
Abstract
Burdurina selandinica Sirel from Paleocene carbonates of the Golbui section of Central Iran is reported and discussed. The stratigraphic position of foregoing succession is consistent with the previous studies depending on the faunal assemblage of the Selandian–early Thanetian, namely Sistanites iranicus Rahaghi, Anatoliella sp., Haymanella paleocenica Sirel, Coscinospira prima Schlagintweit et al., Miscellanea cf. yvettae Leppig, Elazigina dienii (Hottinger), Idalina sinjarica Grimsdale, Pachycolumella acuta Septfontaine et al., Pachycolumella elongata Septfontaine et al., Rotospirella conica (Smouth). After the latest reorganization on Shallow Benthic Zones (SBZs) of the Paleocene, two stages correspond to SBZ3. Three predominant microfacies types including coral-coralline algal framestone, bioclastic foraminiferal-peloidal grainstone-packstone and coral floatstone were recognized in the studied section. The microfacies gradients within the succession and paleoenvironmental analysis suggest deposition in a shallow-marine ramp environment that is located in a global carbonate platform stage I from the middle paleolatitude.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
INTRODUCTION
The early Paleogene period witnessed a large-scale proliferation in coralgal reefs and larger benthic foraminifera in various carbonate platforms of the Neo-Tethyan domain. Shallow-marine carbonate successions pertinent to the Paleocene have been subject to various case studies based on the sedimentological and paleontological observations, especially from Europe, the Peri-Mediterranean and Asia (e.g., Sirel, 1998, 2018; Sarkar, 2020; Scheibner et al., 2007; Sirel and Deveciler, 2018; Zhang et al., 2013). However, widespread Paleocene outcrops across the Middle East remain understudied in terms of detailed biostratigraphic and paleoenvironmental evaluations. In this context, few benthic foraminifera-centric studies from Iran were carried out (e.g., Rahaghi, 1978, 1983; Schlagintweit and Rashidi, 2019; Septfontaine et al., 2019; Schlagintweit et al., 2020, 2021). However, Burdurina Sirel has not been reported till date from any Paleocene sediments of Iran. Considering the paleogeographic implications of the genus Burdurina discussed herein and its interesting distribution pattern over the low-middle paleolatitudes, especially in the Middle East, a detailed investigation on this genus is of considerable significance. In addition to the taxonomic and paleogeographic evaluation of Burdurina, the current paper also focusses on presenting a summary of the microfacies analysis of the studied carbonate succession with potential contribution to the paleoenvironmental reconstruction of the Central Iran.
GEOLOGICAL SETTING
The Iranian plateau is subdivided into nine sedimentary-structural provinces, being part of the largest mountain belt of the Alpine-Himalayan system. These provinces are: (1) the southern Caspian Sea basin, (2) Alborz in NW Iran, (3) Kopet-Dagh in NE Iran, (4) the Central Iranian block or Cimmeria, consisting of three major old continental blocks (from E to W): Lut, Tabas, and Yazd, separated by major faults (e.g., Alavi, 1991) and similar crust to the NW that is mostly buried beneath Cenozoic deposits, (5) Sanandaj-Sirjan zone, (6) Urumieh-Dokhtar (Sahand-Bazman) magmatic arc, (7) The Zagros Fold-Thrust Belt, (8) Eastern Iranian suture zone; and (9) Makran, each province having experienced distinct tectonic and sedimentary histories (Stöcklin, 1968, Shafaii-Moghadam and Stern, 2014) (Fig. 1a). Central Iran is one of the most important and complicated structural zones in Iran, which is located as a triangle in the middle of Iran within the Alpine-Himalayan orogenic system (Fig. 1b). On the other hand, Central Iran features the Alborz Mountains in the north, Birjand-Sistan ocean zone in the east, and Sanandaj-Sirjan zone in the south-southwest (Fig. 1a). Most tectonic studies for the closure of Neo-Tethys show that an active margin formed along southern Eurasia from late Mesozoic through the Cenozoic time (e.g., Berberian and King, 1981; Shafaii-Moghadam and Stern, 2014). Two main Cenozoic collisional phases are recognized in central and NE Iran including Neogene collision between Iran and Arabia plates and the Paleocene collision between central Iran and Eurasia to close the Sabzevar basin (Shafaii-Moghadam et al., 2015). In context of paleogeography, central Iran region was in the northern Neo-Tethyan margin during the Paleogene. Paleocene carbonate successions in the central and eastern Iran are scarce, wherein not defined as a formal unit so far. However, the Palang Formation was discussed by Tirrul et al. (1983) and recently also examined based upon some taxa of larger benthic foraminifera (see Schlagintweit et al., 2020). In general, the Iranian plateau was strongly influenced by the Laramid orogeny and local tectonic activity of different provinces mentioned above such as the Alborz and central Iran regions, which are characterized by conglomerates and red sandstone units as Fajan Formation and Kerman conglomerate in the Kopet-Dagh with modest outcrops of the Pesteligh and Chehel-Kaman formations (Fig. 1b). Likewise, Paleocene successions are known with different lithostratigraphic units by deep marine sediments as the Pabdeh Formation, whilst the Sachun, Jahrum and Taleh-Zang formations are representative of shallow marine benthic foraminifera deposits in the Zagros zone (Fig. 2), however, the extensive outcrops of deep marine silisiclastic units are only exposed as the Amiran Formation (upper Maastrichtian–Paleocene) in Lurestan basin of the Zagros fold-trust belt (Fig. 1b).

(a) General map of Iran showing nine geologic provinces (adapted from Stöcklin, 1968 and Shafaii Moghadam and Stern, 2014); (b) Tethys mountain ranges (simplified from Okay, 1989; Özcan et al., 2016; Hadi et al., 2019) and location of the studied region in the central Iran zone.

(a) Geographic map and location of the studied outcrop (geological map of Kadkan region, 1 : 100 000, from Naderi-Mighan and Torshizian, 1999); (b) outcrop view of the Paleocene shallow marine succession in the Golbui section.

Plate 1 . All figures from Selandian–Thanetian limestone of the Golbui section. (1–10) Burdurina selandinica Sirel: (1) axial section (GB/25/6); (2) almost equatorial section (GB/15/2b); (3) almost equatorial section showing septal flap (sfl) (GB/23B/3); (4) axial section (GB/12/5); (5) oblique section showing funnels (f) and pustules (pu) (GB/12/1); (6) almost equatorial section (GB/23B/2); (7) axial and equatorial section showing enveloping canal sytem (ecs), multispiral (ms), pillars and funnels (GB/23B/1); (8) transvers section to equatorial plan showing pustules (pu) (GB/25/9); (9) almost axial section showing pores, pillars and funnels (GB/14/9); (10) tangential section showing pustules (pu) (GB/23/3). (11–16) Miscellanea cf. yvettae Lepping: (11) axial section of a megalospheric form (GB/37/2); (12) axial section (GB/52/1); (13) axial section of a possible microspheric form (GB/74/1); (14) incomplete equatorial section (GB/41/1); (15) incomplete equatorial section of a megalospheric form (GB/47/3); (16) axial section (GB/37/1). Abbreviations: (ch) chamber, (f) funnel, (icf) intercameral foramen, (isc) intraseptal canal, (msc) marginal sutural canal, (pi) pillar, (pr) prothochonch, (sc) spiral canal.

