
Abstract
Lower Miocene strata are exposed in the northwest of Ashtian, in the Central Iran Zone. Biostratigraphy of the Qom Formation was investigated using dinoflagellate cyst and benthic foraminifera. The stratigraphic distribution of the dinocysts lead to the identification of two distinctive stratigraphically non-continuous palynozones viz., Caligodinium amiculum (late Aquitanian) and Cordosphaeridium cantharellus (early Burdigalian) zones. This biozonation is equivalent to that recognized in Northwest Europe. Moreover, the presence of such stratigraphically distinctive taxa as Borelis melo curdica, Meandropsina iranica, Elphidium sp. 14, and Miogypsina sp. enabled the identification of the two consecutive biozones: the Elphidium sp. 14—Miogypsina Assemblage Subzone and the Borelis melo group Meandropsina iranica Assemblage Zone. The late Aquitanian–Burdigalian age is collectively attributed, therefore, to the studied section. This age assignment is based on the dinocysts and foraminifer species compared with other stratigraphic sections. The close resemblance of the dinoflagellate cyst assemblages of the Qom Basin with Northwest Europe and the northwestern Tethys during the Oligocene-early Miocene indicates a marine connection and faunal exchange between the two regions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 INTRODUCTION
The Qom Formation was deposited on the northeastern coast of the Tethyan Seaway (Reuter et al., 2009a) during the final sea transgression of the Oligocene–early Miocene. In its position as an Iranian gateway, the Qom Sea played a predominant role in connecting the Mediterranean Tethys to the northwest and the Indo-Pacific to the southeast. This sea served as a significant link between the European-Mediterranean and Indo-Pacific areas and as a channel for exchanging the faunal assemblages of these two paleogeographic bioprovinces (Fig. 1). The exact timing of the closure of this seaway, known as the Terminal Tethyan Event, is still a matter of debate (Reuter et al., 2009a). Given the collision of the African/Arabian and Iranian/Eurasian plates, F. Bozorgnia (1966) proposed the late early Miocene (Burdigalian) age as the time of the Event. It is evident that by the beginning of the middle Miocene age, each region had developed its own individual biota (Reuter et al., 2009a; Mohammadi et al., 2013).

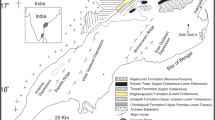
Paleogeographic map for the early Burdigalian (modified from Popov et al., 2004; Cornacchia et al., 2018).
The Qom Formation has several notable features, such as fossil richness, tectonic complexity, unique facies, and petroleum potential, making it an interesting topic for several studies. Y. Zhu et al. (2007) reported six species of dinoflagellate cysts (distributed among six genera) for the first time from two sections in the Qom–Kashan area in Central Iran. They suggested the Eocene age as the time of formation based on dinoflagellate cysts, benthic foraminifers, ostracods, and calcareous nannoplankton assemblages. However, M. Reuter et al. (2009b) criticized their findings, citing poor taxonomy and relevant previous and subsequent studies that broadly dated the formation to the Oligocene-Miocene age. Moreover, J. Daneshian et al. (2008) and J. Daneshian and A. Aftabi (2010) investigated the sections Y. Zhu et al. (2007) had studied. They continued to present foraminiferal assemblages established by previously recognized ages, as they could not find the foraminifera described by Y. Zhu et al. (2007).
In this study, we discuss the previous zonation and age attributed to the Qom Formation in greater detail. The aim of this research is to establish a biostratigraphy framework for the formation using both dinoflagellate cysts and benthic Foraminifera.
2 GEOLOGICAL SETTING
E. Heydari et al. (2003) pointed out eight geological zones of the Iranian Plateau: (1) Central Iran, (2) Sanandaj–Sirjan, (3) Urumieh–Dokhtar magmatic arc, (4) Zagros, (5) Alborz, (6) Koppeh Dagh (a.k.a. Kopet Dagh), (7) Lut, and (8) Makran.
The Qom Formation is widely distributed in three NW-SE trending basins: The Central Iran back-arc basin, the Sanandaj-Sirjan fore-arc basin, and the Urumieh-Dokhtar intra-arc basin (e.g., Stöcklin and Setudehnia, 1991; Daneshian and Ramezani Dana, 2007; Reuter et al., 2009a; Mohammadi et al., 2011, 2013).
According to this subdivision, the study area is located about 3 km northwest of Ashtian in the Central Iranian basin. The geographical coordinates of the studied section are 34°32′15′′ N and 49°58′35′′ E (Fig. 2). The succession, which has a thickness of 72 m, mainly consists of shale, marl, and alternations of limestone and marl at the section studied. This discontinuously follows the Oligocene Lower Red Formation and is disconformably overlain by the Miocene Upper Red Formation.

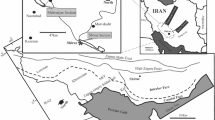
Location map of the studied stratigraphic section and simplified geological map of the study area (modified from the 1 : 1 00 000 geological map of Farmihan by Hajian (1970)).
3 MATERIAL AND METHODS
In the section studied, 35 samples were collected for palynological and foraminiferal biostratigraphy analysis. Following standard palynological processing procedures (e.g., Traverse, 2007), samples were prepared comprising treatment with hydrochloric (10–50%) and hydrofluoric (ca. 40%) acids for the dissolution of carbonates and silicates, respectively. Thereafter, hot HCl (50%) was applied to dissolve the newly formed silica gel during the HF treatment. The solution was neutralized and centrifuged in ZnCl2 (with a specific gravity of 1.9 g/cm3) for density separation. Neither oxidation nor alkali treatments were applied. Residues were sieved through a 20 μm mesh prior to making strew slides. Moreover, thin sections were prepared to study foraminiferal content. All rock samples, mounted slides, and thin sections were stored in the collections of the Faculty of Science, Imam Khomeini International University, Qazvin, Iran.
