
Abstract
In August 2016, the pelagic zooplankton (Cladocera, Copepoda, and Rotifera) and meroplankton (veligers of mollusks of g. Dreissena) were studied in four reservoirs of the Kama River (Kama, Votkinsk, and Nizhnekamsk reservoirs, and the Kama Reach of the Kuibyshev Reservoir). A total of 108 species were found; 14 species were new for the Kama River, including five crustaceans introduced from the Caspian Sea. Three of them (Heterocope caspia, Eurytemora caspica, and Cercopagis pengoi) spread northwards to the Kama Reservoir; two species (Cornigerius maeoticus and Calanipeda aquaedulcis) reached the upstream part of the Kuibyshev Reservoir’s Kama Reach. It is revealed that five southern species (Heterocope caspia, Diaphanosoma orghidani, Pompholyx sulcata, Conochiloides coenobasis and Asplanchna henrietta) first recorded in the Kama River in 2016, locally form high abundance. The summer zooplankton biomass averages 1.8 g/m3 in the Kama Reservoir, 1.3 g/m3 in the Niznekamsk Reservoir, 1.0 g/m3 in the Votkinsk Reservoir, and 0.5 g/m3 in the Kuibyshev Reservoir; its values are similar to those recorded in the 2000s. The changes in species composition, biological invasions, patterns of spatial distribution, and long-term changes in the zooplankton biomass are discussed.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
INTRODUCTION
The Kama River is the left-bank, largest, and most water-rich tributary of the Volga River. Before damming, the length of the river was >2000 km; the average long-term water discharge in the lower reaches is >4000 m3/s (Volga …, 1978). The cascade of reservoirs of the Kama River was created in the period 1954–1979 (Edelshtein, 1998). The great influence of the Kama zooplankton on the composition and structure of the Volga River communities downstream its confluence with the Kama was noted as early as in the 1970s (Volga …, 1978).
To date, the zooplankton of the upstream Kama and Votkinsk reservoirs (Aleksevnina and Presnova, 2017; Kortunova, 1983; Kortunova and Galanova, 1988; Kuznetsova, 2015; Poskryakova, 1977; Presnova and Khulapova, 2015; and Seletkova, 2015) is best studied; the communities of the Nizhnekamsk Reservoir and of the Kuibyshev Reservoir’s Kama Reach (Kuibyshevskoye …, 1983, 2008; Poskryakova, 1977) are studied much more poorly. In August 1975, as part of the IBIW RAS expedition, the reservoirs of the Kama River from its confluence with the Volga River to the mouth of the Vishera River was studied; the zooplankton of all four reservoirs is described by Poskryakova (1977).
Despite the large number of publications on zooplankton of the two upper Kama reservoirs, there is still no analysis of the long-term dynamics of its composition and structure. In most published papers, only the state of the community at the time of the study was ascertained. At the same time, in the Volga reservoirs and other waterbodies, a significant change in characteristics such as biological invasions, change of dominants, an increase in the proportion of cladocerans and copepods, and community productivity resulting from the transformation of ecosystems due to climate warming (Lazareva et al., 2018a, 2018b; Fefilova et al., 2014; Adrian et al., 2006; Lazareva and Sokolova, 2015) took place. The fast northward advance of Ponto-Caspian crustaceans, some of which became very abundant in the Kuibyshev Reservoir and the Kama waterbodies, along the Volga River was revealed (Lazareva et al., 2018a, 2018b; Timokhina, 2000; Lazareva, 2019).
The goal of this paper is to analyze the dynamics of composition and structure of pelagic zooplankton of the Kama River reservoirs for more than 40 years; original and published data are used for the analysis.
MATERIALS AND METHODS
Study region. The dammed part of the Kama River is located downstream the mouth of the Vishera River and includes three reservoirs (Kama, Votkinsk, and Nizhnekamsk); a scheme of their locations is given by Belyaeva et al. (2018). The vast mouth region of the Kama, after impounding of the Kuibyshev Reservoir with the Volga waters, formed two of its eight reaches (Kamsky and Volgo-Kamsky) (Dzyuban, 1960). The trophic status of all Kama reservoirs in the summer low water of 2016 as assessed by the chlorophyll a content in plankton was classified as eutrophic. By the sum of chlorophyll and pheopigment contents in bottom sediments, the Kama Reservoir was classified as eutrophic; the Votkinsk and Nizhnekamsk reservoirs were classified as mesotrophic (Belyaeva et al., 2018).
The Kama (Perm, Verkhnekamsk) reservoir is an upstream manmade lake in the Kama River system impounded in 1954. Its area is 1915 km2, average depth is 6.4 m (maximum 30 m), and the conditional water turnover coefficient value is 4.4 year-1 (Edelshtein, 1998). Downstream the river, between the cities of Perm and Tchaikovsky, there is the Votkinsk Reservoir, impounded in 1966. Its area is 1120 km2, average depth 8.4 m (maximum 28 m), and the conditional water turnover coefficient value is 5.7 year–1 (Edelshtein, 1998). In 1979, between the cities of Tchaikovsky and Naberezhnye Chelny, the Nizhnekamsk Reservoir was created; it has not yet been filled to the planned level and transits the Kama River water discharge into the Kuibyshev Reservoir. The reservoir area is 1000 km2, the average depth is 8.0 m (maximum 14 m), and the conditional water turnover coefficient value is 6.6 year–1 (Edelstein, 1998). The Kuibyshev Reservoir is one of the largest valley reservoirs in the Middle Volga basin, impounded in 1957. Its surface area is 5900 km2, average depth is 8.9 m (maximum >40 m), and the conditional water turnover coefficient value is 4.2 year-1 (Edelshtein, 1998).