All figures from Selandian-Thanetian limestone of the Golbui section. (1–2) Elazigina dienii Sirel: (1) axial section (GB/74/3), (2) axial section (GB/47/2); (3–5) Rotospirella conica: (3) axial section (GB/47/1), (4) axial section (GB/24B/3), (5) almost equatorial section (GB/74/2b-r); (6–11) Idalina sinjarica Grimsdale: (6) centred longitudinal section (GB/71/1b), (7) oblique section (GB/52/8), (8) oblique longitudinal section showing trematophore (tr) (GB/71/1), (9) oblique section (GB/52/10), (10) centred transversal section (GB/50/1), (11) longitudinal section with Pachycolumella acuta (GB/34/1); (12) Globotextularinid type, vertical section (GB/4/1); (13, 14) Pachycolumella elongata Septfontain et al.: (13) axial section (GB/74/2a), (14) oblique axial section (GB/34/3); (15) Pachycolumella acuta Septfontain et al., vertical section (GB/74/2b).

All figures from Selandian–Thanetian limestone of the Golbui section. (1) Keramospherid genus, almost centred section (GB/36/1); (2–4) Sistanites iranicus Rahaghi: (2) axial section (GB/13/2), (3) axial section (GB/33), (4) axial section (GB/13/3); (5) Coscinospira prima Schlagintweit et al., incomplete longitudinal section with miliolids (GB/41/1), (6) incomplete longitudinal section with Miscellanea cf. yvettae and Haymanella sp. (GB/74/1), (7) Longitudinal section (GB/71/4); (8) Anatoliella sp., oblique section (GB/18/1); (9) Haymanella paleocenica Sirel, longitudinal section (GB/24B/5); (10) Spirolinid type, centered longitudinal section (GB/71/2).
MATERIAL AND METHODS
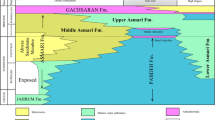
Our analysis in the north of Torbat-e-Heydarieh region (Central Iran) is focussed on the upper Paleocene shallow-water limestones that occur within the Sabzevar-Torbate-e-Heydarieh magmatic belt (Fig. 2a). The Golbui section is ~210 m thick and was measured ~5 km southeast of Golbui-bala village, which is ~35 km south of the Neyshabur town (Fig. 2a). It is located in 65 km southwest of Mashhad town (Fig. 2a). The section is situated in the sheet-7761 (coordinates: 35°44′56′′ N; 58°51′22′′ E). White to light gray limestones are often thick-bedded to massive (Fig. 2b). This thick Paleocene interval can be one of the most complete shallow-marine carbonate sequences in the central Iran region, although the lower and upper boundaries are covered (Fig. 3). However, exposed sedimentary sequences of the Kuh-e-Chah Torsh section in the central Iran, Yazd block was also studied based upon the rich assemblages of the Paleocene large benthic foraminifera previously reported by Schlagintweit and Rashidi (2019). In total, 76 rock samples from the indurated limestone with 1–2 m sampling interval were collected and 140 randomly oriented thin-sections were prepared for the micropaleontological study. The thin sections (2.5 × 7.5 cm) were studied and digitally photographed under transmitted-light and binocular microscopes. Loeblich and Tappan (1987), Leppig (1988), Hottinger (2006, 2014), Leppig and Langer (2015), Holzmann and Pawlowski (2017) and Sirel (2018) are followed for the suprageneric classification and morphological descriptions of the benthic foraminifera. Dunham (1992), Embry and Klovan (1972) and Flügel (2004) are followed in describing the major microfacies types (MFTs). The shallow benthic zones (SBZ) of the foraminiferal species follow Serra-Kiel et al. (2020).

Stratigraphic distribution of benthic foraminifera species in the Golbui section, Central Iran.
SYSTEMATIC PALEONTOLOGY
B. selandinica Sirel is described and discussed in detail. Also, M. cf. yvettae and E. dienii are only examined in terms of previous studies and their biostratigraphic position in the Golbui section. Other associated fauna is given together with the stratigraphic range of foregoing species.
ORDER ROTALIIDA DELAGE ET HÉROURAD, 1896
>FAMILY MISCELLANEIDAE SIGAL IN PIVETEU, 1952
SUBFAMILY MISCELLANEINAE SIGAL IN PREVETEAU, 1952
Genus Miscellanea Pfender, 1935
Type species: Nummulites miscella d’ Archiac et Haime, 1853
Miscellanea cf. yvettae Leppig, 1988
Pl. 1, figs. 11–16
Miscellanea yvettae: Leppig, 1988, p. 702, pl. 1, fig. 2; pl. 2, fig. 2; pl. 3, fig. 2; pl. 6, figs. 1–9.

Structural elements of Burdurina Sirel (type species: Miscellanea ? globularis Rahaghi (in Sirel, 1998)). (a) Almost equatorial of possible microspheric form, section showing chambers, intraseptal canals and pustules (GB/14/5); (b) axial section of megalospheric form, showing multi spiral, funnels and pores (GB/10/3); (c) equatorial section of megalospheric form, showing intraseptal canals, pores and spheric protochonch (GB/10/4); (d) axial section, showing pillars, funnels and pores (GB/12/9); (e) axial section of megalospheric form, showing pillars, funnels and multi spiral (GB/19/1); (f) Almost equatorial section, showing spiral canal and multi spiral (GB/10/9). Abbreviations: (ch) chamber, (ecs) enveloping canal system, (f) funnel, (fr) foramen, (isc) intraseptal canal, (ms) multi spiral, (p) pore, (pi) pillar, (pr) prothochonch, (pu) pustule, (sep) septum, (sfl) septal flap.

Comparison chart of Miscellanea, Miscellanites and Burdurina.

Microfacies components of the Golbui section. (a, b) Coral-coralline algal framestone, note Coral (CR), coralline algal (C), Sample No. GB1; (c, d) bioclastic foraminiferal-peloidal grainstone-packstone, Miliolid (M), Miscellanea (MS), Sample No. GB11/12; (e, f) coral floatstone, note Coral (CR), Bivalve (B), Miliolid (M), Sample No. 68/76.