4 BIOSTRATIGRAPHY
There are a limited number of studies in the literature on dinocysts from the Neogene Period at low latitudes, especially in SW Asia (Mao and Lei, 1996; Mao et al., 2004; Jafar-Nezhad et al., 2015). Almost all integrated stratigraphical studies on the Oligo-Miocene Qom Formation emphasized planktonic and larger benthic foraminifers (e.g., Daneshian and Ramezani Dana, 2007).
Based on the characteristics of the assemblages and the lowest/highest occurrences of index taxa of dinocysts, two of the Northwestern European biozones proposed by E. Dybkjær and E.S. Piasecki (2010), which are Caligodinium amiculum Interval Zone and Cordosphaeridium cantharellus Interval Zone, can be clearly recognized within the Qom Formation. Between these two palynozones, there are two other palynozones named the Thalassiphora pelagica Zone and the Sumatradinium hamulatum Zone, which were not identified due to the barren limestone deposits. This gap was covered by additional foraminifer studies. T.D. Adams and F. Bourgeois (1967) established formal biozones for the Asmari Formation in the Zagros Zone. This biozonation is practical for the Qom Formation in Central Iran because of the faunal similarity between these two regions (e.g., Daneshian and Ramezani Dana, 2019).
Due to the presence of key index taxa of foraminifera, two biozones related to T.D. Adams and F. Bourgeois (1967), Elphidium sp. 14—Miogypsina Assemblage Subzone (late Aquitanian) and Borelis melo group-Meandropsina iranica Assemblage Zone (Burdigalian), were identified in the calcareous layers of the studied section. Our investigations show that the calcareous layer with barren palynomorphs coincides with the upper part of Elphidium sp. 14—Miogypsina Assemblage Subzone and the lower part of Borelis melo group-Meandropsina iranica Assemblage Zone.
Stratigraphic ranges of the dinocyst and foraminiferal species occurring in the Qom Formation, as investigated herein, are illustrated in Fig. 3.

Stratigraphic distribution of dinoflagellate cyst and foraminifera species throughout the Qom Formation as investigated herein.
4.1 Palynostratigraphy
Palynological assemblages from the surface samples of the Qom Formation contain reasonably diverse and moderately preserved palynofloras dominated by marine-derived palynomorphs (especially dinoflagellate cysts, 20 species distributed among 17 genera); acritarchs; and terrestrial elements, including spores, pollen grains, tetrads, and fungal spores occurring as minor constituents. Representative dinocyst specimens are illustrated in Fig. 4.

Selective dinocyst species of the Qom Formation in the northwest of Ashtian, Central Iran Zone: (a, b) Cordosphaeridium cantharellus (Brosius) Gocht, 1969; (c) Melitasphaeridium choanophorum (Deflandre and Cookson) Harland and Hill, 1979; (d) Spiniferites ramosus (Ehrenberg) Mantell, 1854; (e) Dapsilidinium sp.; (f, g) Homotryblium plectilum Drugg and Loeblich, 1967; (h) Homotryblium tenuispinosum Davey and Williams, 1966; (i) Cleistosphaeridium placacanthum (Deflandre and Cookson) Eaton et al., 2001; (j) Operculodinium sp.; (k) Exochosphaeridium insigne de Verteuil and Norris, 1996; (l) Hystrichokolpoma sp.; (m) Thalassiphora pelagica (Eisenack) Eisenack and Gocht (1960) emend. Benedek and Gocht, 1981; (n) Thalassiphora rota Schiøler, 2005; (o) Tuberculodinium vancampoae (Rossignol) Wall, 1967; (p) Caligodinium amiculum Drugg, 1970; (q) Hystrichosphaeropsis obscura Habib, 1972; (r–t) Deflandrea phosphoritica Eisenack, 1938; (u, v) Apteodinium sp.; (w) Cribroperidinium tenuitabulatum (Gerlach) Helenes, 1984; (x) Membranophoridium aspinatum Gerlach, 1961. The scale bars represent 50 μm.
4.1.1. Caligodinium amiculum interval zone. This zone extends from Sample 1 to Sample 16 and includes 31 m of the studied section. It is characterized by the last occurrence of Homotryblium spp. to the last occurrence of Caligodinium amiculum and is assigned the age of late Aquitanian (Dybkjær and Piasecki, 2010). It is characterized by the co-occurrence of the following: Cleistosphaeridium placacanthum, Cribroperidinium tenuitabulatum, Hystrichokolpoma rigaudiae, Lingulodinium machaerophorum, Melitasphaeridium choanophorum, Membranosphaeridium aspinatum, Polysphaeridium zoharyi, Cordosphaeridium cantharellus, Spiniferites ramosus, Thalassiphora, Thalassiphora pelagica, and Thalassiphora rota. The last occurrences of Caligodinium amiculum strongly indicate the Aquitanian age. Indeed, the last occurrences of Deflandrea phosphoritica in the lower section of the studied interval (0–6.5 m) also indicate the Aquitanian age (de Verteuil and Norris, 1996; Dybkjær and Rasmussen, 2007).
4.1.2. Cordosphaeridium cantharellus interval zone. This zone extends from Sample 24 to Sample 32 and includes 14.5 m of the studied section. It is defined from the first occurrence of Exocosphaeridium insigne to the last occurrence of Cordosphaeridium cantharellus and is regarded as early Burdigalian in terms of age (Dybkjær and Piasecki, 2010). It is characterized by the co-occurrence of the following: Apteodinium australiense, Apteodinium spiridoides, Hystrichosphaeropsis obscura, and Tuberculodinium vancampoae.
Additionally, dinocyst species such as Cleistosphaeridium placacanthum, Cordosphaeridium cantharellus, Cribroperidinium tenuitabulatum, Lingulodi-nium machaerophorum, Melitasphaeridium choanophorum, Spiniferites ramosus, and Polysphaeridium zoharyi extend from the first palynozone to this zone.
