
Abstract
Non-alcoholic fatty liver disease (NAFLD) is becoming a significant global public health threat. Seabuckthorn (Hippophae rhamnoides L.) has been used in traditional Chinese medicine (TCM). The hypolipidemic effects of Seabuckthorn polysaccharides (SP) against high-fat diets (HFD)-induced NAFLD were systematically explored and compared with that of Bifidobacterium lactis V9 (B. Lactis V9). Results showed that HFD-induced alanine transaminase (ALT) and aspartate aminotransferase (AST) levels decreased by 2.8-fold and 4.5-fold, respectively, after SP supplementation. Moreover, the alleviating effect on hepatic lipid accumulation is better than that of B. Lactis V9. The ACC and FASN mRNA levels were significantly reduced by 1.8 fold (P < 0.05) and 2.3 folds (P < 0.05), respectively, while the CPT1α and PPARα mRNA levels was significantly increased by 2.3 fold (P < 0.05) and 1.6 fold (P < 0.05), respectively, after SP administration. SP activated phosphorylated-AMPK and inhibited PPARγ protein expression, improved serum oxidative stress and inflammation (P < 0.05). SP supplementation leads to increased hepatic expression of nuclear factor erythroid 2-related factor 2 (Nrf-2), heme oxygenase-1 (HO-1) and Superoxide dismutase-2 (SOD-2). Furthermore, SP treatment improved HFD-induced intestinal dysbiosis. Lentisphaerae, Firmicutes, Tenericutes and Peptococcus sp., RC9_gut_group sp., and Parabacteroides sp. of the gut microbiota were significantly associated with hepatic steatosis and indicators related to oxidative stress and inflammation. Therefore, SP can mitigate hepatic lipid accumulation by regulating Nrf-2/HO-1 signaling pathways and gut microbiota. This study offers new evidence supporting the use of SP as a prebiotic treatment for NAFLD.
Key points
-
SP supplementation mitigates HFD-induced hyperlipidemia and hepatic lipid accumulation.
-
SP elevated the levels of phosphorylated-AMPK and PPARγ protein expression.
-
SP promoted the hepatic anti-oxidative response by upregulating the Nrf-2/HO-1 signaling pathway.
-
SP restores HFD-induced dysbiosis in gut microbiota.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Non-alcoholic fatty liver disease (NAFLD) has become a significant public health concern in many countries (Gong et al. 2017). Its main feature is the abnormal fat accumulation in more than 5% of liver cells (mainly in the form of triglycerides). The histological feature of NAFLD includes microvesicular or macrovesicular steatosis, which is not triggered by excessive alcohol consumption or other clearly defined causes of liver injury (Koo 2013; Friedman et al. 2018; Idalsoaga et al. 2020). Some NAFLD patients may progress from simple hepatic steatosis to non-alcoholic steatohepatitis (NASH) which is characterized by inflammation, hepatocellular ballooning and fibrosis. NASH is at the risk of developing cirrhosis or hepatocellular carcinoma (Kwok et al. 2016; de Vries et al. 2020). In addition, NAFLD is closely associated with metabolic disorders such as obesity, type 2 diabetes, insulin resistance and dyslipidemia (Friedman et al. 2018). However, in addition to exercise and calorie restriction, there are currently no specific pharmaceutical treatments for NAFLD (Xue et al. 2024b). Hence, there is an urgent need to explore new agents with both hypolipidemic and anti-inflammatory effects for treating NAFLD and its complications.
In the occurrence and development of NAFLD, high-fat and high-calorie intake and insulin resistance leads to increased lipid deposition in the liver, resulting in oxidative stress, mitochondrial dysfunction, lipid peroxidation and endoplasmic reticulum stress that promote inflammatory responses in liver cells, ultimately leading to cirrhosis and liver cancer (James et al. 1998). AMP-activated protein kinase (AMPK) is an energy sensor that plays an important role in ameliorating NAFLD de novo lipid synthesis and boosting fatty acid oxidation (Smith et al. 2016; Fang et al. 2022). Researches have demonstrated a notable decrease in both the expression and activity of AMPK within the liver among individuals with metabolic diseases (Xu et al. 2024; Zou et al. 2024). Peroxisome proliferator-activated receptors (PPARs) are ligand-activated receptors belonging to the nuclear hormone receptor superfamily, regulating glucose and lipid metabolism, inflammation, and fibrosis (Francque et al. 2021). PPARγ plays a crucial role in lipid synthesis and hepatic inflammation (Cariello et al. 2021). It has been reported that PPARγ expression is significantly increased in the NAFLD models (Puengel et al. 2022). On the contrary, the PPARγ antagonist GW9662 can improve lipid metabolism in NASH mice (Xiao et al. 2023). Nuclear factor erythroid 2-related factor 2 (Nrf-2), a key transcription factor regulating intracellular antioxidant responses, plays a critical role in regulating lipid peroxidation and cell metabolism (Martín-Fernández et al. 2022; Ngo et al. 2022). Heme oxygenase-1 (HO-1), a target gene of Nrf-2, mediates anti-oxidative and anti-inflammatory effects in response to cellular stress and diverse oxidative stimuli (Shaw et al. 2020). Recent reports have shown that the HO-1 agonist oxyberberine could alleviate oxidative stress in mice with acute liver injury by activating the Nrf-2 signaling pathway (Ai et al. 2022). In addition, bioactive dipeptides can also alleviate metabolic fatty liver disease by up-regulating the Nrf-2/HO-1 signaling pathway (Wayal et al. 2023).
The gut microbiota and its metabolites play crucial roles in many metabolic diseases, such as obesity, type 2 diabetes, NAFLD and cardiovascular disease (Wu et al. 2021b). Studies have shown that commensal Bacteroides fragilis exacerbates NAFLD by inducing intestinal microbiota dysbiosis (Huang et al. 2024). Supplementation of the gut microbial metabolite indole-3-acetate could alleviate diet-induced hepatic steatosis and inflammation in mice (Ding et al. 2024). Recently, growing researches indicate that the traditional Chinese herbs may alleviate metabolic diseases by modulation of gut microbiota (Bao et al. 2022).
Seabuckthorn (Hippophae rhamnoides L.) has a long history of medicinal use in China, as documented in “Si Bu Yi Dian”. Seabuckthorn polysaccharide (SP) is the main active component in Seabuckthorn berries, which exhibits a wide range of pharmacological activities such as liver protection, anti-inflammatory, antioxidant, and immune-enhancing effects (Yuandangongbu 1983). Our previous study has demonstrated that SP could mitigate carbon tetrachloride-induced hepatic injury via its antioxidant and anti-inflammatory activities (Zhang et al. 2017). SP can also alleviate acetaminophen-induced hepatotoxicity by activating the Nrf-2/HO-1 signaling pathway (Wang et al. 2018). Whether SP exhibits beneficial effects against high-fat diet (HFD)-induced NAFLD are not yet known. Given the multitude of studies showcasing the gut microbiota-modulating effects of polysaccharides, there is significant potential for their utilization as prebiotics in the prevention of metabolic associated fatty liver disease (MAFLD) (Guo et al. 2023). Our previous study has shown that Bifidobacterium lactis V9 (B. Lactis V9) can alleviate high-fat diet-induced NAFLD by modulating Adenosine 5’-monophosphate (AMP)-activated protein kinase (AMPK) and TLR-NF-κB signaling pathways (Yan et al. 2020). Whether SP and B. Lactis V9 can modulate the gut microbiota to alleviate NAFLD remains largely unknown. This study aims to investigate the beneficial effects and underlying mechanisms of SP against HFD-induced NAFLD. The inclusion of B. Lactis V9 in the study is depicted for comparative purposes. The study could offer a theoretical foundation for the development of SP as a prebiotic preparation for preventing metabolic diseases.