Paleogeographic sketch map near the Cretaceous/Cenozoic boundary (about 70 Ma) showing the location of the study area and the compared sectors (after Scheibner and Speijer, 2008; Martín-Martín et al., 2020).

Palaeogeographic distribution of Burdurina selandinica in the Neo-Tethys Ocean during the early Paleocene, map modified from Smith et al. (1994). (1) Oman, from Serra-Kiel et al. (2016), (2) Iran, Schlagintweit and Rashidi (2019), (3) Iran, (present study), (4) Turkey, Sirel (2018), (5) Italy, Benedetti et al. (2018).
Miscellanea juliettae Leppig: Sirel, 1997, pp. 78, 81, pl. 1, figs. 1–14; pl. 2, figs. 1–18.
Miscellanea yvettae Leppig: Hottinger, 2009, pp. 5–6, pl. 8, figs. 4–6, 9–11; pl. 9, figs. 1–12.
Miscellanea julietta: Lepping and Langer, 2015, pp. 227–228, 230, pl. 1, figs. 1–16.
Miscellanea yvettae Leppig: Sirel, 2018, pp. 80–81, pl. 18, figs. 1–13.
Miscellanea miscella (d’Archiac et Haime): Schlagintweit et al., 2020, fig. 3D.
Miscellanea yvettae Leppig: Serra-Kiel et al., 2020, pp. 46, 48, figs. 24A–C.
Miscellanea yvettae Leppig: Vršič et al., 2021, pp. 10–11, fig. 11, figs. 1–9.
Remarks. Our samples resemble early ontogeny of M. yvettae by size, test shape and number of whorls (Pl. 1, figs. 11–16). However, due to the inadequate materials, absence of characteristic box-shaped microspheric generations and the possibility of being a different taxon the specimens of the Golbui section left open. Miscellanea sp. of the Selandian in Benedetti et al. (2018, Fig. 3B) and Di Carlo et al. (2010, p. 70, pl. 2, figs. 1–16) is differentiated with swollen test shape from the Golbui samples.
With regard to this matter, we would like to introduce the discussions of M. yvettae and M. juliettae by the following authors: Leppig (1988) made a detailed structural and taxonomic analysis of miscellaneids and created three taxa, namely Miscellanea juliettae pfenderea, M. juliettae villatea and M. yvettae. However Sirel (1997) realized the similarities between M. juliettae villattae, M. juliettae pfendarae and M. yvettae in Leppig (1988) while Hottinger (2009) expressed the problem depending on the taxonomic definition of M. juliettae pfenderea and M. juliettae villatea and remarked the similarities (Hottinger, 2009, p. 6) just as Sirel (1997). Owing to these cases M. juliettae pfenderea and M. juliettae villatea were amended to M. juliettae by Leppig and Langer (2015), but the subspecies were transferred into M. yvettae by Sirel (2018), due to the similar characteristics of the mentioned specimens. Serra-Kiel et al. (2020) defined the aforementioned species as two separate species, however Vršič et al. (2021) decided to follow M. yvettae instead of M. juliettae by pointing out these confusions.
Stratigraphic range. Miscellanea cf. yvettae is recorded with Elazigina dienii (Pl. 2, figs. 1–2), Idalina sinjarica (Pl. 2, figs. 6–10), Pachycolumella acuta (Pl. 2, figs. 11, 15), Pachycolumella elongata (Pl. 2, figs. 13–14), Sistanites iranicus, Haymanella paleocenica (Pl. 3, fig. 8), Coscinospira prima (Pl. 3, figs. 5–6, 9) and Rotospirella conica (Pl. 2, figs. 3–5) in the early late Selandian–early Thanetian (SBZ3; after Serra-Kiel et al., 2020) of the Golbui section.
SUBFAMILY MISCELLANITINAE HOTTINGER, 2009
Genus Burdurina Sirel, 2018
Type species: Miscellanea? globularis Rahaghi, in Sirel, 1998.
Description. This definition as regarded Burdurina Sirel is based on the sections of Sirel (1998, 2018) and studied samples from the Golbui section, Iran.
The test is globular to subglobular in shape (Figs. 4b, 4d, 4e; Pl. 1, figs. 1, 4, 5, 7). Planispirally coiled whorls are followed by a kind of multispire (Figs. 4b–4c, 4f; pl. 1, figs. 1, 3, 4; Sirel, 1998, pl. 58, figs. 1, 4, 8; Sirel, 2018, pl. 19, figs. 1, 5, 6), that is likely curled over the penultimatel whorl of the spiral stage (Figs. 4d–4e; Pl. 1, figs. 7, 9; Sirel, 2018, pl. 19, figs. 1, 6). A megalospheric specimen in Sirel (1998, pl. 58, fig. 4) clearly shows the duplication of spire can start during the early ontogeny. The hyaline calcareous wall has numerous radial canals leading to pores settled between prominent pustules (Figs. 4b, 4c, 4e; pl. 1, figs. 8, 9, 10). Well oriented axial sections show pillars and funnels present on both side of the test (Figs. 4b, 4d, 4e; pl. 1, figs. 1, 5, 7). Thin septa with simple intra-septal canals (Figs. 4a, 4c, 4f) are covered by septal flaps inside the chambers (Figs. 4a, 4c, 4f; pl. 1, figs. 3, 5–7). The characteristic of foramen is uncertain (Fig. 4c), however one oblique thin section in (Sirel, 2018, pl. 19, fig. 6) has multiple foramen at the base of the septal face. Enveloping canal system may be observed in some oriented thin sections (Fig. 4f; pl. 1, figs. 7, 10).
Remarks. Miscellanea globularis was described as a new species from the Paleocene of the Shiraz area (Iran) by Rahaghi (1978, p. 61, pl. 12, figs. 10–20). Afterwards Rahaghi (1983, p. 61, pl. 42, figs. 1–7) reported this species again with few additional figures from the upper Paleocene of Shiraz and Sistan regions from Iran.
Later on Sirel (1998, pl. 58, figs. 1–11; 2018, pl. 19, figs. 1–7) defined the specimens from Selandian of Yarışlı/Burdur area (south of Turkey) as M. ? globularis by pointing out the some differences in generic characteristics with Miscellanea Pfender. Depending on the samples collecting from Haymana (Turkey), Hottinger (2009) transferred M. globularis Rahaghi to Miscellanites Hottinger, however Sirel (2018, p. 83–84, pl. 19, figs. 1–7) described the samples in (Sirel, 1998) as Burdurina selandinica and accepted Miscellanites globularis as junior synonym of B. selandinica. Miscellanea Pfender is characterized by having simple planispirally enrolled test with single foramen, septal-umblical flaps (umblical plate), complicated canal system such as spiral, intraseptal, fan shaped marginal, sutural canals, funnels (verticle canals) between coarse pillars (Lepping, 1988; Hottinger, 2009) and enveloping canal system (Hottinger, 2009, pl. 4, fig. 8; pl. 5, figs. 8–10). In this context, Miscellanites Hottinger (type species: Miscellanea iranica Rahaghi) is basically differentiated from Miscellanea for having multiple intercameral foraminina (Hottinger, 2009, pl. 12, fig. 6; pl. 13, figs. 7, 12) and devoid of spiral and marginal-sutural canals, nevertheless bifurcate intraseptal canals are evident in some figures of Hottinger (2009, pl. 12, fig. 4; pl. 13, figs. 5, 8–12). However, generic characteristics of Miscellanites globularis by Hottinger (2009) does not complately resemble to the type species of Miscellanites in respect of multiple coiling chambers (Hottinger, 2009, pl. 22, figs. 3, 8–10, 16) and simple interaseptal canal. Here described specimens in (Figs. 4a–4f, pl. 1, figs. 1–10) and that of Hottinger (2009, pl. 22, figs. 1–24; pl. 23, figs. 7–13) are identical with B. selandinica Sirel, through the definition in Sirel (2018). On the other hand, Burdurina has septal flap, multiple foramina, pillars and funnels, but may not have a streptospiral arrangement of chambers at early stage unlike in (Sirel, 2018) (Fig. 5). Also, Miscellanitinae subfamily was adopted for the presence of multiple intercameral foramina.
Burdurina selandinica Sirel, 2018
Figs. 4a–4d; Pl. 1, figs. 1–10
Miscellanea ? globularis Rahaghi: Sirel, 1998, p. 97, pl. 58, figs. 1–12.
Miscellanites globularis (Rahaghi): Hottinger, 2009, p. 10, pl. 12, fig. 5; pl. 22, figs. 1–24; pl. 23, figs. 7–13.
Unidentified miscellanid genus (probably n. gen.): Sirel, 2015, pl. 8, figs. 1–7.
Miscellanites globularis (Rahaghi): Serra-Kiel et al., 2016, p. 331, fig. 8.28.
Burdurina selandinica: Sirel, 2018, p. 83–84, pl. 19, figs. 1–7.
Miscellanea sp.: Benedetti et al., 2018, p. 78, figs. 3.D–E.
Miscellanea ? globularis Rahaghi: Acar, 2019, pl. 13, figs. 1–9.
Miscellanites minitus (Rahaghi): Schlagintweit and Rashidi, 2019, p. 314–315, pl. 6, figs. 14, 16–17.
Miscellanites globularis (Rahaghi): Serra-Kiel et al., 2020, p. 48, Fig. 24N.
Description. The globular to sub-globular small test (Figs. 4b, 4d, 4e; Pl. 1, figs. 1, 4, 5, 7, 9) has a diameter of 0.72 to 1.18 mm and thickness of 0.71 to 0.97 mm. The ratio of diameter/thickness is 1.09–1.175. The subspherical megalosphere measures 0.058 to 0.1 mm in diameter (Figs. 4c, 4e). Multiple spiral occurs right after a cyclical arranged chambers and its ultimate part covers the penultimate whorl (Figs. 4b–4f; Pl. 1, figs. 1–10).
The larger specimens with larger chambers as in (Fig. 4a) suggest the existence of two generation.
Stratigraphic range. B. selandinica were reported by Sirel (2018) from the Selandian of Haymana-Polatlı, Cide, Gölköy-Ordu and Yarışlı-Burdur localities (Turkey) associating with Laffiteina mengaudi, L. erki, L. mengaudi trochoidea, L. bibensis, Bolkarina aksarayensis, Akbarina yarisliensis, Pseudolacazina oeztemuueri, P. donattae, Pseudocuvilierina sireli, Globoflarina sphaeroidea, Sistanites iranica and Idalina sinjarica.
It is associated with Pachycolumella acuta (Pl. 2, figs. 11, 15), Sistanites iranica (Pl. 3, figs. 2–4), Rotospirella conica (Pl. 2, figs. 3–5), Anatoliella sp. (Pl. 3, fig. 7), Haymanella paleocenica (Pl. 3, fig. 8) and Coskinospira prima (Pl. 3, figs. 5–6, 9) in the Golbui section. Since P. acuta and C. prima were attributed to the Selandian–Thanetian of the similar successions from Iran (Septfontain et al., 2019; Schlagintweit et al., 2020), B. selandinica is accepted in late Selandian–early Thanetian (SBZ3) limestone of the Golbui section.
FAMILY ROTALIIDAE EHRENBERG, 1839
Genus Elazigina Sirel, 2012
Type species: Kathina subsphaerica Sirel, 1972
Elazigina dienii (Hottinger, 2014)
Pl. 2, figs. 1–2
Plumokathina dienii: Hottinger, 2014, p. 110, Figs. 6.1A–L; pl. 6.8, figs. 1–21.
Elazigina dienii (Hottinger): Benedetti et al., 2018, p. 79, Figs. 5A–5G.
Elazigina cf. dienii (Hottinger): Schlagintweit and Rashidi, 2019, p. 314, pl. 6, figs. 10–11.
Elazigina aff. lenticula (Hottinger): Schlagintweit et al., 2020, Fig. 3E.
Elazigina dienii (Hottinger): Serra-Kiel et al., 2020, p. 38–39, Figs. 20Y’–B”.
Elazigina harabekayisensis Sirel: Benedetti et al., 2021, Fig. 2e.
Remarks. Plumokathina dienii was first described by Hottinger (2014), then it was declared as the junior synonym of Elazingina dienii by Benedetti et al. (2018). Only two specimens from the Golbui section (Pl. 2, figs. 1, 2) resemble the specimens of Hottinger (2014) in terms of biconvex lenticular test and slightly up curved pointed periphery. Although E. harabekayisensis Sirel resembles E. dienii at first glance, E. dienii differs with possessing smaller diameter, less swollen on both dorsal and ventral side and distinguished pointed periphery. Also E. subspherica (Sirel) has a larger test with lamellar wall structure and E. lenticula (Hottinger) has a subglobular test as compared with E. dienii.
Stratigraphic range. E. dienii was first defined in Selandian SBZ2 (according to Serra-Kiel et al., 1998) from Sardinia (Italy) without an associated fauna by Hottinger (2014). Then it was reported from Italy by Benedetti et al. (2018) associated with Cuvillierina cf. sireli, Miscellanea sp. and Ornatorotalia sp. in SBZ2 of the Selandian of Serra-Kiel et al. (1998) whereas Serra-Kiel et al. (2020) considered it as a marker of SBZ2 of the Danian. Schlagintweit et al. (2020) and Benedetti et al. (2021) identified one identical specimen similar to E. dienii with a Selandian–early Thanetian (SBZ3) fauna in Qorban member, Zagros area, Iran.
We consider this species in SBZ3 after Serra-Kiel et al. (2020) (late Selandian–early Thanetian) in the Golbui section of Iran with the foregoing associated benthic foraminifera of M. cf. yvettae. A possible migration in time span between SBZ2 and SBZ3 could have been the cause of the age difference with the Western Mediterranean and Middle East. Thus the biostratigraphic range of E. dienii may be wider throughtout the Tethys region.
MICROFACIES AND PALEOENVIRONMENT
Three major microfacies types (MFTs) were recognized in the presently studied succession: (1) coral-coralline algal framestone; (2) bioclastic foraminiferal-peloidal grainstone-packstone, and (3) coral floatstone.
Coral-coralline algal framestone (MFT 1) (Figs. 6a, 6b). This poorly sorted microfacies type with a thickness of 82 m is characterized by the dominance of branching corals and crustose coralline algae that represent primary framework builders. The scleractinians here display moderate level of preservation, mostly composed of low generic diversity (predominantly of families Faviidae and Poritidae). The encrusting corallines are mainly represented by the melobesioids and sporolithaceans. Abundant sedimentary matrix is observed within the framestone. In the micritic matrix, smaller benthic foraminifera and fragments of bivalves, gastropods, algae and microbial intraclasts and peloids are commonly dispersed in spaces between the corals and corallines. Coral heads commonly show evidences of intense organic boring by the sponges, worms and bivalves. Tabular corals are rare, forming only localized small-scale monotypic aggregations.
Bioclastic foraminiferal-peloidal grainstone-packstone (MFT 2) (Figs. 6c, 6d). This type has a thickness of 78 m and is dominated by moderately sorted and densely packed fabric, with miscellaneid and miliolid foraminifera as the major biotic facies components. Peloids and debris of corals, algae, bivalves, gastropods, echinoderms, and other benthic foraminifera are abundant with exotic micritic infillings in some microfossils. The MFT shows the presence of localized micrite and also features bio- and oomicritic intraclasts. Such foraminiferal assemblages with miscellaneids as one of the major components were also described from other Tethyan localities like Taleh Zang Formation in the Zagros Mountains (Bagherpour and Vaziri, 2012) and Indus Basin, Pakistan (Afzal et al., 2010). Miscellaneids were ascribed mainly in the near-reef zone of the back reef (Ghose, 1977) and also described from the inner ramp together with porcelaneous taxa (Zamagni et al., 2008).
Coral floatstone (MFT 3) (Figs. 6e, 6f). This type contains poorly sorted and dispersed/loosely packed, complete or fragmented corals occasionally encrusted by corallines and sessile foraminifera with a thickness of 40 m. Coral colonies in growth position are present showing hemispherical to globose shapes (10–60 cm height and 10–70 cm width). The MFT is characterized by a foraminiferal-algal wackestone to packstone matrix. Rare gastropods, green algae, bivalves, and red algae are the secondary components. The proportion of bioerosion and micritization due to the activity of microborers is much lower compared to the other MFTs. Fossils within this microfacies are relatively compact and locally in situ. The mud-supported microfacies is inferred to have developed in low hydrodynamic energy environment.
DEPOSITIONAL ENVIRONMENT AND PLATFORM STAGES
In the Golbui section, an obvious deepening or shallowing trend pertaining to the overall Paleocene environments is not inferred. This is due to the evaluated deposits representing only a part of the Selandian and Thanetian stages without confirmed information regarding the lower boundary of SBZ2 and upper boundary of SBZ3. Nevertheless, the microfacies analysis indicates a trend of decreasing hydrodynamic energy toward the distal part of the succession, with ascending abundance of the massive and branching corals. The microfacies components and overall biotic assemblages correlate the studied succession to an inner ramp environment.
The presently studied Paleocene deposits of the Golbui section (central Iran region) were at approximately 27°–28° N paleolatitude. The site was dominated by in situ corals probably of patch-reef type along with benthic foraminifera and coralline algae corresponding to the platform stage I (Scheibner and Speijer, 2008). The region shares close paleogeographic proximity to the northwestern Arabian platform, where both corals and benthic foraminifera were preponderant (Höntzsch et al., 2013, Fig. 6) in the sections belonging to SBZ1–SBZ3 (Fig. 7). As mentioned by Scheibner and Speijer (2008) the biogenetic components of platform stage I were uncertain in Turkey but recently Consorti and Köroğlu (2019) reported the presence of coralgal assemblages as “Coral-algal patch-reef facies” in the Paleocene successions from the Eastern Pontides area. Therefore, by means of the current study for the first time the deposits of the platform stage I have been confirmed in the carbonate platform environments from the middle latitudes from Iran, which was also a part of convergent zone in the Paleocene–Eocene period.
BIOSTRATIGRAPHIC AND PALEOBIOGEOGRAPHIC IMPLICATIONS
Recently, Selandian–Thanetian successions of Iran were carried out by Benedetti et al. (2018), Schlagintweit and Rashidi (2019), Septfontain et al. (2019) and Schlagintweit et al. (2020) from various localities of Zagros area. Although the associated fauna of the Golbui section were described in the foregoing studies, B. selandinica was defined in Paleocene limestone of Iran very first time.
B. selandinica was first introduced by Sirel (2018) from various Selandian localities of Turkey based upon the type species Miscellanea ? globularis (Rahaghi) in Sirel (1998). Later on it was reported under different names from SBZ2-3 (Serra-Kiel et al., 1998) in Mediterranean and Middle East by Hottinger (2009), Serra-Kiel et al. (2016), Acar (2019), Schlagintweit and Rashidi (2019) and Benedetti et al. (2018) (Fig. 8). Now it represents only SBZ3 with together its associated fauna after Serra-Kiel et al. (2020), which made remarkable changes in Shallow Benthic Zones of Paleocene depending on the latest study of Schmitz et al. (2011), Hottinger (2014) and the benthic foraminiferal association from various localities of Pyrenean zone. As a general concept of the study, SBZ1 was accepted as a barren zone between the Crateceous and Paleocene by rejected the existence of Laffitteina bibensis and Bangiana hanseni at the lower Danian, SBZ2 was attributed to the upper Danian and the Selandian–early Thanetian were unified under SBZ3. However, some studies carring out in Anatolia and Iranian region might not entirely comply with it (see İnan, 1996; Sirel, 1998, 2015, 2018; İnan et al., 2005; Hottinger, 2014; Schlagintweit and Rashidi, 2019; Schlagintweit et al., 2020; Benedetti et al., 2021; Benedetti and Papazzoni, 2022). For instance, L. bibensis was reported for K/Pg boundary transition in shallow–very shallow water environment from Gölköy and Dündarlı (İnan, 1996; İnan et al., 2005; Sirel, 2015, 2018), moreover it is common in various Danian successions in other localities of Turkey unlike in Serra-Kiel et al. (2020). Since the regional variations can cause differences in the biostratigraphic ranges of the larger benthic foraminifera, reorganization of Paleocene Shallow Benthic Zones will be much more reliable when the studies not only from the European region but also eastern Tethys zone are taken into account.
Well-connected shelves between south Turkey, Iran and Oman (the Arabian platform) could have been pivotal in the dispersal of the genus in the Middle East region. According to the paleogeographic models (e.g., Scheibner and Speijer, 2008; Höntzsch et al., 2013; Martín-Martín et al., 2020) of Tethyan carbonate platforms, the regions discussed are located in low to intermediate latitudes (between 12°–30° N) in which the true coralgal reefs were replaced by larger benthic foraminifera, in accordance with the most prominent Paleocene eustatic sea-level fall (Scheibner and Speijer, 2008). Furthermore, the connections between vicinity platforms among the Middle East, Arabian and Africa areas are evidenced by some taxa dispersals from the Eocene deposits such as Somalina stefaninii (Silvesteri) and Dictyoconus aegyptiensis (Chapman), suggesting a limited geographic distribution in the above-mentioned basins during the Middle Eocene (Sallam et al., 2018). It can also be confirmed based on the biogeographic range of Alveolina kieli (Sirel) that is only recorded in Oman (Serra-Kiel et al., 2016), Pakistan (Rahman et al., 2021), Iran (Hadi et al., 2020) and Turkey (Sirel and Acar, 2008; Deveciler, 2014) during the middle Eocene. These clearly indicate occurrences of several larger foraminifera genera and species as endemic faunas, limited to the Turkish–Iranian–Pakistan and Oman basins during the Eocene. These findings are in accordance with the detailed paleogeographic reconstruction and the plate tectonic evidence from the Neo-Tethys Ocean in this timespan (see Golonka, 2004). Although the distribution details of the genus Burdurina will be an important, novel contribution in deciphering the correlations between the Middle East benthic ecosystems, whilst the information about this taxa is only limited to Benedetti et al. (2018) from Europe (western Tethys). Thus, the record of Burdurina genus across the Tethys is poor and does not permit a high-resolution paleogeography. Further work and taxonomic revisions on the miscellaneid taxa are needed to confirm the specific and generic attribution of the inclusive genera across the Tethys.
CONCLUSION
The present study undertakes for the very first time the study of upper Paleocene benthic foraminifera with precise details on the systematics of Burdurina selandinica from the Central Iran region (Golbui section). After the first description of Sirel (2018), some additional generic characteristics were defined and it was assigned to the biozone SBZ3 with the associated benthic foraminifera such as S. iranicus, Anatoliella sp., H. paleocenica, C. prima, M. cf. yvettae, E. dienii, I. sinjarica, R. conica, P. acuta and P. elongata. In this context E. dienii, H. paleocenica and R. conica are considered reaching up to SBZ3 in Iran whilst they are shown as a marker of SBZ2 for Mediterranean region. The genus Burdurina provides new insights in context of the paleogeographic implications about the spatial distribution of benthic foraminifera in the Middle East carbonate platforms during the Paleocene. Burdurina selandinica can be now considered to have a distribution ranging from the central (Middle East region) to western (only one location in Italy) Tethys areas, while previously it was considered restricted to the Turkey platform. The Golbui section is mainly characterized by benthic foraminifera and coralgal assemblages, which corresponds to the platform stage I of Scheibner and Speijer (2008). The biogenic components of this platform clearly indicate that the Middle East platforms had strongly distinct taxa composition compared to the western Tethys.
REFERENCES
Acar, S., Selandian benthic foraminiferal assemblages of the Southwestern Burdur (South of Lake Yarışlı, Western Turkey) and some taxonomic revisions, Bull. Miner. Res. Explor., 2019, vol. 158, pp. 49–119.
Afzal, J., Williams, M., Leng, M.J., Aldridge, R.J., and Stephenson, M.H., Evolution of Paleocene to Early Eocene larger benthic foraminifer assemblages of the Indus Basin, Pakistan, Lethaia, 2010, vol. 44, no. 3, pp. 299–320.
Alavi, M., Sedimentary and structural characteristics of the Paleo-Tethys remnants in northeastern Iran, Geol. Soc. Am. Bull., 1991, vol. 103, no. 8, pp. 983–992.
Bagherpour, B. and Vaziri, M.R., Facies, paleoenvironment, carbonate platform and facies changes across Paleocene Eocene of the Taleh Zang Formation in the Zagros Basin, SW-Iran, Hist. Biol., 2012, vol. 24, no. 2, pp. 121–142.
Benedetti, A., Mario, M., and Pichezzi, R.M., Paleocene to lower Eocene larger foraminiferal assemblages from central Italy: New remarks on biostratigraphy, Riv. Ital. Paleontol. Stratigr., 2018, vol. 124, no. 1, pp. 73–90.
Benedetti, A., Consorti, L., Schlagintweit, F., and Rashidi, K., Ornatorotalia pila n.sp. from the late Paleocene of Iran: Ecological, evolutional and paleobiogeographic inferences, Hist. Biol., 2021, vol. 33, no. 9, pp. 1796–1803.
Benedetti, A. and Papazzoni, C.A., Rise and fall of rotaliid foraminifera across the Paleocene and Eocene times, Micropaleontology, 2022, vol. 68, no. 2, pp. 185–196.
Berberian, M. and King, G.C.P., Towards a paleogeography and tectonic evolution of Iran, Can. J. Earth Sci., 1981, vol. 18, no. 2 pp. 210–265.
Di Carlo, M., Accordi, G., Carbone, F., and Pignatti, J., Biostratigraphic analysis of Paleogenelowstand wedge conglomerates of a tectonically active platform margin (Zakynthos Islands, Greece), J. Mediterr. Earth Sci., 2010, vol. 2, pp. 31–92.
Consorti, L. and Köroğlu, F., Maastrichtian-Paleocene larger Foraminifera biostratigraphy and facies of the Şahinkaya Member (NE Sakarya Zone, Turkey): Insights into the Eastern Pontides arc sedimentary cover, J. Asian Earth Sci., 2019, vol. 183, p. 103965.
Deveciler, A., Description of larger benthic foraminifera species from the Bartonian of Yakacık-Memlik region (N Ankara, central Turkey), Bull. Earth Sci. Appl. Res. Center Hacettepe Univ., 2014, vol. 35, no. 2, pp. 137–150.
Dunham, R.J., Classification of carbonate rocks according to depositional texture, in Classification of Carbonate Rocks-A Symposium. Tulsa, OK, USA, Am. Assoc. Petrol. Geol. Mem., 1992, pp. 108–121.
Embry, A.F. and Klovan, J.E., Absolute water depth limits of Late Devonian paleoecological zones, Geol. Rundsch., 1972, vol. 61, no. 2, pp. 672–686.
Flügel, E., Microfacies of Carbonate Rocks, Analysis, Interpretation and Application, Berlin: Springer-Verlag, 2004.
Ghose, B.K., Paleoecology of the Cenozoic reefal foraminifers and algae—A brief review, Palaeogeogr., Palaeoclimatol., Palaeoecol., 1977, vol. 22, no. 3, pp. 231–256.
Golonka, J., Plate tectonic evolution of the southern margin of Eurasian in the Mesozoic and Cenozoic, Tectonophysics, 2004, vol. 381, pp. 235–273.
Hadi, M., Less, G., and Vahidinia, M., Eocene larger benthic foraminifera (alveolinids, nummulitids, and orthophragmines) from the eastern Alborz region (NE Iran): Taxonomy and biostratigraphy implications, Rev. Micropaleontol., 2019, vol. 63, pp. 65–84.
Hadi, M., Consorti, L., and Vahidinia, M., Upper Ypresian to Lower Lutetian (SBZ 10 to 13) Alveolina stratigraphic horizons from the Hormak section, Zahedan District, Sistan Suture Zone, Eastern Iran, Micropaleontology, 2020, vol. 66, no. 4, pp. 317–337.
Holzmann, M. and Pawlowski, J., An updated classification of rotaliid foraminifera based on ribosomal DNA phylogeny, Mar. Micropaleontol., 2017, vol. 132, no. 2017, pp. 18–34.
Höntzsch, S., Scheibner, C., Brock,J., and Kuss, J., Circum-Tethyan carbonate platform evolution during the Palaeogene: The Prebetic platform as a test for climatically controlled facies shifts, Turk. J. Earth Sci., 2013, vol. 22, pp. 891–918.
Hottinger, L., Illustrated glossary of terms used in foraminiferal research, Carnet. Géol., Notebooks on Geology, 2006, p. CG2006 (M02).
Hottinger, L., The Paleocene and earliest Eocene foraminiferal family Miscellaneidae: Neither nummulitids nor rotaliids, Carnet. Geol., Notebooks on Geology, 2009, vol. 6, pp. 1–41.
Hottinger, L., Paleogene Larger Rotaliid Foraminifera from the Western and Central Neotethys, Springer Int. Publ., 2014.
İnan, N., The geographic expandition and stratigraphic distribution of Laffitteinaspecies in Turkey, Geol. Bull. Turk., 1996, vol. 39, no. 1, pp. 43–53.
İnan, N., Tasli, K., and Selim, İ., Laffitteinafrom the Maastrichtian–Paleocene shallow marine carbonate successions of the Eastern Pontides (NE Turkey): Biozonation and microfacies, J. Asian Earth Sci., 2005, vol. 25, pp. 367–378.
Leppig, U., Structural analysis and taxonomic revision of Miscellanea, Paleocene, Larger Foraminifera, Eclogae Geol. Helv., 1988, vol. 81, no. 3, pp. 689–721.
Leppig, U. and Langer, R.M., Emendation and taxonomic revision of Miscellanea juliettaepfenderae and M. juliettae villatea with designation of the respective holotype, Micropaleontology, 2015, vol. 61, no. 3, pp. 227–230.
Loeblich, A. and Tappan, H., Foraminiferal Genera and their Classification, NY: Von Nostrant Reinhold, 1987, vols. 1, 2.
Martín-Martín, M., Guerrera, F., Tosquella J., and Tramontana, M., Paleocene–Lower Eocene carbonate platforms of westernmost Tethys, Sediment. Geol., 2020, vol. 404, p. 105674.
Naderi Mighan, N. and Torshizian, H., Geological Map of Kadkan (1 : 100 000), GSI, 1999.
Okay, A., Alpine–Himalayan blueschists, Ann. Rev. Earth Planet. Sci., 1989, vol. 17, pp. 55–87.
Özcan, E., Abbasi, İ.A., Drobne, K., Govindan, A., Jovane L., and Boukhalfa, K., Early Eocene orthophragminids and alveolinids from the Jafnayn Formation, N Oman: Significance of Nemkovella stockari Less and Özcan, 2007 in Tethys, Geodin. Acta, 2016, vol. 28, pp. 160–184.
Rahaghi, A., Paleogene Biostratigraphy of Some Parts of Iran, Nat. Iran. Oil Comp., Geol. Laboratories, 1978.
Rahaghi, A., Stratigraphy and Faunal Assemblage of Paleocene–Lower Eocene in Iran, Nat. Iran. Oil Comp., Geol. Laboratories, 1983.
Rahman, M.U., Hanif, M., Jiang T., and Wahid, S., Alveolinids from the Lower Indus Basin, Pakistan (Eastern Neo-Tethys): Systematic and biostratigraphic implications, Geol. J., 2021, vol. 56, no. 1, pp. 1–28.
Sallam, E.S., Erdem, N.Ö., Sinanoğlu D., and Ruban, D.A., Mid-Eocene (Bartonian) larger benthic foraminifera from southeastern Turkey and northeastern Egypt: New evidence for the palaeobiogeography of the Tethyan carbonate platforms, J. Afr. Earth Sci., 2018, vol. 141, pp. 70–85.
Sarkar, S., Ecostratigraphic implications of a Late Palaeocene shallow-marine benthic community from the Jaintia Hills, Meghalaya, NE India, J. Earth Syst. Sci., 2020, vol. 129, no. 1, pp. 1–12.
Scheibner, C. and Speijer, R.P., Late Paleocene–early Eocene Tethyan carbonate platform evolution—A response to long-and short-term paleoclimatic change, Earth Sci. Rev., 2008, vol. 90, pp. 71–102.
Scheibner, C., Rasser M.W., and Mutti, M., The Campo section (Pyrenees, Spain) revisited: Implications for changing benthic carbonate assemblages across the Paleocene–Eocene boundary, Palaeogeogr., Palaeoclimatol., Palaeoecol., 2007, vol. 248, no.1–2, pp.145–168.
Schlagintweit, F. and Rashidi, K., Serrakielina chahtorshiana n. gen. et n. sp., and other (larger) benthic Foraminifera from Danian–Selandian carbonates of Mount Chah Torsh (Yazd Block, Central Iran), Micropaleontology, 2019, vol. 65, no. 4, pp. 305–338.
Schlagintweit, F., Rashidi, K., and Kohkan, H., Coscinospira prima n. sp., a new peneroplid foraminifer from Paleocene of Iran and its bearing on the phylogeny of the Peneroplidae, J. Mediterr. Earth Sci., 2020, vol. 12, pp. 3–14.
Schlagintweit, F., Rashidi R., and Mosavinia, A., First record of calcareous green algae (Dasycladales, Halimedaceae) from the Paleocene Chehel-Kaman Formation of North-Eastern Iran (Kopet-Dagh basin), Acta Palaeontol. Romaniae, 2021, vol. 17, no. 1, pp. 51–64.
Schmitz, B., Pujalte, V., Molina, E., Monechi, S., Orue-Etxebarria, X., Speijer, R.P., Alegret, L., Apellaniz, E., Arenillas, I., Aubray, M.P., Baceta, J.I., Berggren, W.A., Bernaola, G.F., Caballero, A., Clemmensen, J., Dinares-Turell, C., Dupuis, C., Heilman-Clausen, A., Hilario, R., Orus, Knox, M., Martin-Rubio, S., Ortiz, A., Payros, M.R., Petrizzo, K., Vonsalis, J., Sprong, E., Steurbaut, E., and Thomsen., E., The Global Stratotype Section and Points for the bases of the Selandian (Middle Paleocene) and Thanetian (Upper Paleocene) stages at Zumaia, Spain, Episodes, 2011, vol. 34, no. 4, pp. 220–243.
Septfontaine, M., Schlagintweit, F., and Rashidi, K., Pachicolumella nov. gen., shallow-water benthic imperforate foraminifera and its species from Maastrichtian and Paleocene of Iran, Micropaleontology, 2019, vol. 65, no. 2, pp. 145–160.
Serra-Kiel, J., Hottinger, L., Caus, E., Drobne, K., Ferrandez, C., Jauhri, A.K., Less, G., Pavlovec, R., Pignatti, J., Samso, J.M., Schaub, H., Sirel, E., Strougo, A., Tambareau, Y., Tosquella J., and Zakrevskaya, E., Larger foraminiferal biostratigraphy of the Tethyan Paleocene and Eocene, Soc. Géol. Fr., 1998, vol. 169, pp. 281–299.
Serra-Kiel, J., Vicedo, V., Razin P.H., and Grélaud, C., Selandian–Thanetian large foraminifera from lower Jafnayn Formation in the Sayq area (eastern Oman Mountains), Geol. Acta, 2016, vol. 14, no. 3, pp. 315–333.
Serra-Kiel, J., Vicedo, V., Baceta, J.J., Bernaota, G., Robador, A., Paleocene larger foraminifera from the Pyrenean Basin with a recalibration of the Paleocene shallow bentic zones, Geol. Acta, 2020, vol. 18, no. 8, pp. 1–69.
Shafaii-Moghadam, H. and Stern, R.J., Ophiolites of Iran: Keys to understanding the tectonic evolution of SW Asia: (I) Paleozoic ophiolites, J. Asian Earth Sci., 2014, vol. 91, pp. 19–38.
Sirel, E., The species of Miscellanea Pfender, 1934 (Foraminiferida) in the Thanetian–Ilerdian sediments of Turkey, Rev. Paléobiol., 1997, vol. 16, no. 1, pp. 77–99.
Shafaii-Moghadam, H., Khedr, M.Z., Arai, S., Stern, R.J., Ghorbani, G., Tamura A., and Ottley, C.J., Arc-related harzburgite–dunite–chromitite complexes in the mantle section of the Sabzevar ophiolite, Iran: A model for formation of podiform chromitites, Gondwana Res., 2015, vol. 27, no. 2, pp. 575–593.
Sirel, E., Foraminiferal description and biostratigraphy of the Paleocene-lower Eocene shallow-water limestones and discussions on the Cretaceous–Tertiary boundary in Turkey, Gen. Dir. Miner. Res. Explor., Monogr. Ser. 2, 1998.
Sirel, E., Reference Sections and Key Localities of the Paleogene Stages and Discussion on C–T, P–E and E–O boundaries by the Very Shallow–Shallow Water Foraminifera in Turkey, Ankara Univ. Publ., 2015.
Sirel, E., Revision of the Paleocene and partly Early Eocene larger benthic foraminifera of Turkey, Ankara Univ. Publ., 2018, vol. 27.
Sirel, E. and Acar, S., Description and Biostratigraphy of the Thanetian–Bartonian Glomalveolinids and Alveolinids of Turkey, TMMOB Jeoloji Mühendisleri Odası, 2008.
Sirel, E. and Deveciler, A., Diagnostic structural elements of Ranikothalia Caurdi and re-description of its six species from Thanetian–Iledian of Turkey, J. Palaeontol. Soc. India, 2018, vol. 63, no. 1, pp. 19–35.
Smith, A.G., Smith D.G., and Funnel, B.M., Atlas of Mesozoic and Cenozoic Coastlines, Cambridge: Cambrige Univ. Press, 1994.
Stöcklin, J., Structural history and tectonics of Iran: A review, AAPG Bull., 1968, vol. 52, pp. 1229–1258.
Tirrul, R., Bell, I., Griffis R., and Camp, V., The Sistan suture zone of eastern Iran, Geol. Soc. Am. Bull., 1983, vol. 94, pp. 134–150.
Vršič, A., Machaniec, E., and Gawlick, H. J., Middle to Late Paleocene larger benthic foraminifera from the Sirt Basin (Libya), Rev. Micropaléontol., 2021, vol. 71, pp. 1–15.
Zamagni, J., Mutti, M., and Košir, A., Evolution of shallow benthic communities during the Late Paleocene-earliest Eocene transition in the Northern Tethys (SW Slovenia), Facies, 2008, vol. 54, no. 1, pp. 25–43.
Zhang, Q., Willems H., and Ding, L., Evolution of the Paleocene-Early Eocene larger benthic foraminifera in the Tethyan Himalaya of Tibet, China, Int. J. Earth Sci., 2013, vol. 102, no. 5, pp. 1427–1445.
ACKNOWLEDGMENTS
We greatly appreciate the discussions and suggestions of Dr. Ercüment Sirel on the larger foraminifera. Many thanks to Prof. Elena Zakrevskaya (Vernadsky State Geological Museum), Prof. Shreerup Goswami (Sambalpur University) and Prof. Yu.D. Zakharov (Far East Geological Institute, Far East Branch of Russian Academy of Sciences) for their careful review which significantly improved the manuscript. Also, we would especially like to thank the editor, Dr. Marina Bylinskaya (Stratigraphy and Geological Correlation Journal), for editorial support.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The authors declare that they have no conflicts of interest.
Additional information
Reviewers: Shreerup Goswami, E.Yu. Zakrevskaya, and Yu.D. Zakharov
Rights and permissions
About this article

Cite this article
Deveciler, A., Hadi, M., Sarkar, S. et al. First Report of Burdurina selandinica Sirel (Foraminiferida) and the Microfacies Analysis of Middle Paleocene Limestones in Central Iran. Stratigr. Geol. Correl. 30, 518–534 (2022). https://doi.org/10.1134/S0869593822060041
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0869593822060041