4.2 Foraminiferal Stratigraphy
Three formal biozones, as well as two subzones, were established by T.D. Adams and F. Bourgeois (1967) for the Asmari Formation, Zagros basin. These include, in ascending order, the Eulepidina-Nephrolepidina-Nummulites Assemblage Zone (Oligocene), the Archaias asmaricus-Archaias hensoni Assemblage Subzone, and the Elphidium sp. 14—Miogypsina Assemblage Subzone, which were differentiated from the Miogypsinoides–Archaias–Valvulinid Assemblage Zone (Aquitanian) and the Borelis melo group-Meandropsina iranica Assemblage Zone (Burdigalian).
With reference to the biozonation introduced by T.D. Adams and F. Bourgeois (1967) and the presence of such stratigraphically distinctive taxa as Borelis melo curdica, Meandropsina iranica, Elphidium sp. 14, and Miogypsina sp., the Elphidium sp. 14—Miogypsina Assemblage Subzone and the Borelis melo group-Meandropsina iranica Assemblage Zone are identified. Representative foraminifer specimens are illustrated in Fig. 5.

Illustrations of the representative foraminifer specimens of the Qom Formation in the northwest of Ashtian, Central Iran: (a) Austrotrillina howchini Schlumberger, 1893; (b) Borelis melo (Fichhtel and Moll) curdica Reichel, 1937; (c) Elphidium sp. 1 Adams and Bourgeois, 1967; (d) Miogypsina spp.; (e) Elphidium sp.14 Thomas, 1949; (f) Dendritina rangi d’Orbigny emend. Fornasini, 1904; (g) Tubucellaria spp.; (h) Ditrupa sp.; (i) Amphistegina sp.; (j) Operculina complanata Defrance, 1822; (k) Meandropsina anahensis Henson, 1950; (l) Lithophyllum sp. The scale bars represent 100 μm.
4.2.1. Elphidium sp. 14—Miogypsina Assemblage Subzone. This subzone extends from Sample 1 to Sample 23 and comprises 47.5 m of the studied interval. It is characterized by the concurrence of Elphidium sp. 14 and Miogypsina sp. The lower boundary of this biosubzone is defined by the first occurrence of Elphidium sp. 14, and the upper boundary is recognized by the first occurrence of Borelis melo curdica. Miogypsina has its stratigraphic importance within the early Miocene age (especially Aquitanian) of Iran (Adams et al., 1983). Associated foraminifera are Asterigerina rotula, Austrotrilina howchini, Dendritina rangi, Elphidium sp. 14, Elphidium sp. 1, Meandropsina anahensis, Meandropsina iranica, Miogypsina sp., Miogypsinoides sp., Operculina complanata, Pyrgo spp., Rotalia viennoti, and Triloculina trigonala.
4.2.2. Borelis melo group-Meandropsina iranica Assemblage Zone. The upper part of the studied section is similar to the Borelis melo group-Meandropsina iranica Assemblage Zone of T.D. Adams and F. Bourgeois (1967), which indicates a Burdigalian age for this part. This zone extends from Sample 24 to Sample 35 and includes 35.5 m of the Qom Formation at the studied section.
The lower boundary of this Assemblage Zone is defined by the first occurrence of the Burdigalian reliable marker species Borelis melo curdica (Wynd, 1965; Adams and Bourgeois, 1967; Laursen et al., 2009; Van Buchem et al., 2010). According to the first occurrence of the marker species, the Aquitanian–Burdigalian boundary is in sample no. 19 in the studied interval. Associated foraminifera are Amphistegina spp., Asterigerina rotula, Dendritina rangi, Elphidium sp. 14, Meandropsina iranica, Miogypsina sp., Miogypsinoides sp., Pyrgo spp., and Triloculina trigonala.
Therefore, based on the dinocyst and foraminifera content, the Qom Formation is collectively dated as early Miocene (late Aquitanian–Burdigalian).
5 DISCUSSION
Historical investigations of the Qom Formation were conducted by J.J. Dozy (1944), who gave these deposits their name. M.A. Furrer and P.A. Soder (1955) defined six lithostratigraphic members (a, b, c, d, e, and f) of the Qom Formation by examining several sections in the type area near the city of Qom. They also proposed an Oligocene–Miocene age. According to lithological and palaeontological characteristics, F. Bozorgnia (1966) extended these units to ten members. Today, the stratigraphic lexicon of Iran includes nine defined lithological members (a, b, c-1, c-2, c-3, c-4, d, e, and f) for the type locality of the Qom Formation (Stӧcklin and Setudehnia, 1991).
Several studies have been conducted based on different fossil groups to investigate the age of the Qom Formation (Fig. 6). Among the different fossil groups, benthic and planktonic foraminifera play a key role in biozonation and age dating of the Qom Formation (Table 1).

Age range of the Qom Formation using different fossil groups including benthic and planktonic foraminifera, brachiopods, bryozoans, nanofossils, echinoderms, and gastropods.