Materials and methods
Reagents
Berberine hydrochloride (purity > 98%) was purchased from Aladdin Reagent Co., Ltd, Shanghai, China. Rat interleukin-1β (IL-1β), interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α) enzyme-linked immunosorbent assay (ELISA) kits were purchased from Xinbosheng Biotechnology, Shenzhen, China. Superoxide dismutase (SOD) and malondialdehyde (MDA) assay kits were purchased from Nanjing Jianchen Bioengineering Institute, Nanjing, China. BCA protein assay kit was purchased from Shanghai Bioengineering Co. Ltd, China. Two types of diets were obtained from Beijing Keao Xieli Feed Co.,Ltd, China: a standard diet containing 12% fat energy (RD) and a high-fat diet containing 35% fat energy (HFD). Trizol reagent was purchased from Tiangen Biochemical Technology Co., Ltd, Beijing, China. Reverse transcription PCR kit was purchased from TaKaRa Biotechnology Co., Ltd, Dalian, China. Antibodies against Nrf-2 (ab31163), heme oxygenase-1 (HO-1) (ab189491) and glutamate-cysteine ligase catalytic subunit (GCLC) (ab207777) were purchased from Abcam (Cambridge, UK). Superoxide dismutase-2 (SOD-2) (WL02506) and peroxisome proliferator-activated receptor γ (PPARγ) (WL01800) antibodies were purchased from Shenyang Wanlei Biotechnology Co, Ltd, China.
Preparation of SP and B. Lactis V9
The detailed extraction procedure of Seabuckthorn polysaccharides (SP) was described in previous experiments. HPLC analysis showed that SP is composed of mannose, arabinose, glucose, galactose, and rhamnose with a molecular ratio of 2.02:1.02:4.24:1:9.22. The average molecular weight (MW) of SP is 275,420 Da (Zhang et al. 2017). The preparation of B. Lactis V9 was carried out according to the procedures as we described previously (Sun et al. 2010).
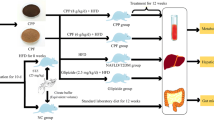
Animal experiments
Forty-eight male Wistar rats (120–140 g) were acquired from Beijing Viton Lever Laboratory Animal Technology Co. Ltd (Beijing, China). The rats were kept in a controlled environment with a temperature of 24 ± 1 ℃, relative humidity of 55 ± 5%, and a 12-hour light/dark cycle. After one week of acclimatization, rats were randomly divided into six groups (n = 8): control (Ctrl), high-fat diet (HFD), HFD + berberine (HFD + Ber), HFD + SP and HFD + B. Lactis V9 (HFD + V9) groups. Rats were maintained on HFD or a chow diet for six weeks. After six weeks, rats in the HFD + SP group received a dose of SP solution (200 mg/kg) via gavage (Liu et al. 2015), rats in the HFD + Ber group were given berberine hydrochloride (300 mg/kg) orally, and rats in the HFD + V9 group were orally administered B. Lactis V9 (1 × 109 CFU/ml) (Yan et al. 2020). An equal volume of distilled water was given to the rats in other groups for a total of 4 weeks. The tests were carried out following the regulations for animal studies set by the Animal Ethics Committee at Inner Mongolia Agricultural University (No.[2020]084). Rats were put to death following an intraperitoneal injection of a 1% solution of pentobarbital sodium (0.17 mL per 100 g). Blood samples, cecum contents and liver tissue samples were collected and weighed. Liver samples were partially fixed in 4% paraformaldehyde overnight, while the rest of the tissues were promptly frozen in liquid nitrogen and kept at – 80 ℃ until additional analysis.
Biochemical analysis
Serum samples were centrifuged at 3000×g (HERMLE, Z326K, German) for 10 min and stored at – 80 °C for subsequent analysis. Serum levels of alanine aminotransferase (ALT), aminotransferase (AST), total cholesterol (TC), and glucose (GLU) were determined using an automated biochemistry analyzer (Olymbus 2700) according to the instructions of the clinical laboratory of Inner Mongolia People’s Hospital (Hohhot, China). Serum concentrations of IL-1β, IL-6, and TNF-α were determined by ELISA kits following the manufacturer’s instructions. Liver levels of SOD and MDA were measured by using commercial biochemical kits (Chen et al. 2024).
Histological analysis
After 24 h of fixation, liver tissues were dehydrated in ethanol treated by xylene, and then embedded in paraffin. The liver samples were sectioned at a thickness of 5 μm and subsequently stained with hematoxylin-eosin (H&E). The images of HE staining were obtained by using a light microscope (Nikon Eclipse, E100, Japan) equipped with a digital camera and were blindly evaluated by a pathologist (Azevedo Tosta et al. 2019).
Real-time PCR assay
Total RNA from liver tissue was extracted using Trizol reagent, and the concentration and purity of RNA (OD 260/280) were determined using a NanoDrop 2000 C spectrophotometer (Thermo Scientific, Waltham, USA). Total RNA was reversely transcribed to cDNA using the Prime Script RT reagent Kit with gDNA Eraser. Real-time PCR analysis was performed to determine the transcription levels of specific genes using the qTOWER system2.2 (Jena Analytical Instruments, Germany). Primer sequences used in this study are listed in Supplemental Table S1. he relative gene expression levels was calculated by the 2−ΔΔCT method (Kralik and Ricchi 2017).
Western blotting analysis
The hepatic protein lysates were obtained by homogenization in radioimmunoprecipitation assay (RIPA) buffer which is added with protease inhibitors. The protein concentrations were determined by using BCA protein assay kits. Equal amounts of lysates were separated by sodium dodecyl sulfate polyacrylamide (SDS-PAGE) gels and then transferred to polyvinylidene difluoride (PVDF) membranes (Invitrogen). After 1 h blocking in a solution of 5% (w/v) skim milk in tris-buffered saline (TBS) containing 0.1% Tween 20 (TBST), the membranes were immunoblotted with primary antibodies at 4℃ overnight. The membranes were then washed 5 times with TBST and incubated with secondary goat anti-rabbit IgG H&L (IRDye® 800CW, ab216775) at room temperature for 1 h. The images of protein expression were acquired by using the LI-COR Odyssey® Infrared Imaging System. Quantification of the density of the protein bands were performed by using the Image Studio (version 5.2) software (Taylor and Posch 2014).
16 S rRNA sequencing analysis of gut microbiota
To ensure the integrity of the DNA in the fecal samples, the fecal samples were placed in a DNA-protecting solution immediately after dissociation. Total DNA was then extracted according to the instructions of the E.Z.N.A.® soil kit (Omega Bio-tek, Norcross, GA, U.S.A). The purity, as well as the concentration of DNA was determined by using a NanoDrop2000. An Illumina-based high-throughput sequencing of the 16S rRNA genes V3-V4 regions approach was used to characterize the gut microbiota. The V3-V4 region of the bacterial 16S rRNA gene was amplified with primers as follow (343F 5’- TACGGRAGGCAGCAG-3’ and 798R 5’-AGGGTATCTAATCCT-3’). The sequencing results were clustered into operational taxonomic units (OTUs) with 3% difference (97% similarity). The Vsearch software (v.2.4.2) was used for subsequent bioinformatics analysis. Significant differences (P < 0.05) between groups were determined using linear discriminant analysis (LDA) effect size (LEfSe) (LDA > 2) (Pollock et al. 2018).
Statistical analysis
In this study, we used SPSS software (version 21.0) to conduct the Shapiro-Wilk test on the experimental data and found that P > 0.05, indicating that the data follows a normal distribution. The experimental data were analyzed by one-way analysis of variance (ANOVA) and Duncan’s multiple range test using SPSS software (version 21.0). P < 0.05 was considered as statistically significant difference. Data were expressed as mean ± standard deviation (SD). The gut microbiota data for this study was processed using the Wekemo Bioincloud (https://www.bioincloud.tech) (Gao et al. 2024). Abundance differences of gut microbiota at the phylum and genus levels were analyzed, identifying key gut microbiota between groups using LEfSe. Additionally, the relationship between different abundance OTUs and environmental factors was determined using Spearman correlation analysis.
