
Abstract
Soil microbial communities are considered a prominent soil fertility component since they highly influence soil processes. Despite the current challenges of soil fertility in Ethiopia, the biological properties of soils have been rarely incorporated into soil management decisions and research. The present study describes the bacterial community structure, pattern of soil microbial respiration (SMR), and their relationship with soil physicochemical properties in selected arable soils from four sites, with Luvisols, Cambisols, Vertisols, and Nitisols in the Tigray Regional State of Ethiopia. We employed amplicon sequencing and basal respiration methods to investigate the bacterial community structure and microbial respiration rate, respectively. SMR was higher in the Nitisol samples with a high amount of P2O5, soil organic carbon (SOC), and exchangeable Mg2+. Amplicon sequencing results (400 bp/OTU reads) revealed that the bacterial community was dominated by Actinobacteria, Chloroflexi, and Proteobacteria phyla. The highest ratio of Actinobacteria was found in Vertisol, while that of Chloroflexi and Proteobacteria was detected in Luvisol and Nitisol, respectively. Among the Alphaproteobacteria, order Rhizobiales (including Rhizobiaceae, Beijerinckiaceae, Xanthobacteraceae, Devosiaceae family) was the most abundant nitrogen-fixing bacteria in the soil samples. From the known P-mobilizing and Indole Acetic Acid (IAA) bacteria, members of Bacillus and Pseudomonas were found in low abundance (<1%). Overall, this study highlighted that P2O5, SOC, and Mg2+ probably influenced the variations of bacterial community structure and SMR. The relatively low abundance of important plant growth promoting bacteria (PGPB) in the investigated sites suggests the need for appropriate soil management practices for better crop yield. We recognized that this study was preliminary research, and much work still needs to be done to get a comprehensive view of the agrobacterial community structure and SMR of soils in the study area.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
INTRODUCTION
The land resource is the bases for survival and economic development of agrarian society, like Ethiopia. Agriculture has been the centerpiece of economic growth and accounts for 90% of the export, 85% of employment, and 47% of the GDP [71]. Despite its vital role in overall economic development, Ethiopia’s agricultural sector suffers from mismanagement and unfavorable land-use practices leading to widespread soil degradation. A decline in soil fertility has a huge impact on crop yield and quality and, therefore, on food security [64].
In recent years, the growing scientific recognition of soil biota in soil fertility and land productivity has led to renewed interest in soil biodiversity as a critical component of a strategy towards agricultural sustainability. The microbial fraction of the soil is a vital soil fertility component since soil microbes positively influence soil processes [56]. Bacteria and fungi play crucial roles in litter decomposition processes, soil aggregate formation, nutrient mobilization and uptake, and degradation of pollutants [53]. Plant growth promoting bacteria (PGPB) enhance plant growth by producing numerous plant growth regulators, protecting plants from phytopathogens, providing better nutrient uptake, and increasing plant tolerance towards drought and moisture stress [70]. Microbial communities represent the living and active part of the soil [40]. More than any other organisms, they are highly adaptable to varying conditions and respond to changes rapidly [60]. Usually, the change in soil microbial community activity is measured by basal microbial respiration, the steady rate of respiration in soil resulted from the mineralization of organic matter by microbes [57]. Recently, the development of molecular techniques based on DNA extraction from soil, followed by gene sequencing, uncovered the enormous abundance and diversity of soil microorganisms [51]. Among these techniques, next generation sequencing has been considered a promising molecular method for microbial community analysis [58] due to its precision and high throughput capability [8]. It can be used to examine microbial community structure and community dynamics in response to changes in different environmental variables based on the variation in the 16S rRNA gene [49].
Vertisols, Nitisols, Cambisols, and Luvisols are among the most common reference soil groups [29] in Ethiopia’s agricultural landscapes [31]. Several authors have described and characterized the chemical and physical properties [e.g., 1, 2, 68] and the influence of land use change on those properties [42, 45] of the major soil groups of the country. Yet, the biological properties have rarely been studied and characterized. Very few studies have shown the effects of land use type and soil properties on specific soil microbes, for instance, arbuscular mycorrhizal fungi [74] and symbiotic bacteria [11]. The most comprehensive study was conducted by Delelegn et al. [72]. They investigated soil microbial (bacteria and fungi) diversity and community composition across five land use systems in the Ethiopian highlands and found a significant shift of microbial community due to land use change from forest to agricultural dominated land.
In this study, we described, characterized [22] and classified [29] selected arable soils in the Tigray Regional State of Ethiopia and then examined bacterial community structure and the pattern of soil microbial respiration (SMR) across these soils. The objectives were to i) investigate how SMR and bacterial community structure shift in response to different soil properties; ii) identify primary soil properties that best predict the variation of bacterial community structure and microbial respiration; iii) describe the soil’s bacterial community and investigate the abundance of bacteria, which are important from the crop growing aspects, i.e., PGPB. Understanding the microorganisms’ response to soil and environmental parameters is crucial to crop productivity and the long-term sustainability of soil in agricultural ecosystems [60]. To our knowledge, this is the first attempt to examine both the bacterial community structure and microbial respiration of dominant agricultural soils of Ethiopia. We hope that this study would generate baseline information that is important to support the development of management alternatives to maximize and sustain soil functions in dominant agricultural soils of Ethiopia.
MATERIALS AND METHODS
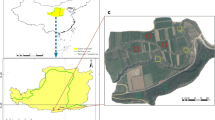
Description of the study area. The study was conducted in Laelay Maichew and Atsbi Wenberta districts. Laelay Maichew district is geographically located at longitude 13°55′53″ E and latitude 38°12′19″ N in the central part of Tigray Regional State; one of the nine Regional States of Ethiopia (Fig. 1). The district altitude varies between 1842 and 2250 m. It is agro-ecologically classified in a semi-arid region and characterized by a short rainy period. The average annual rainfall ranges between 550–750 mm and the mean minimum and maximum temperatures are 11.7 and 26.1°C, respectively [61]. The main World Reference Base (WRB) reference soil groups are Cambisols on undulating plains and rolling landforms; Leptosols on hilly and steep to very steep lands; Vertisols on the flat plateau plains; Luvisols on sloping (5–10%) to moderately steep (15–30%) gradients. The land use is predominantly shrubland (44%), followed by cropland (33%), and settlement area (8%). The farming system is crop farming mixed with livestock husbandry. Teff (Eragrostis tef (Zuccagni) Trotter) cultivation accounted for most arable lands and followed by wheat (Triticum aestivum L.) crop [23].

Location map showing Tigray region within Ethiopia and Laelay Maichew and Atsbi Wenberta districts within Tigray region.
Atsbi Wenberta district is situated in the eastern part of Tigray Regional State. Geographically, it is bounded between 13°30′–13°45′ N and 39°30′–39°45′ E (Fig. 1). The elevation of the district varies from 918 to 3069 m. 75% of the district is upper highlands (2600 m or above), and only 25% is found in midlands (between 1500 and 2600 m) and lowlands (below 1500 m) [14]. The district falls in the sub-tropical agro-climatic zone and has an average daily temperature between 15 and 30°C, and the mean annual precipitation rate is about 529 mm. The district is drought-prone with intense and short duration rainfall; hence, the soils are susceptible to erosion due to high run-off. Lithic Leptosols are the predominant soil types in the area [14]. General site description can be found in Table 1.
Soil profile description and classification. Soil profiles were opened, characterized according to the FAO [22] guidelines, and classified based on World Reference Base for soil resources [29]. Soil sampling locations and details of soil profiles description and classification are presented in supplementary materials. The sites represent four common soil reference groups in Ethiopia: Luvisols, Cambisols, Vertisols, and Nitisols. Nitisols are deep, well-drained, red tropical soils with diffused boundary. They are characterized by a clay-rich ‘nitic’ subsurface horizon with polyhedric, blocky structure elements with shiny ped. Vertisols are heavy clay soils with a high proportion of swelling clay that can swell and shrink in response to a change in soil moisture. Luvisols have a characteristic textural differentiation, with increasing clay content in the subsoil. Cambisols are young soils with limited pedogenetic changes. They underwent various pedological alterations that are enough to distinguish them from other shallow soils such as Leptosols and Regosols but not enough for the development of horizon needed for the classification of other major groups [29].
Soil sampling. The soil sampling was carried out on four agricultural fields that have been under cultivation of teff and wheat (Table 1). Soil samples were taken after crop harvest. At each site, eight points around the main soil profile with a 10 m radius were designated as sampling points. Soil samples from a depth of 0–25 cm were collected from these points, compiled, and mixed thoroughly to make a composite sample. From the composite sample, three subsamples were taken for: a) physiochemical, b) SMR, c) soil bacterial genomic analyses. Soil samples for microbial respiration and DNA analyses were sieved on site through a 2 mm sieve to remove stones, roots, macrofauna, litter materials and transported in icebox. Soil samples for SMR stored in the fridge at 4°C while soils for bacterial genomic analysis at –20°C until analyses (for two weeks). During soil sampling and soil processing, considerable care was taken to avoid contamination between the samples by using gloves and cleaning all equipment with 70% ethanol. Soils for physicochemical analyses were air dried and sieved through 2 mm mesh and stored in room temperature until analyses. Except for the moisture content, all analyses were conducted in the laboratory of the Department of Soil Science, Hungarian University of Agriculture and Life Sciences, Gödöllő, Hungary.
Soil physicochemical measurements and determination of SMR. Soil pH was measured on soil suspended in a solution of deionized water in 1 : 2.5 ratio (w/v) using pH1100L meter (VWR International Ltd, USA) [26]. Soil moisture content (SMC) was measured gravimetrically at 105°C for 24 h with Hotbox Oven (Sanyo Gallenkamp PLC, UK) [27]. Available nitrogen (\({\text{NH}}_{4}^{ + }\)-N and \({\text{NO}}_{3}^{ - }\)-N) was measured using Parnas-Wagner Apparatus (VTR Glass, Germany) [25]. Potassium (K2O) and phosphorus (P2O5) were estimated based on ammonium-lactate solution method (AL method) using Jenway Flame Photometer (Exacta Optech, Italy) and Jenway M6705 UV-VIS Spectrophotometer (Exacta Optech, Italy), respectively [25]. The Walkley-Black technique was used to measure soil organic carbon (SOC) [10]. CaCO3 content was determined by SCM5 Scheibler Calcimeter (Behr Labor Technik GmbH Düsseldorf, Germany) [26]. Cation exchange capacity (CEC) and exchangeable basic cations (Ca2+, Mg2+, K+, and Na+) were extracted following the Mehlich method [6], and the base cations were measured on ICP-OES Spectrometer (HORIBA Jobin Yvon ACTIVA-M, USA). Humic material (E4/E6) was determined following the method given by Page et al. [5] on Jenway M6705 UV-VIS Spectrometer (Exacta Optech, Italy) [5]. All analyses were performed in triplicates.
Determination of SMR. The analysis of SMR followed the ISO 16072:2002(E) guideline [28] and Cheng et al. [17] with minor modification. In short, fifty grams of moist field soil was measured and placed in an airtight jar with a suspended conical flask containing 10 mL of 1.0 M NaOH. The jars were flushed with clean air with low CO2 content, tightly closed and incubated at 22°C for ten days. After ten days, the conical flask was removed and 1 mL BaCl2 was added in the NaOH solution to precipitate the trapped CO2. Three drops of phenolphthalein were added and titrated against 0.5 M HCl. The determination was carried out in triplicates. Controls (triplicate flasks without soil) were also prepared.
Bacterial genomic analysis. DNA isolation and purification. For extracting the total DNA, the method of Högfors-Rönnholm et al. [18] was carried out modified with sonication on ice for one minute and break the slurry in 50 mL falcon tubes for one minute in three rounds after vortexing. DNA was extracted from the collected supernatant using Quick-DNA Soil Microbe Miniprep Kit (Zymo Research co., USA) according to the manufacturer’s instruction. DNA was visualized by gel electrophoresis. Furthermore, the quality and the integrity of isolated DNA was determined by Nanodrop spectrophotometer ND 2000 (Nano-Drop Technologies, Wilmington, DE, USA).
16S rDNA amplicon sequencing and data handling. From the composite soil sample four individual DNA isolation was done and Terminal restriction fragment length polymorphism (TRFLP) analysation was processed for evaluation of the DNA integrity before the NGS amplicon sequencing. For the Terminal restriction fragment polymorphism (TRFLP) Hin6I (5' G˅C G C) and RsaI (5' GT˅AC) restriction enzymes were used. With freeware data analyser PAST software (https:// past.en.lo4d.com/download), the Shannon index (not discussed in the paper) were calculated. These were compared by ANOVA, the standard deviation was under 0.2 in all cases. According to the TRFLP results the pulled DNA was transferred for amplicon sequencing.
The amplicon sequencing and data handling were carried out, according to Révész et al. [21]. Briefly, to assess the soils sample’s bacterial community composition precisely, Illumina 16S rDNA amplicon sequencing was carried out. The variable V3 and V4 region of the 16S rDNA were amplified by using the primers recommended by Klindworth et al. [3], 16S amplicon PCR forward (5′-TCGT CGGCAGCGTCAGATGTG TATAAGAGACAGCCTA CGGGNGGCWGCAG-3′), named 16S Amplicon PCR Forward Primer-S-D-Bact-0341-b-S-17-N and reverse (5′-GTCT CGTGGGCT CGGAGATGTGTATAAGAGAC AGGACTACHVGGGTATCTAATCC-3′), named 16S Amplicon PCR Reverse Primer-S-D-Bact-0785-a-A-21-N primers with Illumina adapter overhanging nucleotide sequences written in bold [3]. PCR reaction mixture in a final volume of 50 μL that contained 12.5 ng of DNA, 0.2 μM of each Illumina 16S primers and 12.5 μL of 2X KAPA HiFi Environ Sci Pollut Res HotStart Ready Mix (KAPABiosystems, London, UK). The temperature profile used was an initial denaturation for 3 min at 95°C, followed by 25 cycles of denaturation for 0.5 min at 95°C, annealing for 0.5 min at 55°C and elongation for 0.5 min at 72°C. The last step was a final extension for 5 min at 72°C. All amplifications were carried out in a ProFlex PCR System (Applied Biosystems by Life Technologies, USA). Amplicons were analyzed under UV light after electrophoresis in 1% (w/v) agarose gel stained with EtBr. Paired-end fragment reads were generated on an Illumina MiSeq sequencer using MiSeq Reagent Kit v3 (600-cycle) (Seqomics Ltd). Read numbers were the following: 77065 for LMH-1, 60 265 for LMH-10, 67 937 for LMH-2, and 58 532 for ATS-3 samples (BF). Primary data analysis (base-calling) was carried out with Bbcl2fastq^ software (v2.17.1.14, Illumina). Reads were quality and length trimmed in CLC Genomics Workbench Tool 9.5.1 using an error probability of 0.05 (Q13) and a minimum length of 50 nucleotides as a threshold. Trimmed sequences were processed using mothur v1.35 [50] as recommended by the MiSeq SOP page (http://www.mothur.org/wiki/MiSeq_SOP downloaded at 22/06/2019) [33]. Sequences were assorted based on the alignment using SILVA 132 SSURef NR99 database [17]. Chimera detection was performed with mothur’s uchime command [55], and the ‘split.abund’ command was also used to remove singleton reads according to Kunin et al. [69]. After all quality control, 38132 reads/sample (400 bp/read) were taxonomically investigated. Taxonomic assignments were made against SILVA release 132, applying a minimum bootstrap confidence score of 80%. Operational taxonomic units (OTUs) were assigned at 97% similarity threshold level, as suggested by Tindall et al. [15] for prokaryotic species delineation. Raw sequence reads were deposited in NCBI SRA under BioProject ID SAMN14390016, SAMN14390017, SAMN14390018, SAMN14390019.
Statistical analyses. Descriptive statistics were employed to examine the variation in SMR and the abundance of bacterial communities among soil types using R software [54]. According to Metacoder R package, the bacterial abundance was calculated, and heat tree was created to visualize the bacterial community structure [75].
RESULTS
Physicochemical properties and SMR. The basic physicochemical properties of soil samples are presented in (Table 2). Briefly, SOC was highest in Nitisol and lowest in Cambisol. The soil pH (H2O) ranged from neutral (7.2) to slightly alkaline (7.9). While P2O5 content was highest in Nitisol, followed by Cambisol, Vertisol, and Luvisol, the K2O content showed the opposite trend. Generally, all available forms of nitrogen and exchangeable bases were recorded highest in Nitisol. The rate of SMR ranged from 0.94 in Cambisol to 3.92 µg CO2 g–1 soil h–1 in Nitisol.
Bacteria community of the investigated soils samples. Comparing the genus number according to the 400 bp amplicon sequencing results, Nitisol was the most diverse with 475 genera, followed by Cambisol with 389 genera, Luvisol with 351 and Vertisol with 315, respectively. From these genera’s OTUs (family or order in some case), only 31 were showing relative abundance over 1%, from that a heat tree (Fig. 2), according to the 38 132 reads (400 bp)/sample was created by Metacoder R package in R software. The relative abundance of the taxonomy was made and compared (Table 3). The 31 genera showing abundance over 1% covered the 79% of all OTUs of Vertisol, 72% of Luvisol, 66% of Cambisol, and 59% of Nitisol. According to the heat tree figure, the Luvisol and Cambisol have a slight difference among the identified bacteria genera. The Vertisol showed a shift into the direction of Actinobacteria—Pseudoarthrobacter and Gaiellales family. In Nitisol samples, the Proteobacteria—Alphaproteobacteria class: Sphingomonas genus and Rhizobiaceae family were prevalent. At the phylum level, there was only a slight difference among the investigated soils. The following phyla showed the greatest abundance in the samples: Actinobacteria, Chloroflexi, Proteobacteria, Acidobacteria, Planctomycetes, and Gemmatimonadetes. Actinobacteria was the most abundant phylum in all samples, and accounted (44%) in Vertisol, followed by Luvisol and Nitisol (27%). In the case of Chlorflexi, it was found abundantly in Luvisols (18%). While Proteobacteria showed the greatest abundance in Nitisol (16%) and the lowest in Vertisol (6%), the abundance of Acidobacteria decreased to the lowest (1.38%) in Nitisol. In general, these three phyla (Actinobacteria, Chloroflexi, Proteobacteria) gave the majority among the four soils: Vertisol: 65%, Luvisol: 54%, Nitisol: 51%, and Cambisol: 46% (Table 3).

Heat trees showing the taxonomy abundance of bacterial community of the soil samples, according to the 32812 reads of 400 bp OUT’s of each sample. The size and color of nodes and edges are correlated with the abundance of organisms in each community.
The richest phylum, Actinobacteria, was represented by the following classes over 1% abundance: Thermoleophilia, Actinobacteria, Acidimicrobiia, and Rubrobacteria. In Vertisol, from the total abundance of phylum Actinobacteria (44%), Thermoleophilia and Actinobacteria accounted for 20 and 18%, respectively. The Thermoleophilia class was represented by the Gaiella genus in a very high abundance Vertisol (17%), Luvisol (14%), Cambisol (8%), and Nitisol (3%). The Gaiella genus has only one identified species yet [38]. The greatest abundance of class Actinobacteria was found in Vertisol. Within Proteobacteria, Alphaproteobacteria was the most abundant class, followed by Betaproteobacteria.
Among the Chloroflexi phylum, two classes were abundant over 1%: Chloroflexi and an uncultured class: Bacterium Ellin-6519. All soil samples from Laelay Maichew sites had a greater abundance (over 4%) of Bacterium Ellin-6519 but its abundance was only 0.84% in the Nitisol (Table 3). Interestingly among the genera of the Chloroflexi phylum, the Thermomicrobium roseum species was the most abundant (4–6%) in all soils. This species is an extremely thermophilic bacterium first isolated from an alkaline hot spring in Yellowstone National Park. It is an obligate aerob and grows optimally at 70 to 75°C at a pH of 8.2 to 8.5 [65]. Bacterium Ellin-6519 is one of several uncultured isolates collectively named ‘Ellin’. These isolates were obtained by Janssen and coworker [e.g., 62] Currently, no information about these bacteria is available since none of them described taxonomically [12].