The combined expedition run by the Papanin Institute for Biology of Inland Waters, Russian Academy of Sciences (IBIW RAS) in August 2016 actually repeated the route of the previous one (1975). In the second half of the month along the flooded riverbed of the Kama River, the Volga–Kama and Kama reaches of the Kuibyshev Reservoir, as well as the Nizhnekamsk, Votkinsk, and Kama reservoirs, were studied. Samples were collected at 6–8 sampling stations in the pelagic zone of each reservoir in the region of greatest depths. The southernmost sampling point was located at the confluence of the Kama River with the Volga River against the Kamskoye Ust’ye Village (55°14.776′ N, 49°16.355′ E) in the Volga–Kama Reach of the Kuibyshev Reservoir; the northernmost point was in the upper part of the Kama Reservoir near the town of Usolye (59°26.242′ N, 56°41.322′ E).
Crustaceans and rotifers were studied in total zooplankton samples, which were collected using a Juday net with an inlet diameter of 12 cm and a sieve with 105 μm mesh diagonal. The samples were preserved with 4% formalin and examined in the laboratory under a StereoDiscovery-12 stereo microscope (Carl Zeiss, Jena). The abundance and biomass of each detected species were determined in the samples, and the total values of these indices were calculated for four large taxonomic groups (Cladocera, Cyclopoida, Calanoida, and Rotifera) and the entire community. The species with an abundance of ≥10% of the total number of crustaceans or rotifers were considered dominants.
The concentration of dissolved oxygen, temperature, and water electric conductivity were measured using a YSI ProODO (YSI Inc., United States) hand-held probe with anoptical gauge.
RESULTS
Hydrophysical conditions. The depth at the sampling points on the Kama River bed varied from 8 to 23 m; its average values in all reservoirs were similar (11–16 m). The white-disc water transparency was 90–170 cm. The color was low, 25°–45° Cr–Co; the highest values are noted in the Votkinsk Reservoir. The water pH level in all reservoirs varied within 7.8–8.6 limits.
The water electrical conductivity in the Kama River in August, reflecting the level of its mineralization, increased stepwise upstream from the Volga–Kama Reach of the Kuybyshev Reservoir (275–360 μS/cm)Footnote 1 to the upstream part of the Kama Reservoir (730–860 μS/cm). In some of sections of the latter (near the cities of Berezniki and Usolye, downstream from the confluence of the In’va and Kos’va rivers) in the near-bottom layer, it reached 3480–4870 μS/cm. This relates to the discharge into the Kama River channel of natural brines widespread along the left bank of the Kama River near the city of Solikamsk, as well as to water pollution from wastes of industrial enterprises producing potassium salts (Pecherkin et al., 1980). The minimal water-conductivity values (175–250 μS/cm) were recorded in the Nizhnekamsk Reservoir upstream the Belaya River mouth and in the Votkinsk Reservoir from the dam of the Votkinsk hydroelectric power station to the Tulva River mouth.
During the study period in August 2016, the water temperature was very high (>24°С), which is 6–6.4°С higher than normal. The norm for August is 18°С in the Kama and Votkinsk reservoirs and 19°С in the Nizhnekamsk and Kama part of the Kuibyshev Reservoirs (Long-term …, 1988). The highest temperature (to 25.5°C) of the surface layer was observed in the Kuibyshev Reservoir. Because of this phenomenon, a rise in water temperature near large cities (+1°С near Tchaikovsky and downstream of Perm) and the zone of discharge of heated water (+0.3°С at Perm electric power station) was weakly expressed. At the bottom of waterbodies in the largest part of the flooded Kama River channel, the temperature was 0.1–1.1°С lower than at the surface. The maximum difference between the surface and bottom water temperatures (3–7°С) was recorded in the conditions of direct stratification in deepwater (13–24 m) areas near the dam of the hydroelectric power station in the Kama and Votkinsk reservoirs. The minimum water temperature at the bottom was 18–24°С.
In the trophogenic layer (0–3 m) of all the examined waterbodies during the day, a supersaturation (up to 165%) of water with dissolved oxygen (O2) was observed as determined by the massive development of algae. In the mouth area of the Kama River (Kuibyshev Reservoir) and upstream in the Nizhnekamsk Reservoir, the O2 content in the entire water column exceeded 8 mg/L (>80% saturation). Significant O2 deficiency was observed along the Kama River channel in the Votkinsk and Kama reservoirs at a depth of >8 m. Oxygen content of <1.5 mg/L (<20% saturation) in a layer 1–6 m above the bottom was observed in the downstream part of the Votkinsk Reservoir from the dam to the Tulva River mouth. The same was also recorded almost along the entire Kama River channel within the Kama Reservoir, in the near-dam part of which between the cities of Perm and Dobryanka, the anoxic layer (O2 < 1 mg/L) reached 5–14 m above the bottom.
Composition and structure of zooplankton. In the zooplankton of the Kama River reservoir, 180 species are known with the base built by rotifers and (48%) and cladocerans (33%) (Table 1). In the summer of 2016, 108 species were found in the pelagial (Cladocera 33, Copepoda 22, and Rotifera 53), 92 of which were recorded in the Kuibyshev Reservoir, 61 in the Nizhnekamsk, 75 in the Votkinsk, and 72 in the Kama reservoirs. Most often (>80% of the samples), 7 species were found in the Kuibyshev Reservoir, 13 in the Nizhnekamsk, 21 in the Votkinsk, and 20 in the Kama reservoirs (Table 1). The following species were common in all the Kama reservoirs: cladocerans Limnosida frontosa, Diaphanosoma orghidani, Daphniagaleata, and Bosmina longirostris; copepods Mesocyclops leuckarti, Thermocyclops oithonoides, T. crassus, and Heterocope caspia; rotifers Polyarthra luminosa, P. major, and two subspecies of Euchlanis dilatata. In addition, in the Kama and Votkinsk reservoirs located to north, crustaceans Daphnia cristata, Bosmina crassicornis, B. coregoni, Сyclops vicinus, and Eurytemora caspica and rotifers Trichocerca similis, Polyarthra longiremis, Asplanchna priodonta, A. herriсki, Brachionus angularis, Keratella cochlearis, K. quadrata, Kellicottia longispina, Conochilus unicornis, and Pompholyx sulcata often occurred.
A comparative analysis of new data and lists of zooplankton published based on the materials of the 1970s–2000s (Kortunova and Galanova, 1988; Seletkova, 2015; Timokhina, 2000) revealed that the composition of the species has changed significantly. In each waterbody, from 20 to >30 species (mainly rotifers) were identified that were not previously recorded (Table 1). Most of them (40–65%) are common for modern zooplankton of the Volga reservoirs (Lazareva, 2007; Lazareva et al., 2018a; Ecological …, 2001), as well as other reservoirs in the forest belt of European Russia (Borutsky et al., 1991; Pidgayko, 1984). The remaining species new to the Kama River could be divided into three groups: southern freshwater forms, which expand their range northward due to climate warming; Ponto-Caspian invasive crustaceans that established in the Kama River advancing via the Volga River from the Caspian Sea; and invasive species that entered the Volga basin from other continents.
The first group includes crustaceans Diaphanosoma orghidani and Acanthocyclops americanus and rotifers Asplanchna henrietta, Brachionus diversicornis, B. budapestinensis, Keratella tropica, Pompholyx sulcata, and Conochiloides coenobasis. These species reached the Upper Volga (Lazareva, 2007; Ecological …, 2001) as early as in the last century and are now found in the Kama reservoirs (Table 2). The first finding in the Kama River opposite the Izh River mouth (Nizhnekamsk reservoir, 56° N) of Diaphanosoma mongolianum (>300 ind./m3) together with the more numerous D. orghidani (2000 ind./m3) may be attributed to the same group. Diaphanosoma mongolianum is distributed in the Palearctic up to 57° N, but is more common in its southern part (Korovchinsky, 1987). It should be noted that a number of southern species in the Kama reservoirs form numerous populations (crustaceans 5–20 and rotifers 10–125 thousand ind./m3) and are locally included in the dominant groups of zooplankton (Table 2).
The second group includes Caspian copepods Heterocope caspia, Eurytemora capsica, and Mediterranean Calanipeda aquaedulcis, along with predatory Caspian cladocerans Cornigerius maeoticus and Cercopagis pengoi. Calanipeda aquaedulcis and Cornigerius maeoticus were found only in the Kama part of the Kuibyshev Reservoir (to 55° N); Cercopagis pengoi was also noted in the upstream part of the Votkinsk Reservoir and the near-dam area of the Kama Reservoir (to 58° N); Heterocope caspia and Eurytemora caspica established over all Kama reservoirs to 59° N (Table 2). Among these species, copepod Heterocope caspia is numerous, dominating in the Kuibyshev Reservoir and locally in the Votkinsk Reservoir (Lazareva et al., 2018b; Lazareva, 2019). Other species are relatively innumerous (<5000 ind./m3).
The third group includes one species: American rotifer Kellicottia bostoniensis, introduced in the Volga basin from Western Europe and rapidly spreading eastwards (Zhdanova et al., 2016). By 2012 it was established all over the Kama Reservoir, but in August 2016 this species was absent from the plankton (Krainev et al., 2018).
In addition to the above species, a group (ten species) of new but rare forms that are either innumerous or only found as a single specimen (Table 1) was registered. The rotifer Collotheca pelagica was found among them most often in all reservoirs, especially in the Kama. This species is also common in the Upper and Middle Volga (Lazareva et al., 2018a). The first finding of copepod Eudiaptomus transylvanicus (20 ind./m3) in the near-dam area of the Kama Reservoir is worth noting. This species is widespread in the waterbodies of the forest belt (Borutsky et al., 1991), but was not previously found in the reservoirs of the Volga and Kama rivers (The Volga …, 1978; Lazareva, 2007; Lazareva et al., 2018a).
In our own samples in the Nizhnekamsk Reservoir, a new species of g. Bythotrephes, B. lilljeborgi Korovchinsky, 2018 (Korovchinsky, 2018), was found, while other representatives of this genus dwelling in the Kama and Volga rivers are classified as hybrids of Bythotrephesbrevimanus × B. cederströmii (Korovchinsky, 2019). They were described earlier as Bythotrepheslongimanus (Leydig, 1860).
The most numerous zooplankton of each reservoir were 3–6 species of crustaceans and 2–4 species of rotifers. In August, the base (60–90%) of the crustacean number (Ncr) in all reservoirs was formed by copepods; Mesocyclops leuckarti (on average 20–60 thousand ind./m3 or 40–60% Ncr) and Thermocyclops crassus (7–30 thousand ind./m3 or 10–20% Ncr) dominated. T. oithonoides is also abundant in the northern Votkinsk and Kama reservoirs (24–30 thousand ind./m3, or 17–25% Ncr); in the southern Kuibyshev Reservoir, it is invasive Heterocope caspia (8 thousand ind./m3 or 18% Ncr).
Locally, most often at the mouth areas of the large Kama tributaries, high numbers were formed by cladocerans. For instance, in the Kondas River mouth (Kama Reservoir), 16% of Ncr was formed by small (0.3–0.8 mm) Diaphanosoma orghidani (21 000 ind./m3) and Chydorus sphaericus (28 000 ind./m3), while large (up to 1.8 mm) Daphnia galeata was especially abundant (15–26 thousand ind./m3 or 15–20% Ncr) near the Obva and In’va river mouths in the Kama, the Tulva River in the Votkinsk, and Siva and Izh rivers in the Nizhnekamsk reservoirs.
High abundance (10–220 thousand ind./m3) was a characteristic of 11 rotifer species: Brachionus angularis, Pompholyx sulcata, Keratella cochlearis, K. quadrata, Conochilus hippocrepis, Euchlanis dilatata lucksiana, Conochiloides coenobasis, Polyarthra luminosa, P. major, Synchaeta pectinata, and Asplanchna henrietta.
Their abundance in the reservoir pelagial zooplankton was distributed extremely unevenly (coefficient of variation value ranged 150–430), most species dominated locally in certain parts of the reservoirs. The maximal (>300 000 ind./m3) number of rotifers (Nrot) was recorded in the Kama Reservoir near the mouth of the Kondas River (dominated by Brachionus angularis: 220 000 ind./m3 or >70% Nrot) and the upper river section of the Nizhnekamsk Reservoir from the dam of the Votkinsk hydroelectric power station to the city of Sarapul (Table 3). In the latter, a large group of rotifers was numerous, in which Pompholyx sulcata (90–125 thousand ind./m3, 26% Nrot), Keratella cochlearis (100–119 thousand ind./m3, 26% Nrot), Conochilus hippocrepis (100–113 thousand ind./m3, 26% Nrot), and Euchlanis dilatata lucksiana (33–39 thousand ind./m3, 9% Nrot) prevailed.
In the upstream section of the Kama reservoir near the city of Berezniki, with a rotifer number of 95 000 ind./m3, Conochiloides coenobasis (21 000 ind./m3, 22% Nrot) and Brachionus angularis (16 000 ind./m3, 17% Nrot) dominated. In the section from the confluence of the In’va and Kos’va rivers to the dam of the Kama hydroelectric power station, upon a relatively small total number of the group (6–52 thousand ind./m3), the dominance was weakly expressed; Keratella cochlearis (<25% Nrot) and Polyarthra major (<17% Nrot) prevailed.
Between the mouth of the Nytva and Tulva rivers in the midstream part of the Votkinsk Reservoir, at 60–120 thousand ind./m3 rotifers, Polyarthra luminosa (reaching 18 000 ind./m3, an average of 17% Nrot) and Synchaeta pectinata (reaching 36 000 ind./ m3, 11% Nrot) dominated. Near the mouth of the Ocher River, along with these species, Asplanchna henrietta was relatively numerous (14 000 ind./m3, 11% Nrot). From the bay near the village of Elovo to the dam of the Votkinsk hydroelectric power station, the number of rotifers reached 15–60 thousand ind./m3; Keratella cochlearis (<20 000 ind./m3, 18% Nrot) and K. quadrata (<13 000 ind./m3, 9% Nrot) prevailed and, in the near-dam area, Conochilus hippocrepis formed about 80% of Nrot (13 000 ind./m3).
At the downstream part of the Nizhnekamsk Reservoir, the number of rotifers was very small (<10 000 ind./m3); only the number of Conochilus hippocrepis at the mouth of the Belaya River reached 5000 ind./m3. The number of rotifers in the Kama and Volga–Kama reaches of the Kuibyshev Reservoir was just as low; only at the mouth of the Vyatka River it increased to >70 000 ind./m3 (Fig. 1a). Brachionus angularis (31 000 ind./m3, 42% Nrot) and Conochiloides coenobasis (24 000 ind./m3, 33% Nrot) dominated there. In the rest of the Kama River, within the borders of this reservoir, along with the two species indicated above, a small number (1–1.5 thousand ind./m3) was formed by Polyarthra luminosa, P. major, Brachionus calyciflorus, and Asplanchna henrietta.

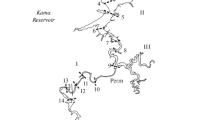
Distribution of abundance (thousand ind./m3) of the main groups of zooplankton (a) and total biomass (g/m3) (b) along the longitudinal profile of the Kama River from the Kama Reservoir headwaters near the town of Usolye to the confluence with the Volga River in the Kuibyshev Reservoir near the village of Atabaevo. (1) Cladocera, (2) Copepoda, and (3) Rotifera. Abscissa, sampling stations in the reservoirs: 1–8 Kama, 9–15 Votkinsk, 16–21 Nizhnekamsk, and 22–27 Kuibyshev.
Number and biomass. The total abundance of zooplankton in the Kama part of the Kuibyshev Reservoir was on average three times lower, while the amount of meroplankton (veligers of the mollusk Dreissena) was much higher than in the other three reservoirs (Table 3). Rotifers (25–68% of the total number) and cyclopoid copepods (25–61%) prevailed everywhere. The abundance of the community, especially of rotifers and cladocerans, was much (2–10 times) lower in the midstream part of the Nizhnekamsk Reservoir and remained low over all distance to the Kama River mouth (Fig. 1a). The contribution of meroplankton to the total abundance of zooplankton reached 25% in the Kuibyshev Reservoir; in the other reservoirs it did not exceed 6%.
The largest zooplankton biomass was recorded in the Kama and Nizhnekamsk reservoirs; the fewest were in the Kuibyshev Reservoir (Table 3). In the Kama Reservoir, it exceeded 1 g/m3 over almost the entire Kama River channel and reached a maximum (~4 g/m3) at the Kondas Rriver mouth (Fig. 1b). In the Votkinsk Reservoir, its high values (1.1–1.8 g/m3) were noted at the Tulva River mouth and in the area from Votkinsk to the dam of the hydroelectric power station. In the Nizhnekamsk Reservoir, very large biomass variations (to a magnitude of 30 times) were observed with maxima (1.7‒2.9 g/m3) in the upper section from the Siva River mouth to the city of Sarapul and at the Izh River mouth. In the Kuibyshev Reservoir, the zooplankton biomass reached 1 g/m3 only in the upper Kama Reach below the town of Elabuga.
The base of biomass was formed by cladocerans (27–63% of the total biomass) and cyclopoid copepods (25–49% Вsum) everywhere, dominated by Mesocyclops leuckarti (22–29%) and Daphnia galeata (15–43%). In the Kuibyshev Reservoir, together with the above species, a significant proportion of the biomass (32%) was formed by Calanoida, mainly Heterocope caspia (24%). The contribution of this invader to the zooplankton biomass was also high (20%) in the Nizhnekamsk Reservoir; in the northern Votkinsk and Kama it was only 7–8%.
DISCUSSION
In the summer of 2016, the IBIW RAS expedition turned out to be the first time in 40 years in which the zooplankton of all Kama River reservoirs was researched in a single week. As a result, new habitats of many, mainly southern, species have been identified. A review of the spread of Ponto-Caspian brackish-water crustaceans in the reservoirs of the Volga–Kama cascade is given in (Lazareva et al., 2018b; Lazareva, 2019). Here we only note that until 2016 the invaders from the Caspian and Azov seas were not found upstream the dam of the Nizhnekamsk hydroelectric power station in the Kama River (Istomina et al., 2016; Kortunova and Galanova, 1988; Kuibyshevskoye …, 1983; Presnova and Khulapova, 2015; Seletkova, 2015; Timokhina, 2000; Popov, 2011). In 2016, it was found that three (Heterocope caspia, Eurytemora caspica, and Cercopagis pengoi) of the five invaders of this group spread northward along the river to the Kama Reservoir. The other two species, Cornigerius maeoticus and Calanipeda aquaedulcis, had penetrated only to the Kama River headwaters of the Kama reach in the Kuibyshev Reservoir by 2016.
In 2015–2016 Eurytemora gr. affinis, inhabiting the reservoirs of the Volga and Kama rivers, was identified as E. caspica (Lazareva et al., 2018a, 2018b; Lazareva, 2019), which is confirmed by molecular genetic studies (Sukhikh et al., 2020). E. caspica is a new species recently described using materials from the Volga River delta and Northern Caspian (Sukhikh and Alekseev, 2013). The typical Eurytemora affinis (Poppe, 1880) was not found in any sample.
Among the southern species expanding their ranges northward, in the second half of the 2000s, two species of rotifers of g. Brachionus (B. diversicornis and B. budapestinensis) (Podshivalina and Yakovlev, 2012) were indicated. In the same paper, the findings of Keratella tropica in the Kama River and its left-bank tributaries south of the city of Naberezhnye Chelny are noted. I found this species in the same section of the river (Volga–Kama and Kama reaches of the Kuibyshev Reservoir), while Brachionus diversicornis and B. budapestinensis were common in all Kama River reservoirs (Table 2).
The information appeared about findings (not indicating the date) of the East Asian copepod Thermocyclops taihokuensis (Harada, 1931) (syn. T. asiaticus) in the lower part of the Votkinsk Reservoir and in the Izh River mouth in the downstream zone of the Nizhnekamsk Reservoir (Sidorovsky et al., 2018). In 2016, this species was absent in our samples from the Kama reservoirs, but in 2017 it was found in the Volga River downstream the city of Volgograd (Lazareva et al., 2018b) and in 2018 it was found in the Sura River mouth area (Cheboksary Reservoir) (Zhikharev et al., 2019). It is highly probable that thermophilic species penetrate into reservoirs from tributaries in particular.
As earlier (Kortunova, 1983; Kortunova and Galanova, 1988; Poskryakova, 1977; Presnova and Khulapova, 2015; Seletkova, 2015), in 2016, the main number of crustacean plankton in the Kama River reservoirs was formed by cyclopoid copepods Mesocylops leuckarti, Thermocyclops oithonoides, and T. crassus common for the taiga zone, as well as by cladoceran Daphnia galeata, which in old papers was identified as D. longispina (OF Müller). In the Volga reservoirs, as a result of warming, an increase in the abundance of cyclopoid copepods of gg. Mesocylops and Thermocyclops, especially T. crassus (Lazareva et al., 2018a, 2018b; Lazareva and Sokolova, 2015), was noted. The same was also registered in the reservoirs of Western Europe (Adrian et al., 2006). A possibly similar phenomenon also occurs in the Kama River reservoirs, but we were not able to confirm this, since the data from a short-term survey in August 2016 are difficult to compare with the results of earlier studies carried out at other periods of the vegetation season. However, a tendency towards an increase in the share of copepods in the biomass of zooplankton and a decrease in the contribution of cladocerans (Table 4) was revealed, which is especially noticeable in the Kuibyshev Reservoir, starting from the 2000s. This is mainly due to the spread and rise in the number of large invasive copepod Heterocope caspia.
Until the mid-2000s, in the Kama Reservoir, Bosmina coregoni (common for the second half of the summer) were numerous (Poskryakova, 1977; Seletkova, 2015) and, for the Votkinsk Reservoir, B. longirostris and Eurytemora velox (Kortunova and Galanova, 1988; Presnova and Khulapova, 2015). In August 2016, all three species were innumerous. However, a relatively high number of invasive Heterocope caspia and Diaphanosoma orghidani was revealed. In August of the next 2017, in the Nizhnekamsk Reservoir, Mesocylops leuckarti, Daphnia galeata, and Bosmina coregoni (Melnikova et al., 2019) were noted as common crustacean species, as was also found in the original samples.
In 2016, 11 numerous rotifers were recorded, the peak of development of which falls on the period May to September. However, previously only five (Synchaeta pectinata, Polyarthra major, Keratella cochlearis, K. quadrata, and Euchlanis dilatata), or <50% of them, were earlier attributed to mass species (Kortunova, 1983; Kortunova and Galanova, 1988), which is also true about the relatively recent materials (2011–2015) (Istomina et al., 2016; Kuznetsova, 2015; Seletkova, 2015). In part, this indicates a rapid change in the abundance of short-cycle Rotifera species in the reservoir zooplankton. This change is confirmed by a relatively high (30%) proportion of new species of southern origin among the dominants of this group for the Kama River (Pompholyx sulcata, Conochiloides coenobasis and Asplanchna henrietta). However, it should be recognized as important that in the fishery-related papers small rotifers (Polyarthra, Conochilus, and Synchaeta) with a low contribution to the biomass of the communities are often identified only to the genus level.
In the 1970s, the composition of the zooplankton of the Kuibyshev Reservoir Kama section was almost the same as in its Volzhsky Reach (Kuibyshevskoye …, 1983). Now in the Kama and, especially, in the Volga–Kama reaches, it is far from that observed in the Volga Reach and is close to the composition of the community in the southern part of the reservoir (Lazareva et al., 2018a). The reason for this was the wide spread and high number of the Caspian invaders, most of which have not yet entered the Volzhsky Reach (Lazareva et al., 2018a, 2018b; Lazareva, 2019).
The abundance of zooplankton of all Kama reservoirs largely fluctuates from year to year (Aleksevnina and Presnova, 2017; Istomina et al., 2016; Kortunova, 1983; Kortunova and Galanova, 1988; Kuibyshevskoe …, 2008; Mahotina, 1985; Seletkova, 2015). In 2016, the biomass of the community of most of the reservoirs was close to that observed in the 2000s, and the abundance was approaching the maximum for this period (Table 4). In the Votkinsk and Kama reservoirs, the highest biomass was recorded in the 1970–1980s and, in the Kama reaches of the Kuibyshevsky Reservoir, up to the mid-1990s.
In the 1980–1990s, a peak in biomass was also observed in the Rybinsk Reservoir (Upper Volga) (Lazareva and Sokolova, 2015). Original data (Table 4) on the amount of zooplankton in the Nizhnekamsk Reservoir are close to those recorded in the first years after the reservoir’s impoundment (Mahotina, 1985), while the community structure is similar to that recorded in the second half of the 2000s (Shakirova et al., 2013). A considerably lower abundance of the community (an average of 32 000 ind./m3 and 0.16 g/m3) was observed in August 2017 (Melnikova et al., 2019), which once again confirms its large fluctuations from year to year.
CONCLUSIONS
In 2016, 108 zooplankton species were found in the of Kama reservoirs pelagial, from 61 to 92 species in each of the four water bodies studied; rotifers prevailed (50% of the species list). Fourteen invasive species new to Kama River were identified, represented by three groups: southern freshwater forms (eight species) expanding their range northwards due to climate warming, Ponto-Caspian brackish-water crustaceans (five species) that penetrated the Kama River via the Volga River, and invaders from other continents (one species). The largest abundance of zooplankton in each reservoir is formed by a small set of species: three–six of crustaceans and two–four of rotifers.
Dominant groups are most diverse near the mouth areas of large tributaries to the Kama River and in the upstream sections of all four reservoirs studied. Four southern species (Diaphanosoma orghidani, Pompholyx sulcata, Conochiloides coenobasis, and Asplanchna henrietta), first discovered in the Kama River in 2016, as well as one alien from the Caspian (Heterocope caspia) locally form a high abundance in the Kama River zooplankton, including northern Kama and Votkinsk reservoirs. The zooplankton biomass was maximal in August (>1 g/m3) in the Kama and Nizhnekamsk reservoirs and minimal (0.5 g/m3) in the Kuibyshev Reservoir. In 2016, the zooplankton biomass of the Kama and Votkinsk reservoirs and Kama Reach of the Kuibyshev Reservoir was close to that in the 2000s; the abundance was similar to the largest for this period. In the Nizhnekamsk Reservoir, the biomass of the community was high and similar to that noted in the 1980s.
Notes
All electrical conductivity values are given at temperature 18°C.
REFERENCES
Adrian, R., Wilhelm, S., and Gerten, D., Life-history traits of lake plankton species may govern their phenological response to climate warming, Global Change Biol., 2006, vol. 12, p. 1652. https://doi.org/10.1111/j.1365-2486.2006.01125.x
Aleksevnina, M.S. and Presnova, E.V., The structure of plankto- and benthocenoses of the central area of the Votkinsk Reservoir and its changes in long-term aspect, in Trudy VI Mezhdunar. nauch.-prakt. konf. “Sovremennye problemy vodokhranilishch i ikh vodosborov” (Proc. VI Int. Sci.-Pract. Conf. “Modern Problems of Reservoirs and Their Catchments), Perm: Perm. Gos. Nats. Issled. Univ., 2017, vol. 3, p. 4.
Belyaeva, P.G., Mineeva, N.M., Sigareva, L.E., et al., The content of plant pigments in water and bottom sediments of the Kama River reservoirs, Tr. Inst. Biol. Vnutr. Vod Ross. Akad. Nauk, 2018, no. 81 (84), p. 97.
Borutskii, E.S., Stepanova, L.A., and Kos, M.S., Opredelitel’ Calanoida presnykh vod SSSR (Identification Guide to Calanoida of Fresh Waters of the USSR), Leningrad: Nauka, 1991.
Dzyuban, N.A., Zoning of the Kuibyshev Reservoir, Byul. Inst. Biol. Vodokhran., 1960, nos. 8–9, p. 53.
Edel’shtein, K.K., Vodokhranilishcha Rossii: ekologicheskie problemy i puti ikh resheniya (Reservoirs of Russia: Environmental Problems and Their Solutions), Moscow: GEOS, 1998.
Ekologicheskie problemy Verkhnei Volgi (Environmental Problems of the Upper Volga River), Yaroslavl: Yaroslav. Gos.-Tekhn. Univ., 2001.
Fefilova, E.B., Baturina, M.A., Kononova, O.N., et al., Long-term changes in communities of aquatic organisms in Kharbey Lakes, Zh. Sib. Fed. Univ., Biol., 2014, no. 7, p. 240.
Istomina, A.M., Belyaeva, P.G., Istomin, S.G., et al., The current state of the plankton, benthic, and ichthyofauna of the Votkinsk Reservoir, in Sovremennoe sostoyanie bioresursov vnutrennikh vodoemov i puti ikh ratsional’nogo ispol’zovaniya: Mater. dokl. Vseros. konf. s mezhdunar. uchastiem, posvyashchennoi 85-letiyu Tatarskogo otdeleniya Gos. nauch.-issled. inst. ozer. i rech. ryb. khoz. (Kazan’, 24—29 oktyabrya 2016 g.) (The Current State of Biological Resources of Inland Water Bodies and the Ways of Their Sustainable Use: Proc. All-Russia Conf. with Int. Part. Devoted to the 85th Anniversary of the Tatar Branch of the State Research Institute of Lake and River Fisheries (Kazan, October 24–29, 2016)), Kazan: Gos. Nauchno-Issled. Inst. Ozern. Rechn. Rybn. Khoz., 2016, p. 430.
Korovchinsky, N.M., A study of Diaphanosoma species (Crustacea: Cladocera) of the “mongolianum” group, Int. Rev. gesamt. Hydrobiol., Hydrogr., 1987, vol. 72, no. 6, p. 727.
Korovchinsky, N.M., Further revision of the genus Bythotrephes Leydig (Crustacea: Cladocera: Onychopoda): redescription of B. brevimanus Lilljeborg, reevaluation of B. cederströmii Schödler, and description of a new species of the genus, Zootaxa, 2018, vol. 4379, no. 3, p. 347. https://doi.org/10.11646/zootaxa.4379.3.2
Korovchinsky, N.M., Morphological assessment of the North Eurasian interspecific hybrid forms of the genus Bythotrephes Leydig, 1860 (Crustacea: Cladocera: Cercopagididae), Zootaxa, 2019, vol. 4550, no. 3, p. 340. https://doi.org/10.11646/zootaxa.4550.3.3
Kortunova, T.A., Zooplankton of the Kama Reservoir and its production, in Kompleksnye issledovaniya rek i vodokhranilishch Urala (Integrated Studies of Rivers and Reservoirs of the Urals), Perm: Perm. Gos. Univ., 1983, p. 68.
Kortunova, T.A. and Galanova, A.A., Zooplankton, in Biologiya Votkinskogo vodokhranilishcha (Biology of the Votkinsk Reservoir), Irkutsk: Irkutsk. Gos. Univ., 1988, p. 50.
Krainev, E.Yu., Tselishcheva, E.M., and Lazareva, V.I., American rotifer Kellicottia bostoniensis (Rousselet, 1908) (Rotifera: Brachionidae) in the Kama Reservoir (Kama River, Russia), Inland Water Biol., 2018, vol. 11, no. 1, p. 42. https://doi.org/10.1134/S199508291801008X
Kuibyshevskoe vodokhranilishche (nauchno-informatsionnyi spravochnik) (Kuibyshev Reservoir (Scientific Information Guide)), Tolyatti: Inst. Ekol. Volzh. Basseina Ross. Akad. Nauk, 2008.
Kuibyshevskoe vodokhranilishche (Kuibyshev Reservoir), Leningrad: Nauka, 1983.
Kuznetsova, E.M., Zooplankton of the Kama Reservoir in summer 2013, in Fundamental’nye i prikladnye issledovaniya v biologii i ekologii: Mater. reg. studencheskoi nauch. konf. 14–19 apr.2014 g. (Fundamental and Applied Research in Biology and Ecology: Proc. Reg. Student Sci. Conf. April 14–19, 2014), Perm: Perm. Gos. Nats. Issled. Univ., un-t, 2015, p. 57.
Lazareva, V.I., Composition of crustacean and rotifers of the Rybinsk Reservoir, in Ekologiya vodnykh bespozvonochnykh (Ecology of Aquatic Invertebrates), Nizhny Novgorod: Vektor TiS, 2007, p. 127.
Lazareva, V.I., Spreading of alien zooplankton species of Ponto-Caspian origin in the reservoirs of the Volga and Kama rivers, Russ. J. Biol. Invasions, 2019, no. 4, p. 328. https://doi.org/10.1134/S2075111719040040
Lazareva, V.I. and Sokolova, E.A., Metazooplankton of the plain reservoir during climate warming: biomass and production, Inland Water Biol., 2015, vol. 8, no. 3, p. 250. https://doi.org/10.1134/S1995082915030098
Lazareva, V.I., Sabitova, R.Z., and Sokolova, E.A., Features of the structure and distribution of late summer (August) zooplankton in the Volga reservoirs, Tr. Inst. Biol. Vnutr. Vod Ross. Akad. Nauk, 2018a, no. 82 (85), p. 28.
Lazareva, V.I., Sabitova, R.Z., Bykova, S.V., et al., Distribution of the summer zooplankton in the cascade of reservoirs of the Volga and Kama rivers, Tr. Inst. Biol. Vnutr. Vod Ross. Akad. Nauk, 2018b, no. 83 (86), p. 62.
Makhotina, M.K., Kharakteristika zooplanktona Nizhnekamskogo vodokhranilishcha na pervom etape ego formirovaniya (Characterization of Zooplankton of the Nizhnekamsk Reservoir at the First Stage of Its Formation), St. Petersburg: Gos. Nauchno-Issled. Inst. Ozern. Rechn. Rybn. Khoz., 1985, vol. 240, p. 112.
Mel’nikova, A.V., Lyubina, O.S., Gvozdareva, M.A., et al., Water quality assessment in the Nizhnekamsk Reservoir by hydrobiological indicators, Voda: Khim. Ekol., 2019, nos. 3–6, p. 67.
Mnogoletnie dannye o rezhime i resursakh poverkhnostnykh vod sushi. Bassein reki Kamy (Multiannual Data on the Regime and Resources of Surface Land Waters. Kama River Basin), Leningrad: Gidrometeoizdat, 1988, vol. 1, no. 25.
Pecherkin, I.A., Dvinskikh, S.A., Tikhonov, V.P., and Kitaev, A.B., Factors of pollution of water masses of the Kama Reservoir and ways of its prevention, in Biologicheskie resursy vodoemov Zapadnogo Urala (Biological Resources of the Western Urals Reservoirs), Perm: Permsk. Univ., 1980, p. 9.
Pidgaiko, M.L., Zooplankton vodoemov Evropeiskoi chasti SSSR (Zooplankton of Reservoirs of the European Part of the USSR), Moscow: Nauka, 1984.
Podshivalina, V.N. and Yakovlev, V.A., Monitoring the status of small and medium rivers of the forest-steppe Trans-Volga region by zooplankton, Voda: Khim. Ekol., 2012, no. 1, p. 56.
Popov, A.I., Alien species of zooplankton in Saratov Reservoir (Russia, Volga River), Russ. J. Biol. Invasions, 2011, vol. 2, nos. 2–3, p. 126.
Poskryakova, N.P., Zooplankton of the Kama River in August 1975, in Biologiya vnutrennikh vod: Inform. byul. (Inland Water Biol.: Inform. Bull.), Leningrad: Nauka, 1977, no. 36, p. 57.
Presnova, E.V. and Khulapova, A.V., Structure and distribution of zooplankton in the central area of the Votkinsk Reservoir, Vestn. Perm. Univ, Biol., 2015, no. 4, p. 366.
Seletkova, E.B., Zooplankton of the Kama Reservoir, in Biotsenozy rek i vodokhranilishch Zapadnogo Urala (Biocenoses of Rivers and Reservoirs of the Western Urals), St. Petersburg: Gos. Nauchno-Issled. Inst. Ozern. Rechn. Rybn. Khoz., 2015, p. 136.
Shakirova, F.M., Govorkova, L.K., and Anokhina, O.K., The current state of the Nizhnekamsk Reservoir and the possibilities of sustainable use of its fish resources, Izv. Samar. Nauchn. Tsentra Ross. Akad. Nauk, 2013, vol. 15, no. 3 (1), p. 518.
Sidorovskii, S.A., Kargapol’tseva, I.A., and Kholmogorova, N.V., Catalog of crustaceans (Arthropoda, Crustacea) of the Republic of Udmurtia, Amur.Zool. Zh., 2018, vol. 10, no. 2, p. 101.
Sukhikh, N.M. and Alekseev, V.R., Eurytemora caspica sp. nov. from the Caspian Sea—one more new species within the E. affinis complex (Copepoda: Calanoida, Temoridae), Proc. Zool. Inst. RAS, 2013, vol. 317, no. 1, p. 85.
Sukhikh, N.M., Lazareva, V.I., and Alekseev, V.R., Copepod Eurytemora caspica Sukhikh et Alekseev, 2013 (Crustacea, Calanoida) in Volga and Kama River Reservoirs, Inland Water Biol., 2020, vol. 13, no. 2. https://doi.org/10.1134/S1995082920020145
Timokhina, A.F., Zooplankton kak komponent ekosistemy Kuibyshevskogo vodokhranilishcha (Zooplankton as a component of the ecosystem of the Kuibyshev Reservoir), Tolyatti: Inst. Ekol. Volzh. Basseina Ross. Akad. Nauk, 2000.
Volga i ee zhizn' (Volga and Its Life), Leningrad: Nauka, 1978.
Zhikharev, V.S., Gavrilko, D.E., and Shurganova, G.V., Finding of the tropical species Thermocyclops taihokuensis Harada, 1931 (Copepoda: Cyclopoida) in the European part of Russia, Povolzh. Ekol. Zh., 2019, no. 2, p. 264. https://doi.org/10.35885/1684-7318-2019-2-264-270
ACKNOWLEDGMENTS
I am grateful to A.I. Tsvetkov (IBIW RAS) for providing hydrographic characteristics of the Kama River reservoirs.
Funding
This study was carried out as part of State Task no. АААА-А18-118012690106-7.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest. The author declares that she has no conflict of interest.
Statement on the welfare of animals. All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Additional information
Translated by D. Pavlov
Rights and permissions
About this article

Cite this article
Lazareva, V.I. Long-Term Changes in the Composition and Abundance of the Zooplankton Community in Kama River Reservoirs. Inland Water Biol 13, 214–229 (2020). https://doi.org/10.1134/S199508292002025X
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S199508292002025X