Results
SP treatment attenuates high-fat diet-induced liver injury
In order to investigate the alleviating effect of high-fat diet (HFD)-induced liver injury by SP supplementation, we studied the changes in liver function and hepatic pathohistology. The serum levels of ALT and AST were significantly increased in the HFD group, which were 1.6-fold and 2.2-fold higher than those in the control group (P < 0.05). SP supplementation significantly reduced the HFD-induced increases in ALT and AST levels by 2.8-fold and 4.5-fold, respectively (P < 0.05, Fig. 1A–B, Table S2). The morphological changes of the liver tissue were observed by Hematoxylin and Eeosin (HE) staining. As shown in Fig. 1C, there existed disorganized hepatic cell cords and a marked accumulation of lipid droplets in the liver of the HFD group. In contrast, treatment with SP and B. Lactis V9 evidently reduced the hepatocellular accumulation of lipid droplets (P < 0.05, Fig. 1C–D). These results suggest that SP treatment attenuate HFD-induced hepatic steatosis and injury.

SP attenuates the extent of liver injury and hepatic histopathological lesions in NAFLD rats. A, B Serum expression levels of alanine aminotransferase (ALT) and glutamine aminotransferase (AST). C Representative hepatic tissue H&E staining images (200×). D Histologic scores (n = 3). Results were expressed as Mean ± SD. Significant differences between groups were expressed as P < 0.05
Effects of SP supplementation on lipid metabolism-related genes in NAFLD rats
To investigate the alleviating effect of SP on high-fat diet-induced disorder in lipid metabolism, the serum levels of TG and TC were measured. As shown in Fig. 2A and B, in agreement with the supplementation of B. Lactis V9, SP treatment significantly reduced the serum TG and TC levels (P < 0.05) In addition, SP supplementation also significantly reduced serum GLU levels (P < 0.05, Fig. 2C, Table S2). The hepatic expression levels of lipid-related genes were detected subsequently. The mRNA levels of ACC, FASN and PPARγ were significantly elevated by 1.6-fold (P < 0.05), 1.2-fold (P < 0.05) and 1.2-fold (P < 0.05) in rats of HFD group, respectively. In contrast, the mRNA levels of CPT1α and PPARα were significantly reduced by 2.3-fold (P < 0.05) and 1.2-fold (P < 0.05), respectively. This trend was reversed in those rats treated with SP as well as V9 (P < 0.05, Fig. 2D–H).

SP supplementation improves lipid metabolism in NAFLD rats. A Serum TG levels. B Serum TC levels. C Serum GLU levels. D–H mRNA levels of lipid metabolism-related genes in the liver. Results are expressed as mean ± SD. Group differences are indicated at P < 0.05
In addition, we also examined the protein expression levels of AMPK and PPARγ. As shown in Fig. 3, the protein expression of phosphorylated-AMPK (p-AMPK) was significantly reduced in the HFD group, and SP supplementation significantly elevated the expression levels of p-AMPK (P < 0.05). Meanwhile, SP treatment decreased HFD-induced PPARγ expression (P < 0.05). These results suggest that SP supplementation activates the AMPK/PPARγ signaling pathway in HFD-induced NAFLD rats, which in turn alleviates HFD-induced lipid metabolism disorders.

The impact of SP treatment on AMPK and PPARγ. Representative immunoblot images of phosphorylated AMPK (p-AMPK), total AMPK, and PPARγ. The results are presented as mean ± SD. Group differences are indicated by P < 0.05
SP treatment attenuates HFD-induced oxidative stress
In order to investigate the mitigating effect of SP on HFD-induced oxidative stress, the serum levels of SOD and MDA were measured. As shown in Fig. 4A and B, the SOD levels were decreased while MDA increased in the HFD group which is reversed by SP treatment (P < 0.05, Table S2). We went on to measure the expression levels of hepatic antioxidant-related genes by RT-PCR. A significant decrease in the mRNA expression of Nrf-2, HO-1, GCLC and SOD-2 were found in the HFD group (P < 0.05), and SP treatment significantly up-regulated the transcription of these genes (P < 0.05, Fig. 4C). The protein expression of Nrf-2, HO-1, SOD-2 and GCLC was quantified via Western blotting followed by densitometry analysis. As shown in Fig. 4D, the protein expression of Nrf-2, HO-1, SOD-2 and GCLC was significantly decreased in the HFD group (P < 0.05). The oral administration of SP as well as B. Lactis V9 reversed the suppression of these anti-oxidative protein levels (P < 0.05). These results suggest that SP treatment may attenuate HFD-induced oxidative stress by modulating the Nrf-2/HO-1 signaling pathway.

The effects of SP treatment on oxidative stress in NAFLD rats. A Serum MDA levels. B Serum SOD levels. C Involvement of liver genes Nrf-2, HO-1, SOD-2, and GCLC in antioxidant defense at the mRNA levels. D Representative western blot images of Nrf-2, HO-1, SOD-2 and GCLC. Results are expressed as mean ± SD. Inter-group differences are indicated by P < 0.05
SP supplementation suppresses pro-inflammatory gene expression in NAFLD rats
The effects of SP on HFD-induced inflammatory cytokine production were evaluated by ELISA. The results showed that the levels of TNF-α, IL-6 and IL-1β were significantly elevated in the HFD group (P < 0.05), while significantly reduced in the HFD + SP group (P < 0.05, Fig. 5A–C). It was also found that TNF-α, IL-6 and IL-1β mRNA levels in the HFD group were significantly higher than those in the Ctrl group (P < 0.05). SP treatment significantly down-regulated the mRNA levels of TNF-α, IL-6 and IL-1β (P < 0.05), which is in line with berberine hydrochloride and B. Lactis V9 supplementation (Fig. 5D–F). These results suggest that SP supplementation inhibits the expression of pro-inflammatory genes in NAFLD rats.

The effects of SP on the expression of pro-inflammatory genes in NAFLD rats were supplemented. A Serum TNF-α levels. B Serum IL-1β levels. C Serum IL-6 levels. D–F mRNA levels of TNF-α, IL-1β, and IL-6 in the liver. Results are presented as mean ± SD. Significant differences between groups are indicated by P < 0.05
SP supplementation modulates gut microbiota in NAFLD rats
The impact of SP on the intestinal microbiota of NAFLD rats were expored using 16 S ribosomal RNA (rRNA) gene sequencing. After sequence processing and filtering, a total of 648,013 high-quality sequences were obtained. We ultimately obtained 8,091 operational taxonomic units (OTUs) after clustering. As illustrated in the Venn diagram, there were 576 shared OTUs among the five groups, with 4, 5, 4, 18, and 12 uniquely identified OTUs in the Ctrl, HFD, HFD + BER, HFD + SP, and HFD + V9 groups, respectively (Fig. S1A). To investigate the specific changes in the intestinal microbiota due to SP treatment, the relative abundance of dominant taxa determined by sequencing was analyzed. The taxonomic analysis indicated that at the phylum level, bacterial abundance was primarily classified as Firmicutes, Bacteroidetes, Proteobacteria, Tenericutes, Cyanobacteria, Lentisphaerae, Verrucomicrobia, and Actinobacteria. Compared to the Ctrl group, the HFD group showed a significant decrease in the levels of Firmicutes and Tenericutes, and a significant increase in Cyanobacteria and Lentisphaerae. SP treatment resotored of the abundance of intestinal microbiota (Fig. 6A). Environmental factors have a significant impact on the host and intestinal microbiota homeostasis. Therefore, Spearman correlation coefficients were calculated to analyze whether environmental factors were associated with the intestinal microbiota. As shown in Fig. 6B, Lentisphaerae showed significant positive correlation (P < 0.05) with ALT, ACC and IL-6 (P < 0.01), and significant negative correlation (P < 0.05) with SOD. Firmicutes showed significant positive correlation (P < 0.05) with GCLC. Tenericutes showed significant positive correlation (P < 0.05) with CPT1α, PPARα and SOD-2 were significantly positively correlated (P < 0.05, P < 0.01).

The effects of SP supplementation on the gut microbiota of NAFLD rats were evaluated. A Proportion of bacterial gate composition in each group. B Correlation analysis between gates and NAFLD parameters. Red color indicates positive correlation and blue color indicates negative correlation. Significant differences are indicated by *P < 0.05, **P < 0.01, ***P < 0.001
Subsequently, the composition of the intestinal microbiota was further analyzed at the genus level. The addition of SP significantly increased the abundance of Acetatifactor sp., Corynebacterium sp., Facklamia sp., Peptococcus sp., and RC9_gut_group sp., while significantly reducing the abundance of Parabacteroides sp. (Fig. 7A, B). LEfSe analysis was performed to identify the major intestinal microbiota in different groups. The results revealed that there were many different genera in the intestinal microbiota of different treatment groups. In the Ctrl group, Anaerovorax sp. and Alistipes sp. had a relatively high abundance, while in the HFD group, Candidatus_Arthromitus sp. had a much higher relative abundance compared to other treatment groups. Additionally, it was found that following SP treatment, Peptococcus sp. and Acetatifactor sp. had a considerably higher relative abundance compared to other treatment groups, and following B. Lactis V9 treatment, RC9_gut_group sp., Lachnospiraceae sp., and Helicobacter sp. had higher relative abundances compared to other treatment groups. In the berberine supplement group, Subdoligranulum sp. and Parabacteroides sp. had higher relative abundances compared to other treatment groups (Fig. S1B). In addition, the Spearman correlation analysis showed that Peptococcus sp. and RC9_gut_group sp. were significantly negatively correlated with MDA (P < 0.05) and significantly positively correlated with CPT1α, PPARα, and SOD-2 (P < 0.05, P < 0.01); Parabacteroides sp. were significantly positively correlated with ALT, AST, MDA, and IL-1β (P < 0.05) and significantly negatively correlated with CPT1α, PPARα, SOD-2, and GCLC (P < 0.05, P < 0.01); Acetatifactor sp. and Corynebacterium sp. were significantly negatively correlated with IL-6 (P < 0.05) and significantly positively correlated with GCLC (P < 0.05); Facklamia sp. was significantly negatively correlated with IL-6 and ACC (P < 0.01) and significantly positively correlated with SOD (P < 0.05, Fig. 7C) These results indicate that SP can regulate the intestinal microbiota of NAFLD rats.

The supplementation of SP further affects the horizontal bacterial abundance in NAFLD rats. A Proportion of bacterial genus composition in each group. B Relative abundance of bacteria at the genus level in each subgroup analyzed by multiple significance (P < 0.05). C Correlation analysis between genera and NAFLD parameters. Red color indicates positive correlation and blue color indicates negative correlation. Significant differences are indicated by *P < 0.05, **P < 0.01
Discussion
NAFLD is a high prevalence disease characterized by excessive lipid accumulation or fat degeneration, with a global prevalence of up to 38%, and can progress from simple fat accumulation to cirrhosis and hepatocellular carcinoma (Vitulo et al. 2023; Younossi et al. 2023). A substantial body of research has demonstrated that polysaccharides, as natural products with wide pharmacological activities, can intervene in the occurrence and development of NAFLD by improving glucose and lipid metabolism, exerting antioxidant and anti-inflammatory effects, and regulating the gut microbiota (Hu et al. 2023). Our previous studies have shown that Seabuckthorn polysaccharides have a mitigating effect on carbon tetrachloride, LPS/D-galactosamine (d-GalN), and APAP -induced liver injury (Liu et al. 2015; Zhang et al. 2017; Wang et al. 2018). This study demonstrate that Seabuckthorn polysaccharides can alleviate HFD-induced NAFLD by regulating the AMPK/PPARγ, Nrf-2/HO-1 pathways, and the gut microbiota.
According to statistics, among blood donors with NAFLD, as many as 90% of cases showed elevated levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) (Clark et al. 2003). In this study, the levels of ALT and AST in the serum of HFD-induced NAFLD rats were significantly increased. Supplementation with SP, berberine hydrochloride, and B. Lactis V9 lowered the levels of serum ALT and AST. The liver plays a crucial role in the metabolism of lipids and lipoproteins, with abnormal hepatic lipid accumulation being a key feature of NAFLD (Geng et al. 2021). Therefore, improving liver lipid metabolism is a critical target for treating NAFLD. Previous studies have shown that plant polysaccharides can significantly reduce serum triglyceride (TG) and total cholesterol (TC) levels, alleviating hepatic steatosis in mice (Hao et al. 2024). The results of this study are consistent with this, and supplementation with SP also reduces serum glucose (GLU) levels.
AMP-activated protein kinase (AMPK) serves as a central regulator for multiple metabolic pathways. Activating AMPK can enhance glucose uptake, promote fatty acid oxidation, and simultaneously inhibit fatty acid and cholesterol synthesis. Phosphorylation of AMPK leads to the inactivation of acetyl-CoA carboxylase (ACC), resulting in decreased levels of malonyl-CoA, an inhibitor of carnitine palmitoyltransferase I (CPTI). Therefore, activating AMPK can upregulate CPTI to enhance fatty acid oxidation, thus improving NAFLD (McGarry and Foster 1980). Existing research indicates that plant extracts can alleviate HFD-induced NAFLD through the AMPK signaling pathway (Shen et al. 2023; Xu et al. 2024). Moreover, previous studies have shown that Astragalus polysaccharide (APS) can improve hepatic lipid accumulation in NAFLD rats by activating AMPK (Zhong et al. 2022). Our findings are consistent with the above results by finding that SP supplementation can activate AMPK, upregulate CPT1α, and downregulate genes related to fatty acid synthesis (ACC and FASN). Furthermore, we found that the increased expression of hepatic expression of PPARγ in NAFLD rats was inhibited after treatment with SP. This aligns with the discovery that obesity leads to increased hepatic transcription levels of PPARγ and that millet polyphenols mitigated NAFLD by inhibiting PPARγ (Pettinelli and Videla 2011; Cui et al. 2022). Therefore, we conclude that SP may alleviate lipid accumulation in NAFLD through the AMPK/PPARγ signaling pathway.
Increased lipid accumulation can lead to oxidative stress and lipid peroxidation, resulting in cellular damage and disrupted lipid metabolism.
(James et al. 1998; Morita et al. 2012). Oxidative stress in hepatocytes is caused by an excessive generation of reactive oxygen species (ROS) or a decrease in antioxidant defense capabilities. Superoxide dismutase (SOD) is part of the antioxidant enzyme system and serves to protect cells from ROS damage. Reduced SOD activity in chronic liver disease suggests that the defense mechanisms against oxidative attack are being compromised (Arroyave-Ospina et al. 2021). Malondialdehyde (MDA) serves as an important indicator of oxidative stress and a biomarker of lipid peroxidation. In line with these findings, it was observed that SOD activity decreased and MDA levels significantly increased in the serum of NAFLD rats, while treatment with SP resulted in a significant decrease in MDA activity and an increase in SOD levels. Nrf-2 plays a crucial role in alleviating oxidative stress and regulating lipid peroxidation. Nrf-2 activation induces the expression of fatty acid transport-related gene CPT1α, promotes fatty acid β-oxidation, lowers hepatic lipid content, and mitegates hepatic steatosis (Park et al. 2023). Studies indicate that active compounds from plants can protect against NAFLD by activating the Nrf-2 signaling pathway (Peng et al. 2024; Xue et al. 2024b). In this study, it was also found that SP can activate Nrf-2 expression in NAFLD rats. Additionally, dietary supplementation of SP can also activate the expression of HO-1 and SOD-2 in NAFLD rats, similar to the protective effect of Cudrania tricuspidata Extract via the Nrf-2/HO-1 pathway in NAFLD (Shrestha et al. 2021). Therefore, it can be concluded that SP may improve NAFLD by activating the Nrf-2/HO-1/SOD-2 signaling pathway.
The lipid peroxidation plays a key role in driving the pathogenesis of liver diseases, leading to inflammation and cell damage (Martín-Fernández et al. 2022). In chronic liver disease, Kupffer cells (KCs) play a crucial role in inflammation. During liver injury, KCs are activated and release a large number of inflammatory cytokines and chemokines (Chen et al. 2020). Furthermore, in KCs, Nrf-2 serves as a key regulator of the inflammatory response, maintaining cellular homeostasis and tissue integrity, thereby reducing liver inflammation (Xue et al. 2021). The results of this study indicate that the pro-inflammatory cytokines tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and IL-6 were significantly upregulated in HFD-induced NAFLD rats, but their expression levels were significantly downregulated after SP treatment. In addition, supplementing SP also reduced the transcription levels of TNF-α, IL-1β, and IL-6 in the NAFLD liver. These results are consistent with supplementation of berberine hydrochloride and B. Lactis V9. Therefore, we conclude that SP may improve high-fat diet-induced NAFLD by activating Nrf-2 to downregulate the levels of inflammatory factors.
Numerous studies have shown that the gut microbiota has become an important factor in the development of metabolic diseases, such as obesity, NAFLD, and T2DM (Aron-Wisnewsky et al. 2020; Wu et al. 2021b; Zeng et al. 2024). In recent years, researchers have found that traditional Chinese medicine can exert pharmacological effects by regulating the composition of the gut microbiota and affecting the metabolism of intestinal flora (Bao et al. 2022). Polysaccharides are one of the main active ingredients in Chinese herbal medicines and can improve NAFLD by regulating the gut microbiota (Hu et al. 2023). For example, modified apple polysaccharide can regulate the gut flora and reduce obesity in mice (Li et al. 2020). Polysaccharides from fermented Momordica charantia L. improved high-fat diet-induced obesity by increasing the phylum Firmicutes and decreasing the phylum Proteobacteria and genera Helicobacter sp. (Wen et al. 2021). Our previous studies have found that Astragalus polysaccharides can improve NAFLD by regulating the gut microbiota (Zhong et al. 2022). In addition to regulating the gut microbiota, polysaccharides also have pharmacological activities such as antioxidant, anti-inflammatory, regulation of glucose and lipid metabolism, and immunoregulation, which are related to prebiotic properties (Guo et al. 2023). In this study, SP supplementation increased the bacterial diversity of the gut microbiota in NAFLD rats. The content of Firmicutes and Tenericutes in the HFD group was significantly reduced, while the content of Cyanobacteria and Lentisphaerae was significantly increased, consistent with previous studies (Wu et al. 2021b; Longo et al. 2023; Dai et al. 2024). Furthermore, this study found that after SP treatment, the abundance of phylum Firmicutes, Tenericutes, Cyanobacteria, and Lentisphaerae all showed signs of recovery. It was also observed that Lentisphaerae was significantly positively correlated with ALT, ACC, and IL-6 (P < 0.05, P < 0.01), and significantly negatively correlated with SOD (P < 0.05).
In addition, this study also found that after SP supplementation, the abundance of Acetatifactor sp., Peptococcus sp., Corynebacterium sp., Facklamia sp., and RC9_gut_group sp. increased, while the abundance of Parabacteroides sp. decreased. Furthermore, the LEfSe analysis results showed that after SP treatment, the relative enrichment of Acetatifactor sp. and Peptococcus sp. abundance was much higher than that of other treatment groups (LDA > 2). Previous studies found that the progression of NAFLD is associated with the decrease in abundance of Peptococcus sp. and Corynebacterium sp. (Kordy et al. 2021; Zhang et al. 2023b). In the obesity model, Erchen decoction can down-regulate the relative abundance of Parabacteroides sp. (Zhang et al. 2023b). Our research results are consistent with this. SCFAs have anti-obesity and anti-inflammatory effects, and can prevent and improve the progression of NAFLD (Ohtani et al. 2023). Study found that Compound chenpi tea increased the abundance of Acetatifactor sp., improving the intestinal microbiota imbalance induced by HFD and thus improving diet-induced obesity (Wang et al. 2024). Acetatifactor sp. can metabolize to produce Short-chain fatty acids (SCFAs), affecting appetite and regulating lipid and glucose metabolism, thereby improving diet-induced obesity (Chambers et al. 2015). And study found that α-Lactalbumin peptide Asp-Gln-Trp can increase the relative abundance of RC9_gut_group sp. that produces SCFAs (Chen et al. 2022). RC9_gut_group sp. can effectively promote lipid metabolism (Jiang et al. 2021). In this study, the relative abundance of Acetatifactor sp. and RC9_gut_group sp. decreased in HFD rats, and their abundance recovered after SP treatment. However, whether SP regulates SCFAs requires further study. Furthermore, we further found that Peptococcus sp. and RC9_gut_group sp. are significantly negatively correlated with MDA (P < 0.05) and significantly positively correlated with CPT1α, PPARα, and SOD-2 (P < 0.05, P < 0.01); Parabacteroides sp. are significantly positively correlated with ALT, AST, MDA and IL-1β (P < 0.05), and significantly negatively correlated with CPT1α, PPARα, SOD-2, and GCLC (P < 0.05, P < 0.01); Acetatifactor sp. and Corynebacterium sp. are significantly negatively correlated with IL-6 (P < 0.05) and significantly positively correlated with GCLC (P < 0.05); Facklamia sp. is significantly negatively correlated with IL-6 and ACC (P < 0.01) and significantly positively correlated with SOD (P < 0.05). Therefore, we speculate that SP can improve NAFLD by regulating the intestinal microbiota. However, there are limitations to the contribution of microbiota changes to the effect of SP on improving NAFLD, and it is necessary to assess the regulatory role of the microbiota through studies on the consumption of microbiota by antibiotics or germ-free mice, as well as the role of gut microbiota metabolites.
In summary, SP can exert prebiotic properties to improve lipid deposition, oxidative stress, and inflammatory responses in NAFLD. This may be related to modulating the gut microbiota, regulating AMPK/PPARγ to improve lipid metabolism, and activating the Nrf-2/HO-1 signaling pathway. These findings suggest that SP as a prebiotic preparation may be a promising approach for preventing and improving NAFLD, providing a theoretical basis for subsequent research on the metabolic effects of SP. However, further investigation is needed regarding role of SP in the metabolism of gut microbiota in NAFLD.
Table S2 The impact of SP on the basic indicators of NAFLD. SD standard deviation, ALT Alanine transaminase, AST Aspartate aminotransferase, TG Triglyceride, TC Total cholesterol, MDA Malondialdehyde, SOD Superoxide dismutase. The difference between letters P < 0.05.
Data availability
The data used to support the findings of this study are available in NCBI-SRA under accession number PRJNA1113304 (https://www.ncbi.nlm.nih.gov/bioproject/PRJNA1113304). All data are available within the manuscript and supplementary materials. The raw data are available upon reasonable request from the corresponding authors.
Abbreviations
- NAFLD:
-
Non-alcoholic fatty liver disease
- SP:
-
Seabuckthorn polysaccharides
- V9:
-
B. Lactis V9
- HFD:
-
High-fat diets
- ALT:
-
Alanine transaminase
- AST:
-
Aspartate aminotransferase
- TG:
-
Triglyceride
- TC:
-
Total cholesterol
- HE:
-
Hematoxylin-eosin
- ELISA:
-
Enzyme-linked immunosorbent assay
- AMPK:
-
AMP-activated kinase
- PPARs:
-
Peroxisome proliferative-activated receptors
- Nrf-2:
-
Nuclear factor erythroid 2-related factor 2
- HO-1:
-
Heme oxygenase-1
- SOD-2:
-
Superoxide dismutase-2
- ACC:
-
Acetyl-CoA carboxylase
References
Ai G, Wu X, Dou Y, Huang R, Zhong L, Liu Y, Xian Y, Lin Z, Li Y, Su Z, Chen J, Qu C (2022) Oxyberberine, a novel HO-1 agonist, effectively ameliorates oxidative stress and inflammatory response in LPS/D-GalN induced acute liver injury mice via coactivating erythrocyte metabolism and Nrf2 signaling pathway. Food Chem Toxicol 166:113215. https://doi.org/10.1016/j.fct.2022.113215
Aron-Wisnewsky J, Vigliotti C, Witjes J, Le P, Holleboom AG, Verheij J, Nieuwdorp M, Clément K (2020) Gut microbiota and human NAFLD: disentangling microbial signatures from metabolic disorders. Nat Rev Gastroenterol Hepatol 17:279–297. https://doi.org/10.1038/s41575-020-0269-9
Arroyave-Ospina JC, Wu Z, Geng Y, Moshage H (2021) Role of oxidative stress in the pathogenesis of non-alcoholic fatty liver disease: implications for prevention and therapy. Antioxid. https://doi.org/10.3390/antiox10020174
Azevedo Tosta TA, de Faria PR, Neves LA, do Nascimento MZ (2019) Computational normalization of H&E-stained histological images: progress, challenges and future potential. Artif Intell Med 95:118–132. https://doi.org/10.1016/j.artmed.2018.10.004
Bao Y, Han X, Liu D, Tan Z, Deng Y (2022) Gut microbiota: the key to the treatment of metabolic syndrome in traditional Chinese medicine - a case study of diabetes and nonalcoholic fatty liver disease. Front Immunol 13:1072376. https://doi.org/10.3389/fimmu.2022.1072376
Cariello M, Piccinin E, Moschetta A (2021) Transcriptional regulation of metabolic pathways via lipid-sensing nuclear receptors PPARs, FXR, and LXR in NASH. Cell Mol Gastroenterol Hepatol 11:1519–1539. https://doi.org/10.1016/j.jcmgh.2021.01.012
Chambers ES, Morrison DJ, Frost G (2015) Control of appetite and energy intake by SCFA: what are the potential underlying mechanisms? Proc Nutr Soc 74:328–336. https://doi.org/10.1017/s0029665114001657
Chen J, Deng X, Liu Y, Tan Q, Huang G, Che Q, Guo J, Su Z (2020) Kupffer cells in non-alcoholic fatty liver disease: friend or foe? Int J Biol Sci 16:2367–2378. https://doi.org/10.7150/ijbs.47143
Chen H, Sun Y, Zhao H, Qi X, Cui H, Li Q, Ma Y (2022) α-Lactalbumin peptide asp-gln-trp alleviates hepatic insulin resistance and modulates gut microbiota dysbiosis in high-fat diet-induced NAFLD mice. Food Funct 13:9878–9892. https://doi.org/10.1039/d2fo01343f
Chen L, Lei Y, Lu C, Liu D, Ma W, Lu H, Wang Y (2024) Punicic acid ameliorates obesity-related hyperlipidemia and fatty liver in mice via regulation of intestinal flora and lipopolysaccharide-related signaling pathways. Food Funct 15:5012–5025. https://doi.org/10.1039/d4fo00502c
Clark JM, Brancati FL, Diehl AM (2003) The prevalence and etiology of elevated aminotransferase levels in the United States. Am J Gastroenterol 98:960–967. https://doi.org/10.1111/j.1572-0241.2003.07486.x
Cui K, Zhang L, La X, Wu H, Yang R, Li H, Li Z (2022) Ferulic acid and P-Coumaric acid synergistically attenuate non-alcoholic fatty liver disease through HDAC1/PPARG-mediated free fatty acid uptake. Int J Mol Sci. https://doi.org/10.3390/ijms232315297
Dai X, Jiang K, Ma X, Hu H, Mo X, Huang K, Jiang Q, Chen Y, Liu C (2024) Mendelian randomization suggests a causal relationship between gut microbiota and nonalcoholic fatty liver disease in humans. Med 103:e37478. https://doi.org/10.1097/md.0000000000037478
de Vries M, Westerink J, Kaasjager K, de Valk HW (2020) Prevalence of nonalcoholic fatty liver Disease (NAFLD) in patients with type 1 diabetes Mellitus: a systematic review and Meta-analysis. J Clin Endocrinol Metab. https://doi.org/10.1210/clinem/dgaa575
Ding Y, Yanagi K, Yang F, Callaway E, Cheng C, Hensel ME, Menon R, Alaniz RC, Lee K, Jayaraman A (2024) Oral supplementation of gut microbial metabolite indole-3-acetate alleviates diet-induced steatosis and inflammation in mice. Elife. https://doi.org/10.7554/eLife.87458
Fang C, Pan J, Qu N, Lei Y, Han J, Zhang J, Han D (2022) The AMPK pathway in fatty liver disease. Front Physiol 13:970292. https://doi.org/10.3389/fphys.2022.970292
Francque S, Szabo G, Abdelmalek MF, Byrne CD, Cusi K, Dufour JF, Roden M, Sacks F, Tacke F (2021) Nonalcoholic steatohepatitis: the role of peroxisome proliferator-activated receptors. Nat Rev Gastroenterol Hepatol 18:24–39. https://doi.org/10.1038/s41575-020-00366-5
Friedman SL, Neuschwander-Tetri BA, Rinella M, Sanyal AJ (2018) Mechanisms of NAFLD development and therapeutic strategies. Nat Med 24:908–922. https://doi.org/10.1038/s41591-018-0104-9
Gao Y, Zhang G, Jiang S, Liu YX (2024) Wekemo Bioincloud: a user-friendly platform for meta-omics data analyses. Imeta 3:e175. https://doi.org/10.1002/imt2.175
Geng Y, Faber KN, de Meijer VE, Blokzijl H, Moshage H (2021) How does hepatic lipid accumulation lead to lipotoxicity in non-alcoholic fatty liver disease? Hepatol Int 15:21–35. https://doi.org/10.1007/s12072-020-10121-2
Gong Z, Tas E, Yakar S, Muzumdar R (2017) Hepatic lipid metabolism and non-alcoholic fatty liver disease in aging. Mol Cell Endocrinol 455:115–130. https://doi.org/10.1016/j.mce.2016.12.022
Guo Q, Li Y, Dai X, Wang B, Zhang J, Cao H (2023) Polysaccharides: the potential Prebiotics for Metabolic Associated fatty liver Disease (MAFLD). Nutrients. https://doi.org/10.3390/nu15173722
Hao P, Yang X, Yin W, Wang X, Ling Y, Zhu M, Yu Y, Chen S, Yuan Y, Quan X, Xu Z, Zhang J, Zhao W, Zhang Y, Song C, Xu Q, Qin S, Wu Y, Shu X, Wei K (2024) A study on the treatment effects of Crataegus pinnatifida polysaccharide on non-alcoholic fatty liver in mice by modulating gut microbiota. Front Vet Sci 11:1383801. https://doi.org/10.3389/fvets.2024.1383801
Hu YL, Ma Q, Dong X, Kong Y, Cai J, Li J, Dong C (2023) Research progress on the therapeutic effects of polysaccharides on non-alcoholic fatty liver diseases. Front Nutr 10:1107551. https://doi.org/10.3389/fnut.2023.1107551
Huang Y, Cao J, Zhu M, Wang Z, Jin Z, Xiong Z (2024) Bacteroides fragilis aggravates high-fat diet-induced non-alcoholic fatty liver disease by regulating lipid metabolism and remodeling gut microbiota. Microbiol Spectr. https://doi.org/10.1128/spectrum.03393-23
Idalsoaga F, Kulkarni AV, Mousa OY, Arrese M, Arab JP (2020) Non-alcoholic fatty liver disease and alcohol-related liver disease: two intertwined entities. Front Med 7:448. https://doi.org/10.3389/fmed.2020.00448
James OF, Day CP (1998) Non-alcoholic steatohepatitis (NASH): a disease of emerging identity and importance. J Hepatol 29:495–501. https://doi.org/10.1016/s0168-8278(98)80073-1
Jiang F, Gao H, Qin W, Song P, Wang H, Zhang J, Liu D, Wang D, Zhang T (2021) Marked seasonal variation in structure and function of gut microbiota in forest and alpine musk deer. Front Microbiol 12:699797. https://doi.org/10.3389/fmicb.2021.699797
Koo SH (2013) Nonalcoholic fatty liver disease: molecular mechanisms for the hepatic steatosis. Clin Mol Hepatol 19:210–215. https://doi.org/10.3350/cmh.2013.19.3.210
Kordy K, Li F, Lee DJ, Kinchen JM, Jew MH, La Rocque ME, Zabih S, Saavedra M, Woodward C, Cunningham NJ, Tobin NH, Aldrovandi GM (2021) Metabolomic predictors of non-alcoholic steatohepatitis and advanced fibrosis in children. Front Microbiol 12:713234. https://doi.org/10.3389/fmicb.2021.713234
Kralik P, Ricchi M (2017) A basic guide to real time PCR in Microbial diagnostics: definitions, parameters, and everything. Front Microbiol 8:108. https://doi.org/10.3389/fmicb.2017.00108
Kwok R, Choi KC, Wong GL, Zhang Y, Chan HL, Luk AO, Shu SS, Chan AW, Yeung MW, Chan JC, Kong AP, Wong VW (2016) Screening diabetic patients for non-alcoholic fatty liver disease with controlled attenuation parameter and liver stiffness measurements: a prospective cohort study. Gut 65:1359–1368. https://doi.org/10.1136/gutjnl-2015-309265
Li Y, Xu W, Sun Y, Wang Y, Tang Y, Li Y, Gao X, Song C, Liu L, Mei Q (2020) Modified apple polysaccharide regulates microbial dysbiosis to suppress high-fat diet-induced obesity in C57BL/6J mice. Eur J Nutr 59:2025–2037. https://doi.org/10.1007/s00394-019-02051-z
Liu H, Zhang W, Dong S, Song L, Zhao S, Wu C, Wang X, Liu F, Xie J, Wang J, Wang Y (2015) Protective effects of sea buckthorn polysaccharide extracts against LPS/d-GalN-induced acute liver failure in mice via suppressing TLR4-NF-κB signaling. J Ethnopharmacol 176:69–78. https://doi.org/10.1016/j.jep.2015.10.029
Longo L, de Castro JM, Keingeski MB, Rampelotto PH, Stein DJ, Guerreiro GTS, de Souza VEG, Cerski CTS, Uribe-Cruz C, Torres ILS,Álvares, -da-Silva MR (2023) Nicotinamide riboside and dietary restriction effects on gut microbiota and liver inflammatory and morphologic markers in cafeteria diet-induced obesity in rats. Nutrition 110:112019. https://doi.org/10.1016/j.nut.2023.112019
Martín-Fernández M, Arroyo V, Carnicero C, Sigüenza R, Busta R, Mora N, Antolín B, Tamayo E, Aspichueta P, Carnicero-Frutos I, Gonzalo-Benito H, Aller R (2022) Role of oxidative stress and lipid peroxidation in the pathophysiology of NAFLD. Antioxid. https://doi.org/10.3390/antiox11112217
McGarry JD, Foster DW (1980) Regulation of hepatic fatty acid oxidation and ketone body production. Annu Rev Biochem 49:395–420. https://doi.org/10.1146/annurev.bi.49.070180.002143
Morita M, Ishida N, Uchiyama K, Yamaguchi K, Itoh Y, Shichiri M, Yoshida Y, Hagihara Y, Naito Y, Yoshikawa T, Niki E (2012) Fatty liver induced by free radicals and lipid peroxidation. Free Radic Res 46:758–765. https://doi.org/10.3109/10715762.2012.677840
Ngo V, Duennwald ML (2022) Nrf2 and oxidative stress: a general overview of mechanisms and implications in human disease. Antioxid. https://doi.org/10.3390/antiox11122345
Ohtani N, Kamiya T, Kawada N (2023) Recent updates on the role of the gut-liver axis in the pathogenesis of NAFLD/NASH, HCC, and beyond. Hepatol Commun. https://doi.org/10.1097/hc9.0000000000000241
Park JS, Rustamov N, Roh YS (2023) The roles of NFR2-Regulated oxidative stress and mitochondrial quality control in chronic liver diseases. Antioxid. https://doi.org/10.3390/antiox12111928
Peng W, He CX, Li RL, Qian D, Wang LY, Chen WW, Zhang Q, Wu CJ (2024) Zanthoxylum Bungeanum amides ameliorates nonalcoholic fatty liver via regulating gut microbiota and activating AMPK/Nrf2 signaling. J Ethnopharmacol 318:116848. https://doi.org/10.1016/j.jep.2023.116848
Pettinelli P, Videla LA (2011) Up-regulation of PPAR-gamma mRNA expression in the liver of obese patients: an additional reinforcing lipogenic mechanism to SREBP-1c induction. J Clin Endocrinol Metab 96:1424–1430. https://doi.org/10.1210/jc.2010-2129
Pollock J, Glendinning L, Wisedchanwet T, Watson M (2018) The madness of microbiome: attempting to find consensus best practice for 16S microbiome studies. Appl Environ Microbiol. https://doi.org/10.1128/aem.02627-17
Puengel T, Liu H, Guillot A, Heymann F, Tacke F, Peiseler M (2022) Nuclear receptors linking metabolism, inflammation, and fibrosis in nonalcoholic fatty liver disease. Int J Mol Sci. https://doi.org/10.3390/ijms23052668
Shaw P, Chattopadhyay A (2020) Nrf2-ARE signaling in cellular protection: mechanism of action and the regulatory mechanisms. J Cell Physiol 235:3119–3130. https://doi.org/10.1002/jcp.29219
Shen Q, Chen Y, Shi J, Pei C, Chen S, Huang S, Li W, Shi X, Liang J, Hou S (2023) Asperuloside alleviates lipid accumulation and inflammation in HFD-induced NAFLD via AMPK signaling pathway and NLRP3 inflammasome. Eur J Pharmacol 942:175504. https://doi.org/10.1016/j.ejphar.2023.175504
Shrestha J, Baek DJ, Oh YS, Cho SS, Ki SH, Park EY (2021) Protective effect of Cudrania tricuspidata extract against high-fat diet induced nonalcoholic fatty liver disease through Nrf-2/HO-1 pathway. Molecules. https://doi.org/10.3390/molecules26092434
Smith BK, Marcinko K, Desjardins EM, Lally JS, Ford RJ, Steinberg GR (2016) Treatment of nonalcoholic fatty liver disease: role of AMPK. Am J Physiol Endocrinol Metab 311:E730–e740. https://doi.org/10.1152/ajpendo.00225.2016
Sun Z, Chen X, Wang J, Gao P, Zhou Z, Ren Y, Sun T, Wang L, Meng H, Chen W, Zhang H (2010) Complete genome sequence of probiotic Bifidobacterium animalis subsp. lactis strain V9. J Bacteriol 192:4080–4081. https://doi.org/10.1128/jb.00369-10
Taylor SC, Posch A (2014) The design of a quantitative Western blot experiment. Biomed Res Int 2014:361590. https://doi.org/10.1155/2014/361590
Vitulo M, Gnodi E, Rosini G, Meneveri R, Giovannoni R, Barisani D (2023) Current therapeutical approaches targeting lipid metabolism in NAFLD. Int J Mol Sci. https://doi.org/10.3390/ijms241612748
Wang X, Liu J, Zhang X, Zhao S, Zou K, Xie J, Wang X, Liu C, Wang J, Wang Y (2018) Seabuckthorn berry polysaccharide extracts protect against acetaminophen induced hepatotoxicity in mice via activating the Nrf-2/HO-1-SOD-2 signaling pathway. Phytomedicine 38:90–97. https://doi.org/10.1016/j.phymed.2017.11.007
Wang J, Hao J, Miao D, Xiao P, Jiang X, L EH (2024) Compound chenpi tea consumption reduces obesity-related metabolic disorders by modulating gut microbiota and serum metabolites in mice. J Sci Food Agric 104:431–442. https://doi.org/10.1002/jsfa.12940
Wayal V, Hsieh CC (2023) Bioactive dipeptides mitigate high-fat and high-fructose corn syrup diet-induced metabolic-associated fatty liver disease via upregulation of Nrf2/HO-1 expressions in C57BL/6J mice. Biomed Pharmacother 168:115724. https://doi.org/10.1016/j.biopha.2023.115724
Wen JJ, Li MZ, Gao H, Hu JL, Nie QX, Chen HH, Zhang YL, Xie MY, Nie SP (2021) Polysaccharides from fermented Momordica charantia L. with Lactobacillus plantarum NCU116 ameliorate metabolic disorders and gut microbiota change in obese rats. Food Funct 12:2617–2630. https://doi.org/10.1039/d0fo02600j
Wu J, Wang K, Wang X, Pang Y, Jiang C (2021) The role of the gut microbiome and its metabolites in metabolic diseases. Protein Cell 12:360–373. https://doi.org/10.1007/s13238-020-00814-7
Wu N, Zhou J, Mo H, Mu Q, Su H, Li M, Yu Y, Liu A, Zhang Q, Xu J, Yu W, Liu P, Liu G (2021b) The Gut Microbial Signature of Gestational Diabetes Mellitus and the Association with Diet intervention. Front Cell Infect Microbiol 11:800865. https://doi.org/10.3389/fcimb.2021.800865
Xiao J, Xiang H, Xiang H, Sun Z, Xu J, Ren H, Hu P, Peng M (2023) GW9662 ameliorates nonalcoholic steatohepatitis by inhibiting the PPARγ/CD36 pathway and altering the gut microbiota. Eur J Pharmacol 960:176113. https://doi.org/10.1016/j.ejphar.2023.176113
Xu J, Jia W, Zhang G, Liu L, Wang L, Wu D, Tao J, Yue H, Zhang D, Zhao X (2024) Extract of Silphium perfoliatum L. improve lipid accumulation in NAFLD mice by regulating AMPK/FXR signaling pathway. J Ethnopharmacol. https://doi.org/10.1016/j.jep.2024.118054
Xue R, Qiu J, Wei S, Liu M, Wang Q, Wang P, Sha B, Wang H, Shi Y, Zhou J, Rao J, Lu L (2021) Lycopene alleviates hepatic ischemia reperfusion injury via the Nrf2/HO-1 pathway mediated NLRP3 inflammasome inhibition in Kupffer cells. Ann Transl Med 9:631. https://doi.org/10.21037/atm-20-7084
Xue Y, Peng Y, Zhang L, Ba Y, Jin G, Liu G (2024) Effect of different exercise modalities on nonalcoholic fatty liver disease: a systematic review and network meta-analysis. Sci Rep 14:6212. https://doi.org/10.1038/s41598-024-51470-4
Xue Y, Wei Y, Cao L, Shi M, Sheng J, Xiao Q, Cheng Z, Luo T, Jiao Q, Wu A, Chen C, Zhong L, Zhang C (2024b) Protective effects of scutellaria-coptis herb couple against non-alcoholic steatohepatitis via activating NRF2 and FXR pathways in vivo and in vitro. J Ethnopharmacol 318:116933. https://doi.org/10.1016/j.jep.2023.116933
Yan Y, Liu C, Zhao S, Wang X, Wang J, Zhang H, Wang Y, Zhao G (2020) Probiotic Bifidobacterium lactis V9 attenuates hepatic steatosis and inflammation in rats with non-alcoholic fatty liver disease. AMB Express 10:101. https://doi.org/10.1186/s13568-020-01038-y
Younossi ZM, Golabi P, Paik JM, Henry A, Van Dongen C, Henry L (2023) The global epidemiology of nonalcoholic fatty liver disease (NAFLD) and nonalcoholic steatohepatitis (NASH): a systematic review. Hepatology 77:1335–1347. https://doi.org/10.1097/hep.0000000000000004
Yuandangongbu YT (1983) Si Bu Yi Dian. The Publishing Company of Public Health, Beijing
Zeng H, Liu C, Wan L, Peng L, Wen S, Fang W, Chen H, Wang K, Yang X, Huang J, Liu Z (2024) (-)-Epicatechin ameliorates type 2 diabetes mellitus by reshaping the gut microbiota and gut-liver axis in GK rats. Food Chem 447:138916. https://doi.org/10.1016/j.foodchem.2024.138916
Zhang W, Zhang X, Zou K, Xie J, Zhao S, Liu J, Liu H, Wang J, Wang Y (2017) Seabuckthorn berry polysaccharide protects against carbon tetrachloride-induced hepatotoxicity in mice via anti-oxidative and anti-inflammatory activities. Food Funct 8:3130–3138. https://doi.org/10.1039/c7fo00399d
Zhang L, Chen N, Zhan L, Bi T, Zhou W, Zhang L, Zhu L (2023) Erchen Decoction alleviates obesity-related hepatic steatosis via modulating gut microbiota-drived butyric acid contents and promoting fatty acid β-oxidation. J Ethnopharmacol 317:116811. https://doi.org/10.1016/j.jep.2023.116811
Zhang L, Zi L, Kuang T, Wang K, Qiu Z, Wu Z, Liu L, Liu R, Wang P, Wang W (2023b) Investigating causal associations among gut microbiota, metabolites, and liver diseases: a mendelian randomization study. Front Endocrinol (Lausanne) 14:1159148. https://doi.org/10.3389/fendo.2023.1159148
Zhong M, Yan Y, Yuan H, A R, Xu G, Cai F, Yang Y, Wang Y, Zhang W (2022) Astragalus mongholicus polysaccharides ameliorate hepatic lipid accumulation and inflammation as well as modulate gut microbiota in NAFLD rats. Food Funct 13:7287–7301. https://doi.org/10.1039/d2fo01009g
Zou J, Song Q, Shaw PC, Zuo Z (2024) Dendrobium officinale regulate lipid metabolism in diabetic mouse liver via PPAR-RXR signaling pathway: evidence from an integrated multi-omics analysis. Biomed Pharmacother 173:116395. https://doi.org/10.1016/j.biopha.2024.116395
Acknowledgements
We appreciate the assistance provided by Yuning Yang, Rui Li and Zhiqiang Miao in the animal experiments. We thank Abdul Jamil Khan for proof-reading the manuscript.
Funding
This study was supported by the Basic Research Operating Expenses of Colleges and Universities directly under the Inner Mongolia Autonomous Region (BR220117), the Key Research and Development and Achievement Transformation Programme of Inner Mongolia Autonomous Region (2023YFDZ0029), and supported by Program for Innovative Research Team in Universities of Inner Mongolia Autonomous Region (NMGIRT2219).
Author information
Authors and Affiliations
Contributions
YW and JL conceived and planned the experiments, data analysis, and manuscript revision; YY was responsible for most of the experiments, data mobiles, and manuscript writing; HY, FY, and XY were responsible for the Real-time PCR and ELISA assays, as well as HE staining and pathological analyses; and HN was involved in the writing of the manuscript. All authors provided critical feedback and assistance with the study, analyses, and development of the manuscript. All authors have read and approved the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This article does not contain any studies with human participants performed by any of the authors. All experiments were approved by the Animal Care and Use Committee of Inner Mongolia Agricultural University (China) according to the Chinese Council on Animal Care guidelines. Ethics committee’s reference number: [2020]084.
Consent for publication
All authors listed have read the complete manuscript and have approved submission of the paper.
Competing interests
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
13568_2024_1756_MOESM1_ESM.tif
Fig. S1 (A) Venn diagram of the number of OTUs in each group. (B) Proportion of bacterial genus composition in each group. Supplementary Material 1.
13568_2024_1756_MOESM2_ESM.xls
Table S2 The impact of SP on the basic indicators of NAFLD. SD standard deviation, ALT Alanine transaminase, AST Aspartate aminotransferase, TG Triglyceride, TC Total cholesterol, MDA Malondialdehyde, SOD Superoxide dismutase. The difference between letters P<0.05. Supplementary Material 2.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Yan, Y., Yuan, H., Yang, F. et al. Seabuckthorn polysaccharides mitigate hepatic steatosis by modulating the Nrf-2/HO-1 pathway and gut microbiota. AMB Expr 14, 100 (2024). https://doi.org/10.1186/s13568-024-01756-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13568-024-01756-7