DISCUSSION
The pattern of SMR and its relationship with key soil physicochemical properties. The capacity of soil to provide soil functions can be predicted by the activity and abundance of microbial communities in relation to key soil properties, such as SOC, available nutrients, and pH [7]. All the three pedons in Laelay Maichew (LMH) sites had a vertic property on the surface soils and were similar to most physicochemical properties. Hence, it would be expected that these soils have similar microbial communities and activities, thereby a similar rate of SMR. Consistently, the difference in mean SMR among soils in LMH sites was small, while the difference was big between soils in the ATS-3 site and in LMH sites. The high rate of SMR in Nitisol corresponded to the high amount of SOC and P2O5, which could explain the importance of these parameters for soil microbes’ metabolic activity. SOC is the primary energy source for microbes and considered one of the key soil attributes that greatly influences SMR [57]. Studying 117 different soils with a broad range of physicochemical properties in the Czech Republic, Hofman et al. [32] concluded that SOC strongly correlated with SMR and thus, greatly influenced the overall soil microbial activity. The current study found that soils with lower E4/E6 ratio (more humified) had less SMR. This implies that the reduction of labile SOC could be one of the limiting factors for SMR in the study area [35].
One possible reason for the high rate of SMR in Nitisol might be the very high content of P2O5 related to the continuous amendment of soils with di-ammonium phosphate (DAP). The concentration of P2O5 was almost ten times more in Nitisol compared to other soil types. Currently, most studies concluded that beside C, soil microbial activity in tropical soils is highly limited by P availability. The change in P2O5 concentration in the soil causes a shift in the soil microbial communities’ functional and metabolic potential, resulting in a change in decomposition rates. Phosphorous has a higher affinity to the sorption sites of mineral soils than labile C; thus, the addition of P would release organic matter bound to the sorption sites, which in turn stimulates soil microbial activity [66]. Accordingly, Liu et al. [39] reported that soil respiration significantly increased after long-term P addition in N saturated old-growth tropical forest in southern China, suggesting that soil microbial activity was enhanced by P addition. Conversely, Teklay et al. [67] noted that the SMR pattern was more affected by N than P addition after the amendment of soils with glucose-C, together with N and/or P at Wondo Genet in southern Ethiopia.
The pH has been shown to be one of the significant predictors for SMR [56]. Enzymes, involved in catabolism of carbon substrates, are pH sensitive [7]. Soil pH influences the solubility of SOM and changes the rate of microbial carbon turnover. It also affects the availability and distribution of nutrients, which are essential for microbes to decompose SOM [41]. This study found that soils collected from the ATS-3 site with lower pH (7.2) had four times higher SMR than soils with higher pH at LMH sites (7.7–7.9). Our result was confirmed by the finding of Creamer et al. [56], who noted a decrement of basal respiration with pH. Among the base cations, the concentration of Mg2+, Na+, and K+ was in line with carbon utilization rate. Mg2+, together with other base cations, greatly influences the soil microbial population and their activity since it required for microbial growth and protein synthesis [44]. A similar trend has been previously observed in Irish grassland by Richter et al. [7]. In general, this study highlighted that the variation of SMR in the study area was probably linked with the variation of P2O5, SOC, pH and Mg2+.
The relative abundance of dominant agroecosystem bacteria, their determinants, and ecological roles. Since all the investigated sites were arable land, the focus of this study was on PGPB abundance in the soil. Accordingly, bulk soils were investigated to determine bacterial community differences, particularly PGPB, and their relative abundance with soil type. There is a lack of bulk soil investigations among African soils; rhizosphere soil investigations are more common. In the case of rhizosphere soil, however, the microbial community is shifted to the needs of the specific crop plant and less representing the normal microbial diversity of the soil itself [9]. An agricultural field has an indigenous microbe population, and during the vegetation period, or year-after-year, the most different crop plants could occur in the same field. There should be information about the microbe diversity of most common soil types in a well described region to identify the useful microbes for the crop, for a better yield production management, through the PGPB microbe activity.
In this study, the most represented bacterial phyla were Actinobacteria, Chloroflexi, and Proteobacteria. The predominant PGPB in the studied soil belongs to Proteobacteria. Proteobacteria are a phylum of Gram-negative bacteria commonly found in soil [30]. The Alphaproteobacteria was the most dominant class, occupied the highest abundance in Nitisol. The majority of sequences in the Alphaproteobacteria were affiliated with the order Rhizobiales (including Rhizobiaceae, Beijerinckiaceae, Xanthobacteraceae, Devosiaceae family), which could perform nitrogen fixation, organic matter decomposition, and plant growth promotion [43]. Studies reported that some Proteobacteria, e.g., Pseudomonas spp. and Beijerinckia, can involve both in nitrification and P-solubilization processes [46]. Soil properties are known to condition the microbes’ growth in soils [43]. Generally, pH is found to be a primary driver, that influences the bacterial community at the phylum level [47]. According to Rousk et al. [32], Alphaproteobacteria were most abundant in soils with high pH values. In our case, Alphaproteobacteria (containing Rhizobiales) was only present in 6–8% in the case of LMH soils with 7.7–7.9 pH; thus, the highest abundance was observed in ATS-3 soils (Nitisol) with 15% in 7.2 pH. Similarly, Lauber et al. [16] discussed that the relative abundance of Alphaproteobacteria was not strongly influenced by pH. The SOC content was likely affecting members of this phylum since the abundance of Alphaproteobacteria was more pronounced in soils with high SOC concentration. A similar finding was noted by Tian et al. [52], showed that the relative abundance of Proteobacteria increased with SOC. The results obtained in this study are in line with the copiotroph/oligotroph hypothesis, as suggested by Fierer et al. [48], which postulates that copiotrophs such as Proteobacteria dominate soils with large amounts of available organic carbon. The authors further discussed that SMR, an index of organic carbon availability to microorganisms, was the best predictor of phylum-level abundances. Likewise, in this study, the abundance of Proteobacteria was higher in soils with a high rate of SMR. Similarly, in a study of Jone [59], the relative abundance of Rhizobiales was most strongly correlated with potential carbon mineralization, suggesting the significance of this order in soil carbon dynamics. The influence of crop cultivation system could be another important factor for the high abundance of Rhizobiales in the study area. Farmers in study sites commonly cultivated main crops (teff in Laelay Maichew and wheat in Atsbi Wenberta) in rotation or mixed intercropping with faba bean (Vicia faba L.). Legume plants like faba bean symbiotically associate with rhizobia which the rhizobia supply the legume host with fixed atmospheric dinitrogen in the form of ammonia. Because of its capacity for biological nitrogen fixation with rhizobia bacteria, faba been became a desirable plant to grow in nitrogen deficient areas to promote plant growth [73]. Research reports show that substantial grain yield improvement has been recorded in wheat grown after faba bean [37] and teff intercropping with faba bean [24] in Ethiopia.