Large benthic foraminifera in different areas of the basin were used to estimate the age of the Qom Formation. The age of the Aquitanian–early Burdigalian for the interval examined in Central Iran was suggested based on the occurrence of marker taxa of foraminifera and on the definition of two biozones of T.D. Adams and F. Bourgeois (1967): the Borelis melo group-Meandropsina iranica assemblage zone and the Miogypsinoides–Archaias–Valvulinid assemblage zone (Daneshian and Ramezani Dana, 2007). In the Zanjan province, the age of the Aquitanian-late Burdigalian for the interval examined was suggested based on the recognition of the two biozones of Adams and Bourgeois mentioned above (Daneshian et al., 2010). E. Behforouzi and A. Safari (2011) studied the large benthic foraminifera of the Qom Formation in the Chenar area of northwestern Kashan, Iran. They suggested the age of Oligocene for the interval examined based on the Lepidocyclina–Operculina–Ditrupa assemblage zone of J. Wynd (1965) and G.V. Laursen et al. (2009) applied to the Asmari Formation. M. Yazdi-Moghadam (2011) defined the European Standard Shallow Benthic Zonation SBZ 21 and assigned an early-to-middle Rupelian age for the interval examined to the south of Uromieh, northwestern Iran. M. Yazdi et al. (2012) investigated the Qom Formation in northeastern Isfahan, central Iran. They recognized the Globigerina spp. zone (Zone 55) of J. Wynd (1965) and the Lepidocyclina–Operculina–Ditrupa assemblage zone of J. Wynd (1965) and G.V. Laursen et al. (2009) in the lower and middle parts of the section studied, respectively. The age assigned to these parts is Oligocene (Rupelian–Chattian). Based on the stratigraphic layer position in the uppermost parts of the Qom Formation, Chattian and probably early Miocene (Aquitanian?) ages were assigned to these parts (Yazdi et al., 2012). In the western margin of the Urumieh–Dokhtar magmatic arc, M. Karevan et al. (2014) assigned the interval examined to the Oligocene age based on benthic foraminifera and coral assemblages. They defined Lepidocyclina–Operculina–Ditrupa Assemblage zone that J. Wynd (1965) and G.V. Laursen et al. (2009) had established for the Asmari Formation in Oligocene. I. Maghfouri-Moghadam et al. (2014) reported a Chattian-Aquitanian benthic foraminifera existed at the Qom Formation south and west of Tafresh, Central Iran. M. Nouradini et al. (2014) investigated the Qom Formation in the Bagher Abad area, located in northeast Isfahan. Based on the Lepidocyclina–Operculina–Ditrupa assemblage zone of J. Wynd (1965) and G.V. Laursen et al. (2009), and Nummulites intermedius–Nummulites vascus assemblage zone of J. Wynd (1965), they attributed Oligocene and early Miocene (Aquitanian) ages to the lower and middle parts of the studied interval. Because of the absence of foraminifera in the upper part of the studied section, macrofossils (Bivalves and Echinoids) have been used to determine this part came from the Burdigalian age. In the Sanandaj–Sirjan zone (fore-arc basin in northern Abadeh area), the foraminiferal assemblages were used to attribute the Qom formation to Rupelian–Chattian age (Mohammadi and Ameri, 2015). E. Mohammadi et al. (2015) investigated three stratigraphic sections, Bujan, Varkan, and Khurabad, of the Qom Formation in Sanandaj-Sirjan fore-arc and Central Iran back-arc basins. According to biostratigraphy data and foraminifera assemblages, they attributed Qom Formation to the Rupelian–Chattian, Rupelian, and Rupelian–Burdigalian ages.
In the Sanandaj-Sirjan Metamorphic Belt in the Kerman, M.J. Hassani and F. Hosseinipour (2018) dated the Qom Formation as belonging to the early Miocene age based on benthic foraminifera, such as Borelis pygmaea, Borelis melo curdica, Austrotrillina howchini, Rotalia viennoti, Miogypsina spp., Miogypsinoides spp. and Elphidium spp. Nonspiral hyaline foraminifera of the Qom Formation found in the area north of the Central Iran basin was investigated by J. Daneshian and L. Ramezani Dana (2016b), who suggested it came from the Aquitanian–Burdigalian age.
Larger benthic foraminiferal index taxa have also been used to assign a Burdigalian age for the Qom formation in the Khoy-Bostan Abad area, which is correlated with Zone SBZ 25, the European standard shallow benthic zonation (Yazdi-Moghadam et al., 2018a). This study was not the only one that correlated the Qom Formation foraminifer zones with European standard shallow benthic zonation of B. Cahuzac and A. Poignant (1997). M. Yazdi-Moghadam et al. (2018b) considered hyaline and porcellaneous larger foraminifera of the formation and recognized SBZ 21 Zone of the European standard shallow benthic zonation. An age of early Rupelian was assigned to this studied interval located north of Sonqor in northwest Iran. M. Yazdi-Moghadam et al. (2021) illustrated three outcrop sections of the Qom Formation that are situated in northwest Iran (Hamedan–Miandoab area). They recognized SBZ 25 Zone and assigned a Burdigalian age to the intervals studied. J. Daneshian and L. Ramezani Dana (2019) established three characteristic stratigraphically consecutive biozones, viz., Peneroplis farsensis interval zone (early to early late Aquitanian), Elphidium sp. 14 interval zone (late late Aquitanian) and Borelis melo curdica total range zone (Burdigalian) for the Qom Formation in the north Central Iran Zone. Additionally, they suggested that the first occurrence of Borelis melo curdica is at the Aquitanian/Burdigalian boundary.
H. Noroozpour (2020) by investigating Qom Formation in southwestern Zanjan province, Central Iran, recognized Borelis melo curdica-Borelis melo melo Assemblage Zone, with the age of Burdigalian (early Miocene). The author was assigned 5-meters in the base of the Qom deposits to the Aquitainian (?) due to absence of Borelis melo curdica. Based on the larger benthic foraminifera, an age of early Chattian allocated to Qom Formation at the Tajar-Kuh stratigraphic section, Central Iran (Akbar-Baskalayeh et al., 2020).
Planktonic foraminifera were also the subject of several studies for the Qom Formation. M. Nouradini et al. (2015) investigated Qom Formation in the Bagh section, northeast Isfahan of Central Iran. They recognized two main planktonic foraminiferal zones in their studied succession. These are, in ascending stratigraphical order, the Globigerinoides primordius Zone in the lowermost and the Globigerinoides trilobus Zone in the upper most part of the Qom Formation, which were signifying Aquitanian and Burdigalian age, respectively. J. Daneshian and L. Ramezani Dana (2016a) recorded new Aquitanian–Burdigalian genera and species of agglutinated foraminifera of the Qom Formation, in the Dobaradar region, in the southwest of Qom city. They also defined M1 to M4a, planktovertnic foraminifera biozones of B.S. Wade et al. (2011) with the age of Aquitanian–Burdigalian in their interval studied.
