
Abstract
The summer (July–August) zooplankton of the Kama and Votkinsk reservoirs on the Kama River was studied for six years (2014–2019). A total of 106 crustacean and rotifer species were recorded; ten of them are invaders mainly of southern origin. It has been revealed that 70% of invaders in both reservoirs were recorded from 2012 to 2016. The cladoceran Diaphanosoma orghidani and Ponto-Caspian copepods Eurytemora caspica and Heterocope caspia have become common species; they annually occur in over 30% of samples and locally form up to 30% of the number of crustaceans. An increase in the density (up to 24–30% of the abundance of crustaceans) of the thermophilic copepod Thermocyclops crassus, which was previously not abundant, has been revealed. The average abundance of zooplankton for 2014–2019 (>150 thousand ind./m3) was generally 1.7–2.0 times higher in the littoral zone than in the pelagic zone in both reservoirs. The summer biomass of zooplankton was high (1.1–1.7 g/m3) throughout the water area of the reservoirs. The biomass level was 2.5–3 times higher in 2014–2019 than in the 1950–1960s.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
INTRODUCTION
The Kama River is the largest (>2000 km) and most high-water tributary of the Volga River; the average long-term water discharge in its lower reaches is over 4000 m3/s (Volga …, 1978). The cascade of Kama reservoirs was created in 1954–1979; the regulated part of the river is below the mouth of the Vishera River and includes three reservoirs: Kama, Votkinsk, and Nizhnekamsk (Edel’shtein, 1998). The trophic status of all Kama reservoirs in terms of chlorophyll a concentration in plankton is determined as eutrophic; in terms of the amount of chlorophyll and pheopigments in bottom sediments, the Kama Reservoir is eutrophic, while the Votkinsk Reservoir is mesotrophic (Belyaeva et al., 2018).
The zooplankton of the Kama and Votkinsk reservoirs has been studied since 1956 and 1965, respectively (Ulomskii, 1961; Udalova, 1968; Serkina, 1971, 1975; Poskryakova, 1977; Kortunova and Zueva, 1979; Kortunova and Serkina, 1980; Kortunova, 1983; Kortunova and Galanova, 1988; Kostitsyn et al., 2011; Krainev and Kuznetsova, 2013; Presnova and Khulapova, 2015; Seletkova, 2015). The state of the community is monitored by the Perm Branch of the Russian Federal Research Institute of Fisheries and Oceanography (PermNIRO). It is shown that the abundance of zooplankton in the Kama River is characterized by increasingly large fluctuations year after year (Kortunova, 1983; Kortunova and Galanova, 1988; Seletkova, 2015).
In 2016, a large group of invasive species, including brackish-water Ponto-Caspian crustaceans, was identified in the Kama reservoirs (Lazareva, 2020). The features of the structure and dynamics of the community abundance, taking into account the influence of invaders, have not been discussed in recent years.
The purpose of this research was to analyze the structure and dynamics of the abundance and biomass of summer zooplankton in the Kama and Votkinsk reservoirs for the period of 2014–2019 and determine the features of invader colonization.
MATERIALS AND METHODS
Pelagic and littoral zooplankton was collected at a depth of 5–13 and 1–3 m, respectively, in three parts of both reservoirs (upper (I), central (II), and near-dam (III) parts) in July–August in 2014–2019 according to the standard scheme of sections (stations) and sampling points of the Perm Branch of the Russian Federal Research Institute of Fisheries and Oceanography (Fig. 1). Eighteen sections were established in the two reservoirs. Crustaceans and rotifers were recorded in total zooplankton samples, which were collected using a Juday net with a mouth diameter of 12 cm and a mesh sieve of 100 µm. Samples were fixed with 4% formalin and examined at the laboratory under LOMO MSP-2 and BIOLAM 70 microscopes (LOMO-Micro, St. Petersburg).

Scheme and zoning of the Kama and Votkinsk reservoirs. Reservoir parts: (I) upper part, (II) central part, and (III) near-dam part.
The abundance and biomass of each identified species from the samples were determined and the total values of these parameters were calculated for four large taxonomic groups (Cladocera, Cyclopoida, Calanoida, and Rotifera) and the whole community. The dominants included species with an abundance of over 10% of the total number of crustaceans or rotifers.
The similarity of the structure of the dominant complexes was determined by the Czekanowski–Sorensen index for quantitative data (ICzS) (Pesenko, 1982):
ICzS = Σmin pi,
where pi is the proportion of the species in the total zooplankton abundance.
Changes in the zooplankton biomass in the Kama and Votkinsk reservoirs over the period from the beginning of the research to 2019 (the whole period of the existence of the water bodies) were analyzed using the literature sources and archival materials from the Russian Federal Research Institute of Fisheries and Oceanography, Perm Branch.
The average number of species in the sample, their abundance and biomass, and error of the mean were calculated.
RESULTS
Species richness and dominants. In 2014–2019, 100 species were identified in the Kama Reservoir and 69 species were found in the Votkinsk Reservoir (106 species in both reservoirs). Most of them (40–45%) were represented by Rotifera; the proportion of Cladocera was 35–40% and that of Copepoda was less than 20%. The species density (number of species in the sample) was high in all observation years. On average, 21 ± 3 species were found in samples from the pelagic zone of the Kama Reservoir and 18 ± 3 species were recorded in samples from the Votkinsk Reservoir; there were slightly fewer species in the coastal areas of both reservoirs (17 ± 2 species from each reservoir). Seven rotifer species, Keratella serrulata (Ehrenberg, 1838); Notholca acuminata (Ehrenberg, 1832); Euchlanis lyra Hudson, 1886; Mytilina mucronata (O.F. Müller, 1773); Synchaeta oblonga Ehrenberg, 1831; S. stylata Wierzejski, 1893; and Filinia brachiata (Rousselet, 1901) and two crustacean species, Alonella nana (Baird, 1843) and Ilyocryptus agilis Kurz, 1878, which were not indicated in the review list of (Lazareva, 2020), were identified in 2012 to 2014. Most of these species were found in the Kama Reservoir. Four species, E. lyra, Notholca acuminata, Synchaeta oblonga, and Alonella nana, were recorded in the Votkinsk Reservoir.
The composition of abundant rotifer species in the upper part of the Kama Reservoir significantly differed from their composition in its other two parts (Fig. 2a). It was often dominated here by Asplanchna priodonta Gosse, 1850 (10–60% Nrot); Brachionus angularis Gosse, 1851 (25–50% Nrot); and Synchaeta pectinata Ehrenberg, 1832 (30–40% Nrot). In the central and near-dam parts, the main contribution to the Nrot value was made by Kellicottia longispina (Kellicott, 1879) (20–90%); Euchlanis dilatata lucksiana (Hauer, 1939) (10–90%); Keratella quadrata (O.F. Müller, 1786) (10–20%); and Polyarthra major Bruckhardt, 1900 (10–20%).

Ratio (%) of the number of main dominants in the groups of rotifers (a, c) and crustaceans (b, d) in the pelagic (P) and littoral (L) zones of the Kama (a, b) and Votkinsk (c, d) reservoirs in 2014–2019 (average for 6 years). Reservoir parts: (I) upper part, (II) central part, and (III) near-dam part. Rotifera: (1) Euchlanis dilatata lucksiana, (2) Polyarthra major, (3) Keratella quadrata, (4) Kellicottia longispina, (5) Synchaeta pectinata, (6) Brachionus angularis, and (7) Asplanchna priodonta. Crustacea: (8) Thermocyclops crassus, (9) Mesocyclops leuckarti, (10) Bosmina longirostris, (11) Daphnia galeata, and (12) Bosmina cf. crassicornis.
The abundance of crustaceans throughout the water area of the reservoir was formed mainly by three species (Fig. 2b). These are the copepods Mesocyclops leuckarti (Claus, 1857) (10–70% Ncr) and Thermocyclops crassus (Fischer, 1853) (10–64% Ncr) and cladoceran Daphnia galeata Sars, 1864 (10–60% Ncr). In the littoral zone of the upper part, 10–23% of Ncr was formed by Bosmina (s. str) longirostris (O.F. Müller, 1785). On the whole, the pelagic and coastal communities in each part of the reservoir were characterized by a similar ratio of main dominants (Figs. 2a, 2b).
Bzoo was formed mainly by a large (body length up to 2.0 mm) cladoceran, Daphnia galeata, throughout the water area of the reservoir. Its contribution to the biomass varied from 10 to 90% during the 6 years of observations (on average, 55 ± 15% in the pelagic zone and 40 ± 10% in the littoral zone) (Fig. 3a). Rotifers of the genus Asplanchna significantly contributed in the upper part of the reservoir (on average up to 20% Bzoo and up to 50% Bzoo in some years).

Ratio (%) of the biomass of the main dominants of zooplankton in the pelagic (P) and littoral (L) zones of the Kama (a) and Votkinsk (b) reservoirs in 2014–2019 (average for the six years). Reservoir parts: (I) upper part, (II) central part, and (III) near-dam part. (1) Thermocyclops crassus, (2) Mesocyclops leuckarti, (3) Diaphanosoma orghidani, (4) Daphnia galeata, (5) Asplanchna herricki, (6) A. priodonta, (7) Heterocope caspia, and (8) Eurytemora caspica.
The structure of the dominant complexes of zooplankton in different parts of the reservoir was characterized by a relatively low level of similarity (generally <50%). This is due to “outbreaks” of the abundance of species (up to 15–40% Nrot and 10–30% Ncr) in some years which were not numerous in other years (e.g., Keratella cochlearis (Gosse, 1851), Ploesoma truncatum (Levander, 1894), Chydorus sphaericus (O.F. Müller, 1785), and Bosmina (Eubosmina) cf. crassicornis (Lilljeborg, 1887)) or are untypical for the biotope (Brachionus quadridentatus Hermann, 1783 in the pelagic zone and Eurytemora caspica Sukhikh et Alekseev, 2013 in the littoral zone). The highest values of the index of similarity in the community structure were recorded between the central and near-dam parts; they reached 60–70% in the pelagic zone and 65–75% in the littoral zone. The similarity in the zooplankton structure did not exceed 60% in the pelagic zone and 40% in the littoral zone over a number of years (from 2014 to 2019). It was the highest (50–60%) between years (2014 and 2019 and 2015, 2017, and 2019) with low summer heating of water (17 ± 2°C), while the maximum differences (similarity <35%) were observed between 2016 and 2018, when the water heating was high (22 ± 2°C).
Rotifers Euchlanis dilatata lucksiana and Kellicottia longispina were almost annually abundant throughout the pelagic zone of the Votkinsk Reservoir (15–95 and 10–80% Nrot, respectively) (Fig. 2c). Crustaceans were dominated by the copepods Mesocyclops leuckarti (30–80% Ncr) and Thermocyclops crassus (10–35% Ncr), which together formed up to 75% Ncr (Fig. 2d). In some years (2014–2015), Daphnia galeata was a common species (25–26%).
The taxocene of crustaceans in the shore zone of this reservoir actually did not differ from that in the pelagic one (Fig. 2d). Along with common rotifer species, Polyarthra major and Synchaeta pectinata were relatively abundant species (up to 35 and 30% Nrot, respectively) throughout the water area in the littoral zone of the central part (Fig. 2c).
Almost the entire biomass of zooplankton (up to 85%) throughout the water area of the reservoir was formed by the cladoceran Daphnia galeata (Fig. 3b). In the pelagic zone, 40% Bzoo was formed by Mesocyclops leuckarti. On the whole, the ratio of the biomass of the main dominants did not significantly differ between the pelagic zone and shore zone of the reservoir.
The structure of zooplankton was relatively homogeneous and characterized by a high level of similarity (>50%) in different parts of the reservoir. The highest values (60–70%) of the similarity index were recorded between the complexes of dominants in the central and near-dam parts. In some years (2018 and 2019), the similarity of the zooplankton structure reached 80% in the pelagic zone, while littoral communities differed very markedly during all six observation years (the similarity was less than 45%). As in the Kama Reservoir, a mass development (60–70% Nrot and 13–60% Ncr) of usually small-numbered species (e.g., Brachionus angularis, Asplanchna priodonta, Bosmina longirostris, Eurytemora caspica, and Heterocope caspia Sars, 1897) were locally recorded here in some years. The similarity of the structure of zooplankton in the Votkinsk Reservoir did not exceed 60% year by year; it was closest to 60% (54–60%) in 2014, 2015, 2016, and 2018, while the maximum differences (the similarity was less than 35%) were recorded between 2014 and 2019 and 2016 and 2019. In the intracascade Votkinsk Reservoir, no clear correlation was observed between the community structure and thermal water regime.
Role of invaders in the community. Nine to ten invasive species were recorded in the zooplankton of the studied reservoirs; they were identified here for the first time mainly in the 2010s (Table 1). Most of them (except Kellicottia bostoniensis (Rousselet, 1908)) are represented by southern thermophilic species. Three of them (the cladoceran Cercopagis pengoi (Ostroumov, 1891) and copepods Heterocope caspia and Eurytemora caspica) are brackish-water Ponto-Caspian forms. Three of the ten invaders—the cladoceran Diaphanosoma orghidani Negrea, 1982 and copepods Eurytemora caspica and Heterocope caspia—have become common in both reservoirs (they annually occur in more than 30% of samples).
The maximum abundance of most southern rotifers did not exceed 10 thousand ind./m3; only some of the species were locally dominant in the zooplankton of the reservoirs in some years. Thus, the abundance of Conochiloides coenobasis Skorikov, 1914 reached 20 thousand ind./m3 (22% Nrot) and that of Pompholyx sulcata Hudson, 1885 was 12 thousand ind./m3 (24% Nrot) in the upper part of the Kama Reservoir in August 2016. The abundance of Asplanchna henrietta Langhaus, 1906 was 14 thousand ind./m3 (11% Nrot) in the central part of the Votkinsk Reservoir near the mouth of the Ocher River in 2016.
A significantly greater contribution to the Ncr value was made by southern crustaceans; among them, large species also contributed greatly to Bzoo. The copepod Eurytemora caspica in the shore zone was highly abundant (up to 16 thousand ind./m3) in the central and near-dam parts of the Kama Reservoir in 2015 (25–30% Ncr) and throughout the water area of the Votkinsk Reservoir in 2019 (13–17% Ncr). The contribution of this species to Bzoo reached 25–40% in the littoral zone of the Kama Reservoir and 10–23% in the littoral zone of the Votkinsk Reservoir. The proportion of E. caspica was 14–16% Bzoo in the pelagic zone of the Votkinsk Reservoir.
Two other species made a significant contribution only to Bzoo. In 2016 and 2018, the abundance of Diaphanosoma orghidani reached 20–30 thousand ind./m3 in the upper part of the Kama Reservoir, or 13–17% Bzoo in the pelagic zone and 15–18% in the littoral zone. In the central part of the reservoir, the species was dominant only in the littoral zone (up to 14% Bzoo in 2019). In 2019, the copepod Heterocope caspia formed 12–40% Bzoo in the littoral zone and up to 55% Bzoo in the pelagic zone of the near-dam part in the Votkinsk Reservoir.
Abundance and biomass. In the pelagic zone of the Kama Reservoir, the total abundance of zooplankton varied from 30 to 190 (on average, 110 ± 27) thousand ind./m3 and 0.4 to 1.9 (on average, 1.2 ± 0.2) g/m3 in the summer of 2014 to 2019 (Table 2). The greatest contribution to the community abundance (>50%) was made by Cyclopoida (>20%) and Rotifera (30%); the biomass was formed mainly by Cladocera (>70%). In the central and near-dam parts, the ratio of the main taxa of zooplankton was stable for all 6 years of observations, while it varied greatly in the upper part from year to year (Fig. 4). The maximum (up to 60%) contribution of rotifers to the abundance of the community was observed here. The amount of meroplankton, represented by veligers of mollusks of the genus Dreissena, was very low in all years (on average, 1.0 ± 0.5 thousand ind./m3) at a biomass of less than 0.01 g/m3.

Ratio (%) of large taxonomic groups of zooplankton in different parts (I, upper part; II, central part; and III, near-dam part) of the Kama and Votkinsk reservoirs in 2014–2019.
The seasonal variation of the development of pelagic zooplankton was studied in the Kama Reservoir in 2016; it was characterized by an increase in abundance from spring (May) to summer (August) and a slight decrease in autumn (September–October) (Fig. 5a). In the central part of the reservoir, the number of zooplankton even slightly increased in autumn. On the contrary, the seasonal dynamics of the biomass was characterized by a clearly pronounced summer maximum typical for all parts of the reservoir (Fig. 5b). The biomass was minimal in spring and maximal in autumn in the upper part of the reservoir when compared to other parts.

Seasonal variation in the abundance (a) and biomass (b) of zooplankton in the Kama Reservoir in 2016. (1) Upper part (I), (2) central part (II), and (3) near-dam part (III).
In the pelagic zone of the Votkinsk Reservoir, the abundance of zooplankton varied from 13 to 107 (on average, 76 ± 15) thousand individuals/m3) and biomass was 0.1–1.4 (on average, 1.0 ± 0.2) g/m3) in summer from 2014 to 2019 (Table 3). The greatest contribution to the community abundance was made by copepods (~60%) and rotifers (>25%); the biomass was formed mainly by cladocerans (~80%). The ratio of the abundance of the main zooplankton taxa was unstable from year to year in all three parts of the reservoir; variations were observed in the proportion of both rotifers and copepods (Fig. 4). Variations in the biomass structure were determined mainly by fluctuations in the abundance of Cyclopoida. The average number of veligers of mollusks of the genus Dreissena was 4 times higher in the Votkinsk Reservoir (4.0 ± 0.5 thousand ind./m3) than in the Kama Reservoir; however, their biomass did not exceed 0.01 g/m3.
In both water bodies, the abundance of littoral zooplankton significantly exceeded the abundance of pelagic zooplankton by 1.7–2.0 times (Fig. 6a). On average, it reached 183 ± 47 thousand ind./m3 in the Kama Reservoir and 155 ± 33 thousand ind./m3 in the Votkinsk Reservoir. The littoral community of the Kama Reservoir was dominated by copepods (60% of the community); the community of the Votkinsk Reservoir was dominated by copepods and rotifers (42 and 36%, respectively). The biomass of coastal zooplankton was high in both water bodies (1.5–1.7 g/m3) and did not significantly differ from that observed in the pelagic zone during the same period (1.1–1.4 g/m3) (Fig. 6b).

Abundance (a) and biomass (b) of zooplankton in the pelagic (P) and littoral (L) zones of the Kama and Votkinsk reservoirs in 2014–2019.
The average amount of pelagic zooplankton in 2014–2019 was 20–30% higher in the Kama Reservoir than in the Votkinsk Reservoir (Fig. 6); however, these differences are insignificant. A similar ratio of the abundance was also observed in the littoral zone, where the abundance and biomass differed by 10–15%.
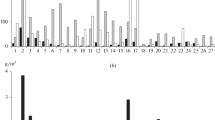
An analysis of the dynamics of the biomass of zooplankton in the Kama Reservoir for 63 years and Votkinsk Reservoir for 54 years showed significant interannual fluctuations during all study periods and a clear trend towards an increase in biomass in the current period in both reservoirs (Fig. 7). In the Kama Reservoir, the biomass of pelagic zooplankton was almost 2.5 times higher in 2014–2019 (1.2 ± 0.2 g/m3) than in the first decade of the existence of the reservoir (0.5 ± 0.1 g/m3). Biomass values comparable to the current ones (1.3 ± 0.1 g/m3) were observed in the 1970s. In the Votkinsk Reservoir, the zooplankton biomass was relatively low until 2010 (0.6 ± 0.1 g/m3). A threefold increase in the community biomass (up to 1.9 g/m3) was recorded in the hot summer of 2010. It usually remained high in subsequent years (until 2018): 1.3 ± 0.2 g/m3 (on average, two times higher than the biomass before 2010). The biomass of zooplankton in the reservoir sharply decreased to 0.1 g/m3 in the cold and high-water year of 2019.

Long-term changes in zooplankton biomass in the Kama (1956–2019) and Votkinsk (1965–2019) reservoirs. (1) Kama Reservoir, average values for the period of 3 to 6 years; (2) Votkinsk Reservoir, average for the period of 3 to 9 years; (3) average for July–August in each year in the Kama Reservoir; and (4) average for July–August in each year in the Votkinsk Reservoir. Source: 1956–1959 (Ulomskii, 1961); 1961–1962 (Kortunova and Serkina, 1980; Serkina, 1971); 1965–1966 (Udalova, 1968); 1971–1975 (Serkina, 1975; Poskryakova, 1977); 1976–1978 (Kortunova and Zueva, 1979; Kortunova and Galanova, 1986); 1979–1982 (Kortunova and Galanova, 1988); 1983–2009 (Kostitsyn et al., 2011; Seletkova, 2015); 2010–2011 (archival data from the Perm Branch of the Russian Federal Research Institute of Fisheries and Oceanography); 2013 (Krainev and Kuznetsova, 2013), and 2014–2019 (this paper).
DISCUSSION
Over 106 zooplankton species live in the Kama and Votkinsk reservoirs; almost half of them are rotifers. According to the archival data from the Perm Branch of the Russian Federal Research Institute of Fisheries and Oceanography, the species richness of the community exceeds 180 species. The most complete composition of zooplankton in the reservoirs of the Kama River is given in the review by Lazareva (2020). All seven species that supplemented this list in our research were identified in unpublished archival materials from the Perm Branch of the Russian Federal Research Institute of Fisheries and Oceanography. They are usually not numerous in the taiga zone of European Russia (Pidgaiko, 1984); therefore, they were not considered invaders.
The analysis of the archival data from the Perm Branch of the Institute of Fisheries and Oceanography made it possible to establish the dates of appearance of southern thermophilic invasive species, including three brackish-water Ponto-Caspian crustaceans, in the Kama and Votkinsk reservoirs (Table 1). Only one freshwater southern rotifer, Pompholyx sulcata, penetrated these water bodies back in the 1980s, two species of the genus Brachionus appeared here in the early 2000s, and the other invaders appeared here in the 2010s (Kuznetsova, 2015; Seletkova, 2015; Krainev et al., 2018; Lazareva, 2020).
Of particular interest is the colonization of Ponto-Caspian crustaceans in the Kama River. The copepod Eurytemora caspica was recorded in both studied reservoirs for the first time in 2012; at that time, the species was determined as E. cf. affinis (Kuznetsova, 2015). For comparison, this species was recorded in the Volga River for the first time in the mid-1980s (Timokhina, 2000). Based on morphological and molecular methods (Sukikh et al., 2020), it has now been established that E. caspica lives in the reservoirs of the Volga and Kama rivers, while typical E. affinis (Poppe, 1880) has not been found anywhere.
The exact habitats of the other two Ponto-Caspian species have been established only since 2016 (Lazareva, 2020). In 2016–2019, the predatory cladoceran Cercopagis pengoi was recorded annually, but only in the near-dam part of the Kama Reservoir. The copepod Heterocope caspia, which is relatively abundant in the second half of summer, presumably appeared in both reservoirs back in the early 2000s. However, the invader was confused with H. appendiculata Sars, 1863, which is common for the taiga zone. H. caspia has been abundant in the reservoirs of the Volga River (Volgograd, Saratov, and Kuibyshev reservoirs) since the mid-1960s (Volga …, 1978; Timokhina, 2000).
An increase in the abundance of cyclopoid copepods of the genera Mesocylops and Thermocyclops, in particular, T. crassus, as a result of warming was recorded in the reservoirs of the Upper Volga (Lazareva and Sokolova, 2015). Similar changes in plankton were recorded in water bodies of Western Europe (Adrian et al., 2006). Until the mid-2000s, the contribution of Cyclopoida to the biomass of pelagic zooplankton varied from 25 to 50% in the Kama Reservoir and from 8 to 30% in the Votkinsk Reservoir (Poskryakova, 1977; Kortunova, 1983; Kortunova and Galanova, 1988; Presnova and Khulapova, 2015; Seletkova, 2015). In 2014–2019, Cyclopoida formed 10–20% of the biomass in the Kama Reservoir and 7–35% in the Votkinsk Reservoir. Until 2010, Mesocyclops leuckarti was mainly dominant in the plankton of both water bodies (Kortunova, 1983; Kortunova and Galanova, 1988; Presnova and Khulapova, 2015; Seletkova, 2015). In 2014–2019, the contribution of Thermocyclops crassus reached 24–30% of the crustacean abundance and 20% of the zooplankton biomass in the Kama Reservoir in some years.
The number of zooplankton in both studied reservoirs is characterized by increasingly large fluctuations year after year (Kortunova, 1983; Kortunova and Galanova, 1988; Seletkova, 2015). This pattern is also confirmed by our data for 2014–2019 (Fig. 7). The biomass of zooplankton in the reservoirs has increased in the current period by 2.5–3.0 times compared to the 1950s–1960s. In the Kama Reservoir, its maxima were recorded in the 1970s and 2010s. In the Votkinsk Reservoir, a significant increase in biomass was observed only in the 2010s. An increase in the amount of summer zooplankton in the past decade has also been recorded for other reservoirs of the cascade, in particular, for the reservoirs of the Upper and Middle Volga (Kopylov et al., 2012; Lazareva et al., 2014). This is presumably determined by the eutrophication of the reservoir ecosystems due to global warming.
CONCLUSIONS
A total of 100 and 69 zooplankton species were recorded in the Kama and Votkinsk reservoirs, respectively, over 6 years (2014–2019). The exact dates of the appearance of ten invasive species of southern origin were determined; 70% of them were recorded in both reservoirs from 2012 to 2016. Among them, three crustacean species (the southern cladoceran Diaphanosoma orghidani and Ponto-Caspian copepods Eurytemora caspica and Heterocope caspia) were annually recorded in more than 30% of samples and locally formed up to 30% of the crustacean abundance. Among southern rotifers, Conochiloides coenobasis, Pompholyx sulcata, and Asplanchna henrietta were abundant in some years (up to 24% Nrot). An increase in the contribution of the thermophilic copepod Thermocyclops crassus to the community (up to 24–30% of the crustacean abundance) was recorded; this species was not previously dominant here. In 2014–2019, the highest abundance of zooplankton in both reservoirs (>150 thousand ind./m3) was observed in the shore zone, while it was 1.7–2.0 times lower in the pelagic zone. The biomass of coastal and pelagic zooplankton did not actually differ and varied from 1.1 to 1.7 g/m3. A clear trend towards an increase in zooplankton biomass was revealed in the reservoirs in the 2010s; the level of biomass became 2.5–3.0 times higher in 2014–2019 than in the 1950s–1960s.
REFERENCES
Adrian, R., Wilhelm, S., and Gerten, D., Life-history traits of lake plankton species may govern their phenological response to climate warming, Global Change Biol., 2006, vol. 12, p. 1652. https://doi.org/10.1111/j.1365-2486.2006.01125.x
Belyaeva, P.G., Mineeva, N.M., Sigareva, L.E., et al., Content of plant pigments in the water and bottom sediments of the Kama River reservoirs, Tr. Inst. Biol. Vnutr. Vod, Ross. Akad. Nauk, 2018, no. 81 (84), p. 97. https://doi.org/10.24411/0320-3557-2018-10007
Edel’shtein, K.K., Vodokhranilishcha Rossii: ekologicheskie problemy i puti ikh resheniya (Reservoirs of Russia: Environmental Problems and Their Solution), Moscow: GEOS, 1998.
Kopylov, A.I., Lazareva, V.I., Mineeva, N.M., et al., Influence of anomalous high water temperatures on the development of the plankton community in the Middle Volga reservoirs in summer 2010, Dokl. Biol. Sci., 2012, vol. 442, p. 11. https://doi.org/10.1134/S0012496612010012
Kortunova, T.A., Zooplankton of the Kama Reservoir and its production, in Kompleksnye issledovaniya rek i vodokhranilishch Urala (Complex Research of Rivers and Reservoirs of the Urals), Perm: Perm. Gos. Univ., 1983, p. 68.
Kortunova, T.A. and Galanova, A.A., Zooplankton of the Kama reservoirs as a food base for fish, in Biologicheskie resursy Zapadnogo Urala, Mezhvuzovskii sbornik nauchnykh trudov (Biological Resources of Western Urals: Interinstitutional Collection of Scientific Papers), Perm, 1986, p. 50.
Kortunova, T.A. and Galanova, A.A., Zooplankton, in Biologiya Votkinskogo Vodokhranilishcha (Biology of the Votkinsk Reservoir), Irkutsk: Irkutsk. Gos. Univ., 1988, p. 50.
Kortunova, T.A. and Serkina, R.A., Distribution and seasonal dynamics of zooplankton of the Kama Reservoir, in Biologicheskie resursy vodoemov Zapadnogo Urala (Biological Resources of Water Bodies of Western Urals), Perm: Perm. Gos. Nats. Issled. Univ., 1980, p. 15.
Kortunova, T.A. and Zueva, E.Yu., The state of the zooplankton of the Kama Reservoir (according to data of 1976), Sb. Tr. Gos. Nauchno-Issled. Inst. Ozern. Rechn. Rybn. Khoz., 1979, no. 2, p. 22.
Kostitsyn, V.G., Pozdeev, I.V., Istomina, A.M., et al., Biologicheskoe obosnovanie k prognozu vylova na 2012 god v osnovnykh rybokhozyaistvennykh vodoemakh Permskogo kraya i v Udmurtskom sektore Votkinskogo vodokhranilishcha (Biological Substantiation for the Forecast of Catch for 2012 in the Main Fishery Water Bodies of the Perm Territory and in the Udmurt Sector of the Votkinsk Reservoir), Perm: Perm. Otd. Gos. Nauchno-Issled. Inst. Ozern. Rechn. Rybn. Khoz., 2011.
Krainev, E.Yu. and Kuznetsova, E.M., Species diversity and distribution of zooplankton in the Votkinsk Reservoir, in Trudy Mezhdunarodnoi nauchno-prakticheskoi konferentsii “Sovremennye problemy vodokhranilishch i ikh vodosborov” (Proc. Int. Sci.-Pract. Conf. “Current Problems of Reservoirs and Their Catchments”), Perm: Perm. Gos. Univ., 2013, p. 137.
Krainev, E.Yu., Tselishcheva, E.M., and Lazareva, V.I., American rotifer Kellicottia bostoniensis (Rousselet, 1908) (Rotifera: Brachionidae) in the Kama Reservoir (Kama River, Russia), Inland Water Biol., 2018, vol. 11, no. 1, p. 42. https://doi.org/10.1134/S199508291801008X
Kuznetsova, E.M., Zooplankton of the Kama Reservoir in summer 2013, in Materialy regional’noi studencheskoi nauchnoi konferentsii “Fundamental’nye i prikladnye issledovaniya v biologii i ekologii,” 14-19 aprelya 2014 g (Proc. Reg. Student Sci. Conf. “Basic and Applied Research in Biology and Ecology,” April 14–19, 2014), Perm: Perm. Gos. Nats. Issled. Univ., 2015, p. 57.
Lazareva, V.I., Long-term changes in the composition and abundance of the zooplankton community in Kama River reservoirs, Inland Water Biol., 2020, vol. 13, no. 2, p. 214. https://doi.org/10.1134/S199508292002025X
Lazareva, V.I. and Sokolova, E.A., Metazooplankton of the plain reservoir during climate warming: biomass and production, Inland Water Biol., 2015, vol. 8, no. 3, p. 250. https://doi.org/10.1134/S1995082915030098
Lazareva, V.I., Mineeva, N.M., and Zhdanova, S.M., Spatial distribution of plankton from the Upper and Middle Volga reservoirs in years with different thermal conditions, Biol. Bull. (Moscow), 2014, vol. 41, no. 10, p. 869. https://doi.org/10.1134/S1062359014100070
Pesenko, Yu.A., Printsipy i metody kolichestvennogo analiza v faunisticheskikh issledovaniyakh (Principles and Methods of Quantitative Analysis in Faunistic Research), Moscow: Nauka, 1982.
Pidgaiko, M.L., Zooplankton vodoemov Evropeiskoi chasti SSSR (Zooplankton of Water Bodies of the European Part of the USSR), Moscow: Nauka, 1984.
Poskryakova, N.P., Kama Zooplankton in August 1975, Biol. Vnutr. Vod: Inf. Byul., 1977, no. 36, p. 57.
Presnova, E.V. and Khulapova, A.V., Structure and distribution of zooplankton in the central area of the Votkinsk Reservoir, Vest. Perm. Univ., Ser. Biol., 2015, no. 4, p. 366.
Seletkova, E.B., Zooplankton of the Kama Reservoir, in Biotsenozy rek i vodokhranilishch Zapadnogo Urala (Biocenoses of Rivers and Reservoirs of Western Urals), St. Petersburg: Gos. Nauchno-Issled. Inst. Ozern. Rechn. Rybn. Khoz., 2015, p. 136.
Serkina, R.A., Seasonal dynamics of zooplankton of the Kama Reservoir, Uch. Zap. Perm. Gos. Univ., 1971, no. 261, p. 126.
Serkina, R.A., The state of zooplankton in the area of pollution of the Votkinsk Reservoir, in Biologicheskie resursy Kamskikh vodokhranilishch (Biological Resources of Kama Reservoirs), Perm: Permsk. Gos. Univ., 1975, p. 143.
Sukhikh, N.M., Lazareva, V.I., and Alekseev, V.R., Copepod Eurytemora caspica Sukhikh et Alekseev, 2013 (Crustacea, Calanoida) in Volga and Kama River reservoirs, Inland Water Biol., 2020, vol. 13, no. 2, pp. 198–205. https://doi.org/10.1134/S1995082920020145
Timokhina, A.F., Zooplankton kak komponent ekosistemy Kuibyshevskogo vodokhranilishcha (Zooplankton as a Component of the Kuibyshev Reservoir Ecosystem), Tolyatti: Inst. Ekol. Volzh. Basseina, Ross. Akad. Nauk, 2000.
Udalova, N.N., Zooplankton of the Votkinsk Reservoir on the Kama River in the open period in 1965–1966, in Konferentsiya po izucheniyu vodoemov basseina Volgi, Tezisy dokladov (Conf. on the Study of Water Bodies in the Volga River Basin, Abstracts of Papers), Tolyatti: Inst. Ekol. Volzh. Basseina, Ross. Akad. Nauk, 1968, p. 125.
Ulomskii, S.N., Raw weight of mass forms of lower crustaceans of the Kama Reservoir and Some Lakes of the Urals and Trans-Urals, Tr. Ural. Otd. Gos. Nauchno-Issled. Inst. Ozern. Rechn. Rybn. Khoz., 1961, vol. 5, p. 200.
Volga i ee zhizn’ (Volga and Its Life), Leningrad: Nauka, 1978.
Funding
This study was performed as part of State Task AAAA-A18-118012690106-7.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests. The authors declare that they have no conflicts of interest.
Statement on the welfare of humans or animals. All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Additional information
Translated by D. Zabolotny
Abbreviations: Nzoo—zooplankton abundance, Bzoo—zooplankton biomass, Nrot—number of rotifers, Ncr—number of crustaceans
Rights and permissions
About this article

Cite this article
Tselishcheva, E.M., Lazareva, V.I. Long-Term Dynamics of Zooplankton in the Kama and Votkinsk Reservoirs. Inland Water Biol 14, 415–426 (2021). https://doi.org/10.1134/S1995082921040118
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1995082921040118