The dominancy of Actinobacteria in all our soil samples may be related to their adaptation ability to typical semi-arid environment, similar to the study area [63]. Actinobacteria are gram-positive bacteria that play vital roles in the cycling of organic compounds, production of antibiotic, and synthesis of growth hormones [4, 26]. Among Actinobacteria, Frankiaceae was found in low abundance in the investigated soils. Frankia is the only genus in the family Frankiaceae, which can fix atmospheric nitrogen both in the free-living state and in association with several tree species [4]. Generally, Actinobacteria are sensitive to low pH and grow well in pH ranges between 6–8 [30], might also contribute to their frequent occurrence in our samples. Their abundance which increased with low organic carbon availability, has already been reported in other previous studies, [e.g., 43]. The \({\text{NH}}_{4}^{ + }\)-N concentration was the greatest 7.23 mg kg–1 in Vertisol; this could also affect the abundance of the Actinobacteria playing an important role in the carbon decomposition [34].
Next to Actinobacteria, Chloroflexi were dominant in the soil samples, particularly in Luvisol. Chloroflexi phylum is one of the large phyla that comprises a large group of bacteria that acquire energy and fix CO2 through photosynthesis, thus contributing to carbon dynamics [30]. The previous findings that Chloroflexi prevails in nutrient-poor soils [48], was also noted by this study as the abundance of Chlorflexi was lowest in Nitisol where the highest concentration of SOC was recorded.
Among all IAA and P-solubilizing bacteria we identified, the majority belonged to Bacillus, Paenibacillus, and Pseudomonas, but only in a very low abundance (0.1%). The exceptionally low abundance of P‑solubilizing bacteria in this study contradicts the finding of Tsegaye et al. [76]. The authors investigated beneficial rhizobacteria from tef rhizosphere samples collected during the seedling stage in Ethiopia and reported that 40.5% isolates were able to solubilize phosphate, of which, Pseudomonas, Enterobacter, and Bacillus were the dominant genera. The reason for exceptionally low abundance in our samples could be that our soils were bulk soils, not rhizosphere soils as in the mentioned study. Tropical soils are considered P-deficient because of their high acidity. Soil microbes help to release phosphorus that is only consumed in the soluble form such as monobasic (\({\text{HPO}}_{4}^{{2 - }}\)) and dibasic (\({{{\text{H}}}_{{\text{2}}}}{\text{PO}}_{4}^{ - }\)) phosphate [4]. Here it is also important to mention the crucial role of P-mobilizing fungi in increasing the bioavailability of soil phosphate for plants and overall soil fertility. Among the rhizosphere microbes, Penicillium and Aspergillus spp. are the dominant P-solubilizing filamentous fungi [20]. In Ethiopia, very few studies have been conducted on the effects of phosphate solubilizing fungi on growth and yield of crops. For instance, the effect of phosphate solubilizing fungus on growth and yield of haricot bean (Phaseolus vulgaris L.) plants [20] and teff [13] were evaluated.
The use of PGPR strains with inherent potential for organic phosphorus mobilization offers a way to replace chemical phosphatic fertilizers, thereby minimizing environmental pollution. Altogether the greatest abundance of the known PGPB was shown by the Nitisol with 15%, represented over 1% abundance by the following taxa: Streptomyces, Sphingomonas, Ralstonia genus, Rhizobiaceae, Frankiaceae, Devosiaceae family. The other soils had a total of only 4% PGPB abundance from the previously mentioned taxa.
CONCLUSIONS
Soil samples in this study were originated from long time cultivated fields and chosen as representatives of the agriculture soil types of the region, and the samples were average samples. The results indicated that bacterial diversity and SMR were higher in Nitisol than other soil types. The major phyla were similar, but their relative abundance among soil types differed. Actinobacteria, Chloroflexi and Proteobacteria dominated the bacterial community structure in the study area. P2O5, SOC, and Mg2+ probably explained the variability of bacterial community structure and pattern of SMR.
The abundance of well-known agroecosystem bacteria members (PGPB) was between 4–9%. Among the PGPB group the members of the Bacillus, Pseudomonas, Azospirilum genera were in very low abundance (0.4–0.1%). This could affect the P-circle of the investigated soils negatively. These results are marking the importance of adopting practices that encourage the elevation of SOC and P such as non-removal of crop residues from agricultural lands, application of manure, and most importantly, adaptation of P-mobilizing bacteria groups as PGPR fertilizer for better crop yield. This preliminary study could provide baseline information for further bacterial genomic studies in this region and contributes to the identification of soil bacteria that are beneficial to agroecosystem functions. However, for comprehensive view of agrobacterial community structure and SMR profile of the investigated soils, both temporal and spatial replicates of each soil type is needed. Additionally, further studies that examine the influence of plant type and agricultural management is required for a better conclusion.
REFERENCES
A. Ali, A. Esayas, and S. Beyene, “Characterizing soils of Delbo Wegene watershed, Wolaita Zone, Southern Ethiopia for planning appropriate land management,” J. Soil Sci. Environ. Manage. 1 (8), 184–199 (2010). https://doi.org/10.5897/JSSEM.9000018
A. Deressa, M. Yli-Halla, M. Mohamed, and L. Wogi, “Soil classification of humid Western Ethiopia: A transect study along a toposequence in Didessa watershed,” Catena 163, 184–195 (2018). https://doi.org/10.1016/j.catena.2017.12.020
A. Klindworth, E. Pruesse, T. Schweer, J. Peplies, C. Quast, F. O. Horn, and M. Glöckner, “Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies,” Nucleic Acids Res. 41 (1), 1–11 (2013). https://doi.org/10.1093/nar/gks808
Plant Growth Promoting Rhizobacteria for Agricultural Sustainability: From Theory to Practices, Ed. by A. Kumar and V. S. Meena (Springer-Verlag, New York, 2019), pp. 978–981. https://doi.org/10.1007/978-981-13-7553-8
Methods of Soil Analysis, Part 2: Chemical and Microbiological Properties, Agron. Monogr. no. 9.2, Ed. by A. L. Page, R. H. Miller, and D. R. Keene (America Society of Agronomy, Soil Science Society of America, Madison, WI, 1982), pp. 591–592.
A. Mehlich, Determination of P, Ca, Mg, K, Na and NH 4 (Department of Agriculture, Raleigh, NC, 1953), pp. 1–53.
A. Richter, D. O. Huallacháin, E. Doyle, N. Clipson, J. P. van Leeuwen, G. Heuvelink, and R. E. Creamer, “Linking diagnostic features to soil microbial biomass and respiration in agricultural grassland soil: a large-scale study in Ireland,” Eur. J. Soil Sci. 69 (3), 414–428 (2018). https://doi.org/10.1111/ejss.12551
A. Schöler, S. Jacquiod, G. Vestergaard, S. Schulz, and M. Schloter, “Analysis of soil microbial communities based on amplicon sequencing of marker genes,” Biol. Fertil. Soils 53, 485–489 (2017). https://doi.org/10.1007/s00vc4-017-1205-1
A. Sugiyama, Y. Ueda, T. Zushi, H. Takase, and K. Yazaki, “Changes in the bacterial community of soybean rhizospheres during growth in the field,” PLoS One 9 (6), e100709 (2014). https://doi.org/10.1371/journal.pone.0100709
A. Walkley and I. A. Black, “An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method,” Soil Sci. 37 (1), 29–38 (1934).
A. A. Aserse, L. A. Räsänen, F. Aseffa, A. Hailemariam, and K. Lindström, “Diversity of sporadic symbionts and nonsymbiotic endophytic bacteria isolated from nodules of woody, shrub, and food legumes in Ethiopia,” Appl. Microbiol. Biotechnol. 97, 10117–10134 (2013). https://doi.org/10.1007/s00253-013-5248-4
A. A. Ivanova, A. D. Zhelezova, T. I. Chernov, and S. N. Dedysh, “Linking ecology and systematics of acidobacteria: distinct habitat preferences of the Acidobacteria and Blastocatellia in tundra soils,” PLoS One 15 (3), e0230157 (2020). https://doi.org/10.1371/journal.pone.0230157
A. Hailemariam, “The effect of phosphate solubilizing fungus on the growth and yield of tef (Eragrostis tef) in phosphorous fixing soils,” in Proceedings of the Workshop on the 3rd Cycle Local Research Grant, Ethiopian Science & Technology Research Report (Addis Ababa, 1993), pp. 12–14.
B. Gebremedhin, Atsbi Wemberta Pilot Learning Site Diagnosis, Program Design and Atlas (Nairobi, 2004).
B. J. Tindall, R. Rosselló-Móra, H. J. Busse, W. Ludwig, and P. Kämpfer, “Notes on the characterization of prokaryote strains for taxonomic purposes,” Int. J. Syst. Evol. Microbiol. 60 (1), 249–266 (2010). https://doi.org/10.1099/ijs.0.016949-0
C. L. Lauber, M. Hamady, R. Knight, and N. Fierer, “Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale,” Appl. Environ. Microbiol. 75 (15), 5111–5120 (2009). https://doi.org/10.1128/AEM.00335-09
C. Quast, E. Pruesse, P. Yilmaz, J. Gerken, T. Schweer, P. Yarza, J. Peplies, and F. O. Glöckner, “The SILVA ribosomal RNA gene database project: improved data processing and web-based tools,” Nucleic Acids Res. 41 (1), 590–596 (2012). https://doi.org/10.1093/nar/gks1219
E. Högfors-Rönnholm, S. Christel, S. Engblom, and M. Dopson, “Indirect DNA extraction method suitable for acidic soil with high clay content,” MethodsX 5, 136–140 (2018). https://doi.org/10.1016/j.mex.2018.02.005
F. Cheng, P. Peng, J. Zhao, C. Yuan, Y. Zhong, C. Cheng, S. Cui, and S. Zhang, “Soil microbial biomass, basal respiration and enzyme activity of main forest types in the Qinling Mountains,” PLoS One 8 (6), e67353 (2013). https://doi.org/10.1371/journal.pone.0067353
F. Elias, D. Woyessa, and D. Muleta, “Phosphate solubilization potential of rhizosphere fungi isolated from plants in Jimma Zone, Southwest Ethiopia,” Int. J. Microbiol. 2016, 5472601 (2016). https://doi.org/10.1155/2016/5472601
F. Révész, P.A. Figueroa-Gonzalez, A. J. Probst, B. Kriszt, S. Banerjee, S. Szoboszlay, G. Maróti, and A. Táncsics, “Microaerobic conditions caused the overwhelming dominance of Acinetobacter spp. and the marginalization of Rhodococcus spp. in diesel fuel/crude oil mixture-amended enrichment cultures,” Arch. Microbiol. 202 (2), 329–342 (2020). https://doi.org/10.1007/s00203-019-01749-2
Guidelines for Soil Description, 4th ed. (UN Food and Agriculture Organization, Rome, 2006). ISBN 92-5-105521-1
G. Brhane, and K. Mekonen, “Estimating soil loss using Universal Soil Loss Equation (USLE) for soil conservation planning at Medego watershed, Northern Ethiopia,” J. Am. Sci. 5, 58–69 (2009).
G. Agegnehu, A. Ghizaw, and W. Sinebo, “Crop productivity and land-use efficiency of a teff/faba bean mixed cropping system in a tropical highland environment,” Expl. Agric. 42 (4), 495 (2006). https://doi.org/10.1017/S0014479706003863
H. Egnér, H. Riehem, and W. Domingo, “Untersuchungen über die chemische Bodenanalyse als Grundlage für die Beurteilung des Nährstoffzustandes der Böden II, Chemische Extraktionsmethoden zur Phosphor- und Kaliumbestimmung,” Kungl. Lantbrukshögsk. 26, 199–215 (1960).
Talaj-és Agrokémiai Vizsgálati Módszerkönyv, Vol. 2: A Talajok Fizikaikémiai és Kémiai Vizsgálati Módszerei, Ed. by I. Buzás (Mezőgazdasági Kiadó, Budapest, 1988), pp. 90–92, 96-98, 106–117, 175–177.
Talaj-és Agrokémiai Vizsgálati Módszerkönyv, Vol. 1: A Talaj Fizikai, Vízgazdálkodási és Ásványtani Vizsgálata, Ed. by I. Buzás (Mezőgazdasági Kiadó, Budapest, 1993), pp. 19, 37–41, 63.
ISO 16072:2002: Soil Quality—Laboratory Methods for Determination of Microbial Soil Respiration (International Organization for Standardization, Geneva, 2002).
IUSS Working Group WRB, World Reference Base for Soil Resources 2014, Update 2015, International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, World Soil Resources Reports No. 106 (UN Food and Agriculture Organization, Rome, 2015).
J. Aislabie and J. R. Deslippe, “Soil microbes and their contribution to soil services,” in Ecosystem Services in New Zealand: Conditions and Trends, Ed. by J. R. Dymond (Manaaki Whenua, Lincoln, 2013), pp. 143–161.
J. G. B. Leenaars, E. Eyasu, H. Wösten, M. R. González, B. Kempen, A. Ashenafi, and F. Brouwer, Major Soil-Landscape Resources of the Cascape Intervention Woredas, Ethiopia: Soil Information in Support to Scaling Op of Evidence-Based Best Practices in Agricultural Production (with Dataset) CASCAPE Working Paper Series no. OT_CP_2016_1 (Wageningen, 2016).
J. Hofman, L. Dušek, J. Klánová, J. Bezchlebová, and I. Holoubek, “Monitoring microbial biomass and respiration in different soils from the Czech Republic—a summary of results,” Environ. Int. 30 (1), 19–30 (2004). https://doi.org/10.1016/S0160-4120(03)00142-9
J. J. Kozich, S. L. Westcott, N. T. Baxter, S. K. Highlander, and P. D. Schloss, “Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform,” Appl. Environ. Microbiol. 79 (17), 5112–5120 (2013). https://doi.org/10.1128/AEM.01043-13
J. M. Craine, C. Morrow, and N. Fierer, “Microbial nitrogen limitation increases decomposition,” Ecology 88 (8), 2105–2113 (2007). https://doi.org/10.1890/06-1847.1
J. P. van Leeuwen, I. Djukic, J. Bloem, T. Lehtinen, L. Hemerik, P. C. de Ruiter, and G. J. Lair, “Effects of land use on soil microbial biomass, activity and community structure at different soil depths in the Danube floodplain,” Eur. J. Soil Biol. 79, 14–20 (2017). https://doi.org/10.1016/j.ejsobi.2017.02.001
J. Rousk, E. Bååth, P. C. Brookes, C. L. Lauber, C. Lozupone, J. G. Caporaso, R. Knight, and N. Fierer, “Soil bacterial and fungal communities across a pH gradient in an arable soil,” ISME J. 4 (10), 1340 (2010). https://doi.org/10.1038/ismej.2010.58
K. H. Habtemichial, B. B. Singh, and J. B. Aune, “Wheat response to N2 fixed by faba bean (Vicia faba L.) as affected by sulfur fertilization and rhizobial inoculation in semi-arid Northern Ethiopia,” J. Plant Nutr. Soil Sci. 170, 412–418 (2007). https://doi.org/10.1002/jpln.200625006
L. Albuquerque, L. França, F. A. Rainey, P. Schumann, M. F. Nobre, and M. S. da Costa, “Gaiellaoculta gen. nov., sp. nov., a novel representative of a deep branching phylogenetic lineage within the class Actinobacteria and proposal of Gaiellaceae fam. nov. and Gaiellales ord. nov,” Syst. Appl. Microbiol. 34, 595–559 (2011). https://doi.org/10.1016/j.syapm.2011.07.001
L. Liu, T. Zhang, F. S. Gilliam, P. Gundersen, W. Zhang, H. Chen, and J. Mo, “Interactive effects of nitrogen and phosphorus on soil microbial communities in a tropical forest,” PLoS One 8 (4), e61188 (2013). https://doi.org/10.1371/journal.pone.0061188
M. A. Gorbacheva, N. V. Melnikova, V. R. Chechetkin, Y. V. Kravatsky, and N. A. Tchurikov, “DNA sequencing and metagenomics of cultivated and uncultivated chernozems in Russia,” Geoderma Reg. 14, e00180 (2018). https://doi.org/10.1016/j.geodrs.2018.e00180
M. Ebrahimi, M. R. Sarikhani, A. A. S. Sinegani, A. Ahmadi, and S. Keesstra, “Estimating the soil respiration under different land uses using artificial neural network and linear regression models,”. Catena 174, 371–382 (2019). https://doi.org/10.1016/j.catena.2018.11.035
M. Lulu, B. Lemma, and A. Melese, “Soil organic carbon and nutrients in smallholding land uses in southern Ethiopia,” J. Plant Nutr. Soil Sci. 183 (1), 69–79 (2019). https://doi.org/10.1002/jpln.201900243
M. M. E. Andrea, T. E. A. Carolina, V. G. Anders, and R. G. Laura, “Relationship between soil physicochemical characteristics and nitrogen-fixing bacteria in agricultural soils of the Atlántico department, Colombia,” Soil Environ. 36 (2), 174–181 (2017). https://doi.org/10.25252/SE/17/51202
M. Rutgers, A. J. Schouten, J. Bloem, N. Van Eekeren, R. G. M. De Goede, G. A. J. M. Jagersop Akkerhuis, A. van der Wal, C. Mulder, L. Brussaard, and A. M. Breure, “Biological measurements in a nationwide soil monitoring network,” Eur. J. Soil Sci. 60 (5), 820–832 (2009). https://doi.org/10.1111/j.1365-2389.2009.01163.x
M. Tufa, A. Melese, and W. Tena, “Effects of land use types on selected soil physical and chemical properties: the case of Kuyu District, Ethiopia,” Eur. J. Soil Sci. 8 (2), 94–109 (2019). https://doi.org/10.18393/ejss.510744
N. A. Di Benedetto, M. R. Corbo, D. Campaniello, M. P. Cataldi, A. Bevilacqua, M. Sinigaglia, and Z. Flagella, “The role of plant growth promoting bacteria in improving nitrogen use efficiency for sustainable crop production: a focus on wheat,” AIMS Microbiol. 3 (3), 413 (2017). https://doi.org/10.3934/microbiol.2017.3.413
N. Fierer and R. B. Jackson, “The diversity and biogeography of soil bacterial communities,” Proc. Natl. Acad. Sci. U.S.A. 103 (3), 626–631 (2006). https://doi.org/10.1073/pnas.0507535103
N. Fierer, M. A. Bradford, and R. B. Jackson, “Toward an ecological classification of soil bacteria,” Ecology 88 (6), 1354–1364 (2007). https://doi.org/10.1890/05-1839
N. J. Fredriksson, M. Hermansson, and B. M. Wilén, “The choice of PCR primers has great impact on assessments of bacterial community diversity and dynamics in a wastewater treatment plant,” PLoS One 8 (10), e76431 (2013). https://doi.org/10.1371/journal.pone.0076431
P. D. Schloss, S. L. Westcott, T. Ryabin, J. R. Hall, M. Hartmann, E. B. Hollister, R. A. Lesniewski, B. B. Oakley, D. H. Parks, C. J. Robinson, and J. W. Sahl, “Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities,” Appl. Environ. Microbiol. 75 (23), 7537–7541 (2009). https://doi.org/10.1128/AEM.01541-09
P. Nannipieri, J. Ascher, M. T. Ceccherini, L. Landi, G. Pietramellara, and G. Renella, “Microbial diversity and soil functions,” Eur. J. Soil Sci. 68 (1), 12–26 (2003). https://doi.org/10.1111/ejss.4_12398
Q. Tian, T. Taniguchi, W. Y. Shi, G. Li, N. Yamanaka, and S. Du, “Land-use types and soil chemical properties influence soil microbial communities in the semi-arid Loess Plateau region in China,” Sci. Rep. 7, 45289 (2017). https://doi.org/10.1038/srep45289
Q. Zhang, J. Yang, R. T. Koide, T. Li, H. Yang, and J. Chu, “A meta-analysis of soil microbial biomass levels from established tree plantations over various land uses, climates and plant communities,” Catena 150, 256–260 (2017). https://doi.org/10.1016/j.catena.2016.11.028
R Development Core Team, R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, Vienna, 2017).
R. C. Edgar, B. J. Haas, J. C. Clemente, C. Quince, and R. Knight, “UCHIME improves sensitivity and speed of chimera detection,” Bioinformatics 27 (16), 2194–2200 (2011). https://doi.org/10.1093/bioinformatics/btr381
R. E. Creamer, D. Stone, P. Berry, and I. Kuiper, “Measuring respiration profiles of soil microbial communities across Europe using MicroResp™ method,” Appl. Soil Ecol. 97, 36–43 (2016). https://doi.org/10.1016/j.apsoil.2015.08.004
R. E. Creamer, R. P. O. Schulte, D. Stone, A. Gal, P. H. Krogh, G. L. Papa, P. Murray, G. Peres, B. Foerster, M. Rutgers, and J. P. Sousa, “Measuring basal soil respiration across Europe: Do incubation temperature and incubation period matter?” Ecol. Indic. 36, 409–418 (2014). https://doi.org/10.1016/j.ecolind.2013.08.015
R. Poretsky, R. L. M. Rodriguez, C. Luo, D. Tsementzi, and K. T. Konstantinidis, “Strengths and limitations of 16S rRNA gene amplicon sequencing in revealing temporal microbial community dynamics,” PLoS One 9 (4), e93827 (2014). https://doi.org/10.1371/journal.pone.0093827
R. T. Jones, “Comprehensive survey of soil Rhizobiales diversity using high-throughput DNA sequencing,” in Biological Nitrogen Fixation, Ed. by F. J. E. Bruijn (Wiley, Chichester, 2015), pp. 769–776. https://doi.org/10.1002/9781119053095.ch76
S. K. Hargreaves and K. S. Hofmockel, “Physiological shifts in the microbial community drive changes in enzyme activity in a perennial agroecosystem,” Biogeochemistry 117 (1), 67–79 (2014). https://doi.org/10.1007/s10533-013-9893-6
S. Kahsay and M. Mulugeta, “Determinants of rural household food insecurity in Laelay Maichew Woreda Tigray, Ethiopia,” Afr. J. Agric. Res. 2 (1), 106–112 (2014).
S. J. Joseph, P. Hugenholtz, P. Sangwan, C. A. Osborne, and P. H. Janssen, “Laboratory cultivation of widespread and previously uncultured soil bacteria,” Appl. Environ. Microbiol. 69 (12), 7210–7215 (2003). https://doi.org/10.1128/AEM.69.12.7210-7215.2003
S. Rughöft, M. Herrmann, C. S. Lazar, S. Cesarz, S. R. Levick, S. E. Trumbore, and K. Küsel, “Community composition and abundance of bacterial, archaeal and nitrifying populations in savanna soils on contrasting bedrock material in Kruger National Park, South Africa,” Front. Microbiol. 7, 1638 (2016). https://doi.org/10.3389/fmicb.2016.01638
T. Abera, D. Wegary, E. Semu, B. M. Msanya, T. Debele, and H. Kim, “Pedological characterization, fertility status and classification of the soils under maize production of Bako Tibe and Toke Kutaye districts of Western Showa, Ethiopia,” Ethiop. J. Appl. Sci. Technol. 7, 1–17 (2016).
T. J. Jackson, R. F. Ramaley, and W. G. Meinschein, “Thermomicrobium, a new genus of extremely thermophilic bacteria,” Int. J. Syst. Bacteriol. 23 (1), 28–36 (1973). https://doi.org/10.1099/00207713-23-1-28
T. Mori, X. Lu, R. Aoyagi, and J. Mo, “Reconsidering the phosphorus limitation of soil microbial activity in tropical forests,” Funct. Ecol. 32 (5), 1145–1154 (2018). https://doi.org/10.1111/1365-2435.13043
T. Teklay, A. Nordgren, and A. Malmer, “Soil respiration characteristics of tropical soils from agricultural and forestry land uses at Wondo Genet (Ethiopia) in response to C, N and P amendments,” Soil Biol. Biochem. 38 (1), 125–133 (2006). https://doi.org/10.1016/j.soilbio.2005.04.024
T. Yitbarek, S. Beyene, and K. Kibret, “Characterization and classification of soils of Abobo Area, Western Ethiopia,” Appl. Environ. Soil Sci. 2016, 4708235 (2016). https://doi.org/10.1155/2016/4708235
V. Kunin, A. Engelbrektson, H. Ochman, and P. Hugenholtz, “Wrinkles in the rare biosphere: pyrosequencing errors can lead to artificial inflation of diversity estimates,” Environ. Microbiol. 12 (1), 118–123 (2010). https://doi.org/10.1111/j.1462-2920.2009.02051.x
V. Nehra and M. Choudhary, “A review on plant growth promoting rhizobacteria acting as bioinoculants and their biological approach towards the production of sustainable agriculture,” J. Appl. Nat. Sci. 7 (1), 540–556 (2015). https://doi.org/10.31018/jans.v7i1.642
Ethiopia—Managing Water Resources to Maximize Sustainable Growth (World Bank, Washington, DC, 2006).
Y. T. Delelegn, W. Purahong, H. Sandén, B. Yitaferu, D. I. Godbold, and T. Wubet, “Transition of Ethiopian highland forests to agriculture-dominated landscapes shifts the soil microbial community composition,” BMC Ecol. 18 (1), 58 (2018). https://doi.org/10.1186/s12898-018-0214-8
Y. X. Chen, L. Zou, P. Penttinen, Q. Chen, Q. Q. Li, C.Q. Wang, and K. W. Xu, “Faba bean (Vicia faba L.) nodulating rhizobia in Panxi, China, are diverse at species, plant growth promoting ability, and symbiosis related gene levels,” Front. Microbiol. 9, 1338 (2018). https://doi.org/10.3389/fmicb.2018.01338
Z. Belay, M. Vestberg, and F. Assefa, “Diversity and abundance of arbuscular mycorrhizal fungi associated with acacia trees from different land use systems in Ethiopia,” Afr. J. Microbiol. Res. 7 (48), 5503–5515 (2013). https://doi.org/10.5897/AJMR2013.6115
Z. S. L. Foster, T. J. Sharpton, and N. J. Grünwald, “Metacoder: An R package for visualization and manipulation of community taxonomic diversity data,” PLoS Comput. Biol. 13 (2), e1005404 (2017). https://doi.org/10.1371/journal.pcbi.1005404
Z. Tsegaye, B. Gizaw, G. Tefera, A. Feleke, S. Chaniyalew, T. Alemu, and F. Assefa, “Isolation and biochemical characterization of Plant Growth Promoting (PGP) bacteria colonizing the rhizosphere of Tef crop during the seedling stage,” Biomed. J. Sci. Tech. Res. 14 (2), 1586–1597 (2019). https://doi.org/10.29328/journal.jpsp.1001027
FUNDING
This work was funded by Stipendium Hungaricum Scholarship Program (SHE-935-1/2016 and it was supported by the Ministry of Innovation and Technology within the framework of the Thematic Excellence Program 2020, Institutional Excellence Sub-Program (TKP2020-IKA-12) in the topic of water-related researches of Hungarian University of Agriculture and Life Sciences. Mátyás Cserháti was supported by the ÚNKP-18-4/12 New National Excellence Program of the Ministry of Human Capacities. We are grateful to Mr. Mehretab Haileselassie and other Tigray Agricultural Research Institute members for their help in the fieldwork.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The authors declare that they have no conflict of interest.
Supplementary Information
Rights and permissions
About this article

Cite this article
Weldmichael, T.G., Márton, D., Simon, B. et al. Bacterial Community Characterization and Microbial Respiration of Selected Arable Soils of Ethiopia. Eurasian Soil Sc. 54, 1921–1934 (2021). https://doi.org/10.1134/S1064229321120140
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1064229321120140