In some studies, the Qom formation was investigated using both benthic and planktonic foraminifera species. The presence of key benthic and planktonic foraminifera species within a studied succession in southeastern Ashtian, Central Iran Zone has been used to attribute a late Aquitanian and Burdigalian age to the lower and upper parts of the section investigated, respectively (Daneshian and Deziani, 2004). J. Daneshian and L. Ramezani Dana (2017) recognized the Praeorbulina sicana lowest-occurrence Subzone (M5a), biozone of B.S. Wade et al. (2011) with a late late Burdigalian age in the upper part and Miogypsinoides–Archaias–Valvulinid Assemblage Zone of Adams and Bourgeois (1967) with an Aquitanian age in the lower part of the interval examined in the north-central Iran zone. Therefore, Qom Formation is assigned to Aquitanian-late Late Burdigalian in their studied section.
M.J. Hassani and M.R. Vaziri (2011) undertook a study of the Marine gastropod assemblage of the Qom Formation from Khavich Area, South of Sirjan. These authors focused their study on the lower Miocene, largely based on gastropod evidence. They mentioned early Miocene age is also confirmed by benthic foraminifera and ostracods, found with gastropods in the interval examined. The large size of Strombus roegli and its abundance are interpreted as a Chattian strombid in the basin, which indicates a distinct warming trend (Harzhauser, 2001).
A. Pedramara et al. (2019) reported bryozoans and brachiopods from the lower Miocene deposits of the Qom Formation in northeast Isfahan (Central Iran). Echinoids and planktonic foraminifera of the Qom Formation were investigated by K. Khaksar and I. Maghfouri-Moghadam (2007), which suggested an Oligo-Miocene age. Aquitanian pelecypods and ostracods were also reported from this formation in the west of Ashtian (Maghfouri-Moghadam and Yasboulaghi, 2015; Maghfouri-Moghadam, 2015).
The nanostratigraphy of the Qom Formation in Kamar-Kuh area, North-Central Iran was studied by F. Hadavi et al. (2010), and they suggested that the studied section belonged to the Burdigalian–Serravalian age. Late Oligocene to early Miocene calcareous nannofossil assemblage was also reported from the formation in the Central Iran Basin by M. Parandavar and F. Hadavi (2019).
The palynozones defined in this research study are compared with previously published dinoflagellate cyst zonations from Northwestern Europe (Denmark, Germany, and the Netherlands), the East Coast of the US, Egypt, and the South China Sea to attain a global biostratigraphic framework based on dinoflagellate cysts for the early Miocene age (late Aquitanian-early Burdigalian). Benthic foraminifera and dinoflagellate zonation of the Qom Formation in the present study are correlated with those of a covalent age in Iran and different parts of the world (Table 2).
In this research, the Caligodinium amiculum Interval Zone corresponds to the lower part of Zone DN2 of L. de Verteuil and G. Norris (1996) of Eastern U.S.A., the lower part of DN2 of A. Köthe (2003) of the North Sea, and part of Zone D16 of A.J. Powell and H. Brinkhuis (2004). The Cordosphaeridium cantharellus Interval Zone correlates with the upper part of Zone DN2 of L. de Verteuil and G. Norris (1996) of Eastern U.S.A., Zone DN2 of A. Köthe (2003), and the upper part of Zone D16c of A.J. Powell and H. Brinkhuis (2004). According to this comparison and the lowest/highest occurrences of index taxa of dinocysts, these palynozones are dated as early Miocene (late Aquitanian–early Burdigalian).
Furthermore, of the five biozones introduced by A. Soliman et al. (2012) for the lower and middle Miocene strata of Egypt, only the Lingulodinium machaerophorum Assemblage Biozone (GOS1), which is assigned to Aquitanian through mid-Burdigalian, is more or less equivalent to the Caligodinium amiculum Interval Zone proposed herein. The shared species of these two regions include Cribroperidinium spp., Apteodinium spiridoides, Cleistosphaeridium placacanthum, Cordosphaeridium cantharellus, and Melitasphaeridium choanophorum.
Besides, both the identified Zones are comparable with the Polysphaeridium zoharyi Assemblage Zone of S. Mao et al. (2004) of early Miocene in the South China Sea. A special characteristic of this zone is the first occurrence of typical Miocene species such as Hystrichosphaeropsis obscura and Melitasphaeridium choanophorum (Mao et al., 2004).
Hystrichosphaeropsis obscura from the Miocene is well known in various parts of the world. The first occurrence datum of this species is associated with Tuberculodinium vancampoae, a Neogene indicator of Zone D16 of the Northwest European zonation of A.J. Powell and H. Brinkhuis (2004), which is the latest Oligocene-earliest Miocene (Heilmann-Clausen and Costa, 1990). Moreover, these recognized zones are more or less equivalent to the SNSM2 zone of D.K. Munsterman and H. Brinkhuis (2004) of late Aquitanian (earliest)-early Burdigalian erected in the southern North Sea basin (the Netherlands). In the two regions (latest Oligocene-earliest Miocene), shared species include Hystrichosphaeropsis obscura, Apteodinium spp., Homotryblium plectilum, Lingulodinium machaerophorum, Polysphaeridium zoharyi and Spiniferites spp.
The ranges of Cordosphaeridium cantharellum, Homotryblium plectilum, Hystrichokolpoma rigaudiae, Lingulodinium machaerophorum, and Polysphaeridium zoharyi may extend upward into younger layers. However, upon presenting together with such species as Hystrichosphaeropsis obscura, Melitasphaeridium choanophorum, and Tuberculodinium vancampoae, their co-occurrence is indicative of a Miocene age (Mao et al., 2004).
In terms of common key species, these palynozones are comparable with the ±coeval dinoflagellate cyst biozones of A. Soliman et al. (2012) for the Gulf of Suez, Egypt, S. Mao et al. (2004) for the South China Sea, D.K. Munsterman and H. Brinkhuis (2004) and A. Köthe (2003) for the North Sea basin (Netherlands), A.J. Powell and H. Brinkhuis (2004) for Northwest Europe, as well as L. de Verteuil and G. Norris (1996) for North Atlantic. Described here, these comparisons are illustrated in Table 2. The paleogeographical position of the Qom Formation at the northeastern coast of the Tethyan Seaway result in the similarity of dinocyst species between Qom Formation and Northwestern European coeval strata during the Oligocene-early Miocene age (Reuter et al., 2009a).
6 CONCLUSIONS
Detailed investigation of the dinocysts and foraminifera taxa provides stratigraphic frameworks for the lower Miocene succession of the Qom Formation, northwest of Ashtian, Central Iran. All examined samples yielded moderately preserved palynomorph assemblages. Two non-continuous palynozones can be recognized: the Caligodinium amiculum (late Aquitanian) and Cordosphaeridium cantharellus (early Burdigalian) zones. Moreover, two consecutive foraminifer biozones (Elphidium sp. 14—Miogypsina Assemblage Subzone and Borelis melo group–Meandropsina iranica Assemblage Zone) were identified in the studied interval. An age of early Miocene (late Aquitanian–Burdigalian) is assigned for the studied section, based on the co-occurring ranges of well-known dinocysts and foraminifera taxa to the standard previously published zonations.
REFERENCES
Adams, T.D. and Bourgeois, F., Asmari biostratigraphy, Iranian Oil Operating Company, Geological and Exploration Division, 1967, Report no. 1074, pp. 1–37.
Adams, C.G., Gentry, A.W., and Whybrow, P.J., Dating the terminal Tethyan event, Utrecht Micropaleontol. Bull., 1983, vol. 30, pp. 273–298.
Akbar-Baskalayeh, N., Less, G., Ghasemi-Nejad, E., Yazdi-Moghadam, M., and Pignatti, J., Biometric study of late Oligocene larger benthic Foraminifera (Lepidocyclinidae and Nummulitidae) from the Qom Formation, Central Iran (Tajar-Kuh section), J. Paleontol., 2020, vol. 94, pp. 593–615.
Bassi, D., Braga, J.C., and Iryu, Y., Palaeobiogeographic patterns of a persistent monophyletic lineage: Lithophyllum pustulatum species group (Corallinaceae, Corallinales, Rhodophyta), Palaeogeogr., Palaeoclimatol., Palaeoecol., 2009, vol. 284, pp. 237–245.
Behforouzi, E. and Safari, A., Biostratigraphy and paleoecology of the Qom Formation in Chenar area (northwestern Kashan), Iran, Rev. Mex. Cienc. Geol., 2011, vol. 28, pp. 555–565.
Bozorgnia, F., Qum Formation stratigraphy of the Central Basin of Iran and its intercontinental position, Bull. Iran Pet. Inst., 1966, vol. 24, pp. 69–75.
Cahuzac, B. and Poignant, A., Essai de biozonation de l’Oligo-Miocène dans les bassins européens à l’aide des grands foraminifères néritiques, Bull. Soc. Geol. Fr., 1997, vol. 168, pp. 155–169.
Cornacchia, I., Agostini, S., and Brandano, M., Miocene oceanographic evolution based on the Sr and Nd isotope record of the Central Mediterranean, Paleoceanogr., Paleoclimatol., 2018, vol. 33, pp. 1–17.
Daneshian, J. and Aftabi, A., Foraminiferal biostratigraphy of the Qom Formation on the basis of new investigation at Navab anticline, in southeast Kashan, J. Sci. Univ. Tehran, 2010, vol. 35, pp. 137–157 (in Persian).
Daneshian, J. and Deziani, S., Study of Foraminifera biostratigraphy of the Qom Formation in stratigraphical section of Sorkhdeh section, Southeast Ashtian, J. Sci. I. R. I., 2004, vol. 14, no. 53, pp. 4571–4593 (in Persian).
Daneshian, J. and Ramezani Dana, L., Early Miocene benthic foraminifera and biostratigraphy of Qom Formation, Deh Namak, Central Iran, J. Asian Earth Sci., 2007, vol. 29, pp. 844–858.
Daneshian, J. and Ramezani Dana, L., Lower Miocene uncoiled Agglutinated Foraminifera from Dobaradar Section, southeast Qom, J. Strat. Sediment Res., 2016a, vol. 31, pp. 51–68 (in Persian).
Daneshian, J. and Ramezani Dana, L., Non spiral hyaline foraminifera of the Qom Formation, in type area, north of Central Iran, Earth Sci. Res., 2016b, vol. 7, pp. 126–148 (in Persian).
Daneshian, J. and Ramezani Dana, L., Foraminiferal biostratigraphy of the Miocene Qom Formation, northwest of the Qom, Central Iran, Front. Earth Sci., 2017, vol. 12, pp. 1–15.
Daneshian, J. and Ramezani Dana, L., Benthic foraminiferal events of the Qom Formation in the north Central Iran Zone, Paleontol. Res., 2019, vol. 23, no. 1, pp. 10–22.
Daneshian, J., Ramezani Dana, L., and Sadler, P., A composite foraminiferal biostratigraphic sequence for the Lower Miocene deposits in the type area of the Qom Formation, central Iran, developed by constrained optimization (CONOP), J. Afr. Earth Sci., 2017, vol. 125, pp. 214–229.
Daneshian, J., Shahrabi, M., and Akhlaghi, M., Biostratigraphy and Paleoenvironment of Qom Formation in northeast of Mahneshan, J. Geosci., 2010, vol. 19, no. 76, pp. 45–50 (in Persian).
Daneshian, J., Mosadegh, H., Khalaj, H., and Ghasemi, A., Sequence stratigraphy of the Qom Formation at type area (Kuh-e-Bichareh section), in southeast Qom, north of Central Iran, J. Sci. Univ. Tehran, 2008, vol. 34, pp. 19–54 (in Persian).
de Verteuil, L. and Norris, G., Miocene Dinoflagellate Stratigraphy and Systematics of Maryland and Virginia, Micropaleontol. Press, 1996.
Dozy, J.J., Comments on geological report no. 1, by Thiebaud (on Qum-Saveh area), Geol. Rep., 1944, no. 308.
Dybkjær, K. and Piasecki, S., Neogene dinocyst zonation for the eastern North Sea Basin, Denmark, Rev. Palaeobot. Palynol., 2010, vol. 161, pp. 1–29.
Dybkjær, K. and Rasmussen, E.S., Dinocyst stratigraphy in an expanded Oligocene-Miocene boundary section in the eastern North Sea Basin (the Frida-1 well, Denmark) and correlation from basinal to marginal areas, J. Micropalaeontol., 2007, vol. 26, pp. 1–17.
Filatoff, J. and Price, P.L., A pteridacean spore lineage in the Australian Mesozoic, in Palynological and palaeobotanical studies, Balme, B.E., Jell, P.A., and Playford, G., Eds., Memoirs of the Association of Australasian Palaeontologists, 1988, vol. 5, pp. 89–124.
Furrer, M.A. and Soder, P.A., The Oligo-Miocene marine formation in the Qom region (Central Iran), in Proc. 4th World Petr. Cong., Rome, 1955, pp. 267–277.
Hadavi, F., Notghi-Moghaddam, M., and Mousazadeh, H., Burdigalian-Serravalian calcareous nannoplanktons from Qom Formation, North-Center Iran, Arab. J. Geosci., 2010, vol. 3, pp. 133–139.
Hajian, J., Geological Map of Farmahin (1/100000), Geol. Surv. Iran, 1970.
Harzhauser, M., Strombus (Dilatilabrum) roegli sp. nov., a giant Oligocene strombid (Gastropoda) and its bearing on palaeoclimatic reconstructions of the Western Tethys, Ann. Naturhist. Mus. Wien, 2001, vol. 102A, pp. 51–67.
Hassani, M.J. and Hosseinipour, F., Quantitative analysis, basin evolution and paleoecology of Early Miocene ostracods in the southwest of Kerman, Iran, Geopersia, 2018, vol. 8, no. 2, pp. 213–232.
Hassani, M.J. and Vaziri, M.R., Early-Miocene gastropods from Khavich area, south of Sirjan, (Kerman, Iran): Biostratigraphy, paleogeography and paleoecology, J. Sci. Islamic Repub. Iran, 2011, vol. 22, no. 2, pp. 125–133.
Heilmann-Clausen, C. and Costa, L.I., Dinoflagellate zonation of the uppermost Paleocene? to lower Miocene in the Wursterheide Research Well, NW Germany, Geol. Jahrb., 1990, vol. 111, pp. 431–521.
Heydari, E., Hassanzadeh, J., Wade, W.J., and Ghazi, A.M., Permian-Triassic boundary interval in the Abadeh section of Iran with implications for mass extinction, Part 1: Sedimentology, Palaeogeogr., Palaeoclimatol., Palaeoecol., 2003, vol. 193, pp. 405–423.
Jafar-Nezhad, A.A., Ghasemi-Nejad, E., Mohtat, T., and Aghanabati, A., Paleogene dinoflagellate cysts and thermal maturity from Pabdeh Formation (Zagros basin, west of Iran), J. Appl. Sci. Environ. Manag., 2015, vol. 19, no. 3, pp. 353–358.
Karevan, M., Vaziri-Moghaddam, H., Mahboubi, A., and Moussavi-Harami, R., Biostratigraphy and paleo-ecological reconstruction on Scleractinian reef corals of Rupelian-Chattian succession (Qom Formation) in northeast of Delijan area, Geopersia, 2014, vol. 4, pp. 11–24.
Khaksar, K. and Maghfouri-Moghadam, I., Paleontological study of the echinoderms in the Qom Formation (Central Iran), Earth Sci. Res. J., 2007, vol. 11, no. 1, pp. 57–79.
Köthe, A., Dinozysten-Zonierung im Tertiär Norddeutschlands, Rev. Paléobiol. Genève, 2003, vol. 22, pp. 895–293.
Laursen, G.V., Monibi, S., Allan, T.L., Pickard, N.A., Hosseiney, A., Vincent, B., Hamon, Y., van Buchem, F.H., Moallemi, A., and Driullion, G., The Asmari Formation revisited: Changed stratigraphic allocation and new biozonation, in Shiraz First Int. Petrol. Conf. and Exhibition, Iran, 2009, vol. 4–6.
Maghfouri-Moghadam, I., Initial Identification of Early Miocene Ostracoda (Qom Formation) in West of Ashtian, Central Iran, Strat. Paleontol. Q. J., 2015, vol. 1, no. 20, pp. 1–14 (in Persian).
Maghfouri-Moghadam, I. and Yasboulaghi, S., Small pelecypoda of the Qom Formation (Early Miocene) in west of Ashtian, Central Iran, J. Geosci., 2015, vol. 24, no. 95, pp. 275–280 (in Persian).
Maghfouri-Moghadam, I., Borji, S., Amini, E., Azadbakht, S., and Taherpour Khalil Abad, M., Microbiostratigraphy of the Qom Formation in South and West of Tafresh, Central Iranian, J. Earth Sci., 2014, vol. 6, pp. 51–62.
Mao, S. and Lei, Z., Tertiary dinoflagellates and environmental evolution of Pearl River Mouth Basin, South China Sea, in Research on Micropalaeontology and Palaeoceano-graphy in Pearl River Mouth Basin, South China Sea, Hao, Y.C., Xu, Y.L., and Xu S.C., Eds., China, Wuhan: China Univ. Geosci. Press, 1996, pp. 52–66.
Mao, S., Wu, G., and Li, J., Oligocene-early Miocene dinoflagellate stratigraphy, Site 11448, ODP Leg 184, South China Sea, Prell, W.L., Wang, P., Blum, P., Rea D.K. and Clemens, S.C., Eds., Proceedings of the Ocean Drilling Program, Scientific Results, College Station, TX (O.D.P.), 2004, pp. 1–29.
Mohammadi, E. and Ameri, H., Biotic components and biostratigraphy of the Qom Formation in northern Abadeh, Sanandaj-Sirjan fore-arc basin, Iran (northeastern margin of the Tethyan Seaway), Arab. J. Geosci., 2015, vol. 8, pp. 10789–10802.
Mohammadi, E., Vaziri, M., and Dastanpour, M., Biostratigraphy of the nummulitids and lepidocyclinids bearing Qom Formation based on larger benthic foraminifera (Sanandaj-Sirjan fore-arc basin and Central Iran back-arc basin, Iran), Arab. J. Geosci., 2015, vol. 8, no. 1, pp. 403–423.
Mohammadi, E., Safari, A., Vaziri-Moghaddam, H., Vaziri, M.R., and Ghaedi, M., Microfacies analysis and paleoenvironmental interpretation of the Qom Formation, south of the Kashan, Central Iran, Carbonates Evaporites, 2011, vol. 26, pp. 255–271.
Mohammadi, E., Hasanzadeh-Dastgerdi, M., Ghaedi, M., Dehghan, R., Safari, A., Vaziri-Moghaddam, H., Baizidi, Ch., Vaziri, M.A., and Sfidari, E., The Tethyan Seaway Iranian Plate Oligo-Miocene deposits (the Qom Formation): Distribution of Rupelian (Early Oligocene) and evaporite deposits as evidences for timing and trending of opening and closure of the Tethyan Seaway, Carbonates Evaporites, 2013, vol. 28, pp. 321–345.
Munsterman, D.K. and Brinkhuis, H., A Southern North Sea Miocene dinoflagellate cyst zonation, Netherlands J. Geosci./Geol. Mijnbouw, 2004, vol. 83, no. 4, pp. 267–285.
Noroozpour, H., Microbiostratigraphy and paleoecology of the Qom Formation in the Goylar section, southwest of Zanjan Province, J. Adv. Appl. Geol., 2020, vol. 10, no. 2, pp. 224–234 (in Persian).
Nouradini, M., Yazdi, M., and Ashouri, A., Systematic notes on Burdigalian Echinoids from the Qom Formation in the Bagher Abad area, Central Iran, Geopersia, 2014, vol. 4, no. 2, pp. 155–167.
Nouradini, M., Azami, S.H., Hamad, M., Yazdi, M., and Ashouri, A.R., Foraminiferal paleoecology and paleoenvironmental reconstructions of the lower Miocene deposits of the Qom Formation in Northeastern Isfahan, Central Iran, Bol. Soc. Geol. Mex., 2015, vol. 67, no. 1, pp. 59–73.
Parandavar, M. and Hadavi, F., Identification of the Oligocene–Miocene boundary in the Central Iran Basin (Qom Formation): calcareous nannofossil evidences, Geol. Q., 2019, vol. 63, no. 2, pp. 215–229.
Pedramara, A., Zágorše, K., Bitner, M.A., Yazdi, M., Bahrami, A., and Maleki, Z., Bryozoans and brachiopods from the Lower Miocene deposits of the Qom Formation in North-East Isfahan (Central Iran), Neues Jahr. Geol. Paläontol, Abh., 2019, vol. 294, no. 2, pp. 229–250.
Popov, S.V., Rögl, F., Rozanov, A.Y., Steininger, F.F., Shcherba, I.G., and Kovac, M., Lithological-paleogeographic maps of Paratethys: 10 late Eocene to Pliocene, Cour. Forschungsinst. Senckenberg, 2004, vol. 250, pp. 1–46.
Powell, A.J. and Brinkhuis, H., Neogene Time Scale–Dinoflagellate stratigraphy, in A Geological Time Scale, Gradstein, F.M., Ogg, J., and Smith, A., Eds., Cambridge: Cambridge Univ. Press, 2004, pp. 409–440.
Reuter, M., Piller, W.E., Harzhauser, M., Mandic, O., Berning, B., Rögl, F., Kroh, A., Aubry, M.P., Wielandt-Schuster, U., and Hamedani, A., The Oligo-Miocene Qom Formation (Iran): Evidence for an early Burdigalian restriction of the Tethyan seaway and closure of its Iranian gateways, Int. J. Earth Sci., 2009a, vol. 98, pp. 627–650.
Reuter, M., Piller, W.E., and Harzhauser, M., Comment on “Revision of the age of the Qom Formation in the Central Iran Basin, Iran”, Zhu et al., Ed., J. Asian Earth Sci., 2007, vol. 29, pp. 715–721; J. Asian Earth Sci., 2009b, vol. 34, pp. 699–701.
Soliman, A., Ćorić, S., Head, M.J., Piller, W.E., and El Beialy, S.Y., Lower and Middle Miocene biostratigraphy, Gulf of Suez, Egypt based on dinoflagellate cysts and calcareous nannofossils, J. Palynol., 2012, vol. 36, no. 1, pp. 38–79.
Stӧcklin, J. and Setudehnia, A., Stratigraphic lexicon of Iran Ministry of Industry and Mines, Geol. Surv. Iran, 1991, Report no. 18, pp. 1–376.
Traverse, A., Paleopalynology, Dordrecht, Netherlands: Springer, 2007.
Van Buchem, F.S.P., Allan, T.L., Laursen, G.V., Lotfpour, M., Moallemi, A., Monibi, S., Motiei, H., Pickard, N.A.H., Tahmasbi, A.R., Vedrenne, V., and Vincent, B., Regional stratigraphic architecture and reservoir types of the Oligo-Miocene deposits in the Dezful Embayment (Asmari and Pabdeh Formations) SW Iran, Geol. Soc. Spec. Publ., 2010, vol. 329, pp. 219–263.
Wade, B.S., Pearson, P.N., Berggren, W.A., and Pälike, H., Review and revision of Cenozoic tropical planktonic foraminiferal biostratigraphy and calibration to the geomagnetite polarity and astronomical time scale, