soil N availability is low, plants may be more dependent on mycorrhizal fungi (Vallano and Sparks 2013). Mycorrhizal fungi usually transfer 15N depleted N to plants, resulting in high enrichment of 15N in mycorrhizal mycelium, resulting in 15N depletion in plant tissues (Schweiger, 2016). The δ15N of fine roots of Larix gmelinii is significantly lower than that of needle, branch, and bark (Fig. 2), which may be caused by the fractionation effect of mycorrhizal. The increase of δ15N in needles can also be used to indicate the increase of N cycle rate. Previous studies have also shown that there is a strong correlation between the enrichment level of 15N in leaves and the increase in N cycle rate (Garten 1993; Emmett et al. 1998). The greater the difference in δ15N between the shoot and the root, the greater the proportion of inorganic N absorbed by the leaves (Kalcsits et al. 2014).

Pearson correlation coefficients among the C, N, δ13C, and δ15N of different compartments of Larix gmelinii and soil. * represents p < 0.05, ** represents p < 0.01
We also found that the soil δ15N increased with an increase in soil depth, which is consistent with a large number of studies (Kramer et al. 2017; Högberg et al. 2019; Potapov et al. 2019). This may be because 15N depleted litter and plant residues enter the surface soil, and the accumulation of 15N enriched mycorrhizal fungal residues in the deep soil can lead to a large enrichment of 15N in the deep soil relative to the surface litter.
Effects of N addition on the C content and δ13C of plant and soil and photosynthetic rate
Our results showed that N addition significantly increased the δ13C of needle, but had no significant effect on the δ13C of other plant organs (Fig. 3). The C isotope fractionation caused by photosynthesis mainly takes place in the following two steps: first, there is a certain resistance when 13CO2 enters mesophyll cells. Due to the influence of kinetic fractionation effect, 12CO2 diffuses faster and is easily absorbed by plants; Second, under the action of carboxyl polypeptidase, 12CO2 dissolved in cells and Rubisco carboxylase are preferentially combined into phosphoglyceric acid, while the remaining CO2 is enriched in 13C (Park and Epstein 1960; Flexas et al. 2006; Ulli et al. 2008). Previous studies have shown that the decrease in plant stomatal conductance leads to a decrease of CO2 concentration in plant leaves and the increase of δ13C in leaves (Morecroft and Woodward 1990; Rao et al. 2017). In our study, N addition had no significant effect on stomatal conductance (Fig. 4B). Therefore, we speculate that the increase in needle δ13C may be caused by the assimilation of 12CO2. The increase of 13C in needles could be explained by the addition of N increased Rubisco activity (Fig. 4D) and chlorophyll (Fig. 4C), increased photosynthetic rate (Fig. 4A), and gradually increased the consumption of 12CO2, which is consistent with our third hypothesis. Tomaszewski and Sieving (2007) also showed that N addition increased photosynthetic rate, which could enhance CO2 assimilation but also reduce the thermally dissipated light.
Our previous results found that N addition significantly increased CH4 and CO2 emissions from the Larix gmelinii forest (Yan et al. 2018), and the increase of C output flux usually led to the enrichment of soil 13C. The lack of significant change in soil 13C in response to N addition (Fig. 3A) could be a trade-off between increased levels of plant residue (more 13C depleted than soil C) inputs and 13C enrichment from enhanced soil C decomposition. Litter, fine root, and root exudates are the main sources of C input to soil (Waldrop et al. 2004; Cusack et al. 2010). The decrease of litter δ13C (Fig. 3A) may be related to the fractionation effect of C release during litter decomposition. Easily decomposable organic compounds (such as sugar, starch and cellulose) in litter usually show high δ13C value. In the process of microbial decomposition, this leads to the loss of 12C faster than 13C, resulting in the enrichment of 13C in litter (Cotrufo et al. 2005; Bowling et al. 2008). The retention efficiency of root C in soil is much higher than that of litter C (Clemmensen et al. 2013). Therefore, the change in equilibrium δ13C output may mainly come from the input of roots and root exudates. Gougherty et al. (2018) found that the greater allocation of relatively 13C depleted (i.e., lower δ13C values) non-structure carbohydrate (NSC) from leaves to roots when the photosynthesis was high. Our previous studies also showed that N addition increased the biomass of underground roots of Larix gmelinii and strengthened the transport function of fine roots to overcome the P limitation (Liu et al. 2021). A meta-analysis by Finzi et al. (2015) showed that the contribution of root C input to the C–N mineralization process of forest soil was as high as 33%. The study by Bais et al. (2006) also found that up to 40% of photosynthetic fixed C was secreted into the rhizosphere. And high nutrient availability can promote the release of fine root exudates (Neumann and Romheld 2000). Although we have not measured root exudates, the fractionation effect of root exudates on soil C input cannot be ignored.
Gessler et al. (2012) showed that the difference of δ13C in leaves and δ13C in phloem of branch reached 30%, due to the fractionation effect of photosynthesis. In our results, although there is no significant change in δ13C of branch and bark, it shows a decreasing trend with the N addition, and the δ13C of branch and bark is also 8.4–13% lower than that of needle (Fig. 3A), which may be due to the increased transport of organic matter rich in 12C to the aboveground.
Conclusions
Our study shows that long-term N addition changes the C and N cycle of boreal forest ecosystem. The N addition increased the δ15N value and TN of each organ of Larix gmelinii and also increased the N mineralization of soil, but had no significant effect on the soil δ15N. Compared with N, the C concentration and 13C of Larix gmelinii were less affected. The N addition only increased the δ13C of needle, which was the result of the increase of photosynthetic efficiency of needle. By exploring the stable C and N isotopic signatures in plant–soil compartments in boreal forest ecosystems, this study confirms that the C and N state and cycle of ecosystem may be fluctuated under the projected N enrichment.
Availability of data and materials
Data are available from the corresponding author on reasonable request.
References
Bai E, Houlton BZ, Wang YP (2012) Isotopic identification of nitrogen hotspots across natural terrestrial ecosystems. Biogeosciences 9(8):3287–3304. https://doi.org/10.5194/bg-9-3287-2012
Bais HP, Weir TL, Perry LG et al (2006) The role of root exudates in rhizosphere interactions with plants and other organisms. Annu Rev Plant Biol 57:233–266. https://doi.org/10.1146/annurev.arplant.57.032905.105159
Bostrm B, Comstedt D, Ekblad A (2007) Isotope fractionation and 13C enrichment in soil profiles during the decomposition of soil organic matter. Oecologia 153(1):89–98. https://doi.org/10.1007/s00442-007-0700-8
Bowling DR, Pataki DE, Randerson JT (2008) Carbon isotopes in terrestrial ecosystem pools and CO2 fluxes. New Phytol 178(1):24–40. https://doi.org/10.1111/j.1469-8137.2007.02342.x
Brenner DL, Amundson R, Baisden WT et al (2001) Soil N and 15N variation with time in a California annual grassland ecosystem. Geochim Cosmochim Acta 65(22):4171–4186. https://doi.org/10.1016/S0016-7037(01)00699-8
Busch FA, Holloway-Phillips MH, Stuart-Williams H et al (2020) Revisiting carbon isotope discrimination in C3 plants shows respiration rules when photosynthesis is low. Nat Plants 6(3):245–258. https://doi.org/10.1038/s41477-020-0606-6
Cao X, Jia JB, Li H et al (2012) Photosynthesis, water use efficiency and stable carbon isotope composition are associated with anatomical properties of leaf and xylem in six poplar species. Plant Biol 14(4):612–620. https://doi.org/10.1111/j.1438-8677.2011.00531.x
Chen D, Li J, Lan Z et al (2016) Soil acidification exerts a greater control on soil respiration than soil nitrogen availability in grasslands subjected to long-term nitrogen enrichment. Funct Ecol 30(4):658–669. https://doi.org/10.1111/1365-2435.12525
Clemmensen KE, Bahr A, Ovaskainen O et al (2013) Roots and associated fungi drive long-term carbon sequestration in boreal forest. Science 339(6127):1615–1618. https://doi.org/10.1126/science.1231923
Conrad R, Klose M, Claus AP et al (2010) Methanogenic pathway, 13C isotope fractionation, and archaeal community composition in the sediment of two clear-water lakes of Amazonia. Limnol Oceanogr 55(2):689–702. https://doi.org/10.4319/lo.2010.55.2.0689
Cotrufo MF, Drake B, Ehleringer JR (2005) Palatability trials on hardwood leaf litter grown under elevated CO2: a stable carbon isotope study. Soil Biol Biochem 37(6):1105–1112. https://doi.org/10.1016/j.soilbio.2004.11.009
Crow SE, Sulzman EW, Rugh WD et al (2006) Isotopic analysis of respired CO2 during decomposition of separated soil organic matter pools. Soil Biol Biochem 38(11):3279–3291. https://doi.org/10.1016/j.soilbio.2006.04.007
Cusack DF, Torn MS, McDowell WH et al (2010) The response of heterotrophic activity and carbon cycling to nitrogen additions and warming in two tropical soils. Glob Change Biol 16(9):2555–2572. https://doi.org/10.1111/j.1365-2486.2009.02131.x
Dawson TE, Mambelli S, Plamboeck AH, Templer PH, Tu KP (2002) Stable isotopes in plant ecology. Annu Rev Ecol Syst 33(1):507–559. https://doi.org/10.1146/annurev.ecolsys.33.020602.095451
Dijkstra P, Laviolette CM, Coyle JS et al (2010) 15N enrichment as an integrator of the effects of C and N on microbial metabolism and ecosystem function. Ecol Lett 11(4):389–397. https://doi.org/10.1111/j.1461-0248.2008.01154.x
Ehleringer JR (1993) Variation in leaf carbon isotope discrimination in Encelia farinosa: implications for growth, competition, and drought survival. Oecologia 95(3):340–346. https://doi.org/10.1007/BF00320986
Emmett BA, Kjønaas OJ, Gundersen P, Koopmans C, Tietema A, Sleep D (1998) Natural abundance of 15N in forests across a nitrogen deposition gradient. For Ecol Manage 101(1–3):9–18. https://doi.org/10.1016/S0378-1127(97)00121-7
Fernandez-Alonso MJ, Yuste JC, Kitzler B et al (2018) Changes in litter chemistry associated with global change-driven forest succession resulted in time-decoupled responses of soil carbon and nitrogen cycles. Soil Biol Biochem 120:200–211. https://doi.org/10.1016/j.soilbio.2018.02.013
Finzi AC, Abramoff RZ, Spiller KS et al (2015) Rhizosphere processes are quantitatively important components of terrestrial carbon and nutrient cycles. Glob Change Biol 21(5):2082–2094. https://doi.org/10.1111/gcb.12816
Flexas JM, Ribas-Carbó BJ, Bota J et al (2006) Decreased Rubisco activity during water stress is not induced by decreased relative water content but related to conditions of low stomatal conductance and chloroplast CO2 concentration. New Phytol 172(1):73–82. https://doi.org/10.1111/j.1469-8137.2006.01794.x
Galewsky J, Steen-Larsen HC, Field RD, Worden J, Risi C, Schneider M (2016) Stable isotopes in atmospheric water vapor and applications to the hydrologic cycle. Rev Geophys 54(4):809–865. https://doi.org/10.1002/2015RG000512
Galloway JN, Townsend AR, Erisman JW et al (2008) Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science 320(5878):889–892. https://doi.org/10.1126/science.1136674
Gao Q, Hasselquist NJ, Palmroth S et al (2014) Short-term response of soil respiration to nitrogen fertilization in a subtropical evergreen forest. Soil Biol Biochem 76:297–300. https://doi.org/10.1016/j.soilbio.2014.04.020
Garten CT (1993) Variation in foliar 15N abundance and the availability of soil nitrogen on Walker Branch watershed. Ecology 74(7):2098–2113. https://doi.org/10.2307/1940855
Gessler A, Tcherkez G, Peuke AD et al (2012) Experimental evidence for diel variations of the carbon isotope composition in leaf, stem and phloem sap organic matter in Ricin. Plant Cell Environ 35:1245–1257. https://doi.org/10.1111/j.1365-3040.2008.01806.x
Gougherty SW, Bauer JE, Pohlman JW (2018) Exudation rates and δ13C signatures of tree root soluble organic carbon in a riparian forest. Biogeochemistry 137(1):235–252. https://doi.org/10.1007/s10533-017-0415-9
Guerrieri R, Mencuccini M, Sheppard LJ et al (2011) The legacy of enhanced N and S deposition as revealed by the combined analysis of δ13C, δ18O and δ15N in tree rings. Glob Chang Biol 17:1946–1962. https://doi.org/10.1111/j.1365-2486.2010.02362.x
Gurmesa GA, Lu X, Gundersen P et al (2017) Nitrogen input 15N signatures are reflected in plant 15N natural abundances in subtropical forests in China. Biogeosciences 14(9):2359–2370. https://doi.org/10.5194/bg-14-2359-2017
Hobbie EA, Chen J, Hanson PJ et al (2017) Long-term carbon and nitrogen dynamics at SPRUCE revealed through stable isotopes in peat profiles. Biogeosciences 14(9):2481–2494. https://doi.org/10.5194/bg-2016-261
Högberg P (1997) 15N natural abundance in soil-plant systems. New Phytol 137(2):179–203
Högberg MN, Skyllberg U, Hgberg P et al (2019) Does ectomycorrhiza have a universal key role in the formation of soil organic matter in boreal forests? Soil Biol Biochem 140:107635. https://doi.org/10.1016/j.soilbio.2019.107635
Högberg P, Johannisson C, Högberg MN (2014) Is the high 15N natural abundance of trees in N-loaded forests caused by an internal ecosystem N isotope redistribution or a change in the ecosystem N isotope mass balance? Biogeochemistry 117(2–3):351–358. https://doi.org/10.1007/s10533-013-9873-x
Houghton RA (2007) Balancing the global carbon budget. Annu Rev Earth Planet Sci 35(1):313–347. https://doi.org/10.1146/annurev.earth.35.031306.140057
Houlton BZ, Sigman DM, Hedin LO (2006) Isotopic evidence for large gaseous nitrogen losses from tropical rainforests. Proc Natl Acad Sci 103(23):8745–8750. https://doi.org/10.1073/pnas.0510185103
Jia Y, Wang G, Tan Q et al (2016) Temperature exerts no influence on organic matter δ13C of surface soil along the 400mm isopleth of mean annual precipitation in China. Biogeosciences 13(17):5057–5064. https://doi.org/10.5194/bg-13-5057-2016
Kahmen A, Wanek W, Buchmann N (2008) Foliar δ15N values characterize soil N cycling and reflect nitrate or ammonium preference of plants along a temperate grassland gradient. Oecologia 156:861–870. https://doi.org/10.1007/s00442-008-1028-8
Kalcsits LA, Buschhaus HA, Guy RD (2014) Nitrogen isotope discrimination as an integrated measure of nitrogen fluxes, assimilation and allocation in plants. Physiol Plant 151(3):293–304. https://doi.org/10.1111/ppl.12167
Kramer MG, Lajtha K, Aufdenkampe AK (2017) Depth trends of soil organic matter C:N and 15N natural abundance controlled by association with minerals. Biogeochemistry 136(3):237–248. https://doi.org/10.1007/s10533-017-0378-x
Kriszan M, Amelung W, Schellberg J et al (2009) Long-term changes of the δ15N natural abundance of plants and soil in a temperate grassland. Plant Soil 325(1–2):157–169. https://doi.org/10.1007/s11104-009-9965-5
Lavergne A, Sandoval D, Hare VJ et al (2020) Impacts of soil water stress on the acclimated stomatal limitation of photosynthesis: insights from stable carbon isotope data. Glob Change Biol 26(12):7158–7172. https://doi.org/10.1111/gcb.15364
Liu X, Zhang Y, Han W et al (2013) Enhanced nitrogen deposition over China. Nature 494:459–462. https://doi.org/10.1038/nature11917
Liu J, Wang C, Peng B et al (2017) Effect of nitrogen addition on the variations in the natural abundance of nitrogen isotopes of plant and soil components. Plant Soil 412(1–2):453–464. https://doi.org/10.1007/s11104-016-3081-0
Liu G, Xing Y, Wang Q et al (2021) Long-term nitrogen addition regulates root nutrient capture and leaf nutrient resorption in Larix gmelinii in a boreal forest. Eur J For Res 140(4):763–776. https://doi.org/10.1007/s10342-021-01364-1
Lucander K, Zanchi G, Akselsson C et al (2021) The effect of nitrogen fertilization on tree growth, soil organic carbon and nitrogen leaching—a modeling study in a steep nitrogen deposition gradient in sweden. Forests 12(3):298. https://doi.org/10.3390/f12030298
Morecroft MD, Woodward FI (1990) Experimental investigations on the environmental determination of δ13C at different altitudes. J Exp Bot 10:1303–1308. https://doi.org/10.1093/jxb/41.10.1303
Neumann G, Romheld V (2000) The release of root exudates as affected by the plant's physiological status. In: The rhizosphere, CRC Press, pp 57–110
Niu SL, Classen AT, Dukes JS et al (2016) Global patterns and substrate-based mechanisms of the terrestrial nitrogen cycle. Ecol Lett 19(6):697–709. https://doi.org/10.1111/ele.12591
Park R, Epstein S (1960) Carbon isotope fractionation during photosynthesis. Geochim Cosmochim Acta 21(1):110–126. https://doi.org/10.1016/S0016-7037(60)80006-3
Potapov AM, Tiunov AV, Stefan S (2019) Uncovering trophic positions and food resources of soil animals using bulk natural stable isotope composition. Biol Rev 94(1):37–59. https://doi.org/10.1111/brv.12434
Qiu N (2011) The method of determining photosynthetic rate based on fresh weight of Pinaceae conifer. For Sci Technol 36(4):18–20
Randerson J, Liu H, Flanner M et al (2006) The impact of boreal forest fire on climate warming. Science 314(5802):1130–1132. https://doi.org/10.1126/science.1132075
Rao Z, Guo W, Cao J et al (2017) Relationship between the stable carbon isotopic composition of modern plants and surface soils and climate: a global review. Earth-Sci Rev 165:110–119. https://doi.org/10.1016/j.earscirev.2016.12.007
Reis C, Nardoto GB, Rochelle A et al (2016) Nitrogen dynamics in subtropical fringe and basin mangrove forests inferred from stable isotopes. Oecologia 183(3):841–848. https://doi.org/10.1007/s00442-016-3789-9
Rivero-Villar A, Ruiz-Suárez G, Templer PH et al (2021) Nitrogen cycling in tropical dry forests is sensitive to changes in rainfall regime and nitrogen deposition. Biogeochemistry 153:283–302. https://doi.org/10.1007/s10533-021-00788-6
Robinson D (2001) δ15N as an integrator of the nitrogen cycle. Trends Ecol Evol 16(3):153–162. https://doi.org/10.1016/S0169-5347(00)02098-X
Schweiger PF (2016) Nitrogen isotope fractionation during N uptake via arbuscular mycorrhizal and ectomycorrhizal fungi into grey alder. J Plant Physiol 205:84–92. https://doi.org/10.1016/j.jplph.2016.08.004
Sheng Z, Huang Y, He K et al (2019) Responses of plant 15N natural abundance and isotopic fractionation to N addition reflect the N status of a temperate steppe in China. J Plant Ecol 12(3):550–563. https://doi.org/10.1093/jpe/rty047
Siegwolf R, Matyssek R, Saurer M et al (2001) Stable isotope analysis reveals differential effects of soil nitrogen and nitrogen dioxide on the water use efficiency in hybrid poplar leaves. New Phytol 149(2):233–246. https://doi.org/10.1046/j.1469-8137.2001.00032.x
Tomaszewski T, Sievering H (2007) Canopy uptake of atmospheric N deposition at a conifer forest: part II - response of chlorophyll fluorescence and gas exchange parameters. Tellus Ser B-Chem Phys Meteorol 59(3):493–501. https://doi.org/10.1111/j.1600-0889.2007.00265.x
Ulli S, Rajabi A, Griffiths H, Berry JA (2008) Carbon isotopes and water use efficiency: sense and sensitivity. Oecologia 155(3):441–454. https://doi.org/10.1007/s00442-007-0932-7
Vallano DM, Sparks JP (2013) Foliar δ15N is affected by foliar nitrogen uptake, soil nitrogen, and mycorrhizae along a nitrogen deposition gradient. Oecologia 172(1):47–58. https://doi.org/10.1007/s00442-012-2489-3
Vitousek PM, Cassman K, Cleveland C et al (2002) Towards an ecological understanding of biological nitrogen fixation. Biogeochemistry 57(1):1–45. https://doi.org/10.1023/A:1015798428743
Waldrop MP, Zak DR, Sinsabaugh RL et al (2004) Nitrogen deposition modifies soil carbon storage through changes in microbial enzymatic activity. Ecol Appl 14(4):1172–1177. https://doi.org/10.1890/03-5120
Wang C, Wang X, Liu D et al (2014) Aridity threshold in controlling ecosystem nitrogen cycling in arid and semi-arid grasslands. Nat Commun 5:4799. https://doi.org/10.1038/ncomms5799
Wang R, Peuelas J, Li T et al (2021) Natural abundance of 13C and 15N provides evidence for plant-soil carbon and nitrogen dynamics in a N-fertilized meadow. Ecology 102(6):e03348. https://doi.org/10.1002/ecy.3348
Yan G, Xing Y, Wang J et al (2018) Sequestration of atmospheric CO2 in boreal forest carbon pools in northeastern China: effects of nitrogen deposition. Agric For Meteorol 248:70–81. https://doi.org/10.1016/j.agrformet.2017.09.015
Zhang ZL, Li N, Xiao J et al (2018) Changes in plant nitrogen acquisition strategies during the restoratioN of spruce plantations on the eastern Tibetan Plateau, China. Soil Biol Biochem 119:50–58. https://doi.org/10.1016/j.soilbio.2018.01.002
Acknowledgements
We gratefully acknowledge Professor Yunting Fang and Dr. Feifei Zhu from the Institute of Applied Ecology, Chinese Academy of Sciences for their comments and suggestions on an earlier draft of this manuscript.
Funding
This research was supported by grants from the National Natural Science Foundation of China (42230106, 41773075, 41575137) and Open Grant for Key Laboratory of Sustainable Forest Ecosystem Management (Northeast Forestry University), Ministry of Education (KFJJ2021ZD01).
Author information
Authors and Affiliations
Contributions
QW and YX designed the study, were awarded funding, supervised data collection and contributed to and edited manuscripts. QW, GL, YX, ZY, GY, SL and XW contributed the whole manuscript preparation and design and wrote the main manuscript text. QW, GL, YX, ZY, GY and SL prepared all figures, GL, YX, ZY, GY, SL, XW and QW prepared field experiments, tables and collected literatures. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflict of interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Liu, G., Yin, Z., Yan, G. et al. Effects of long-term nitrogen addition on the δ15N and δ13C of Larix gmelinii and soil in a boreal forest. Ecol Process 11, 37 (2022). https://doi.org/10.1186/s13717-022-00382-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13717-022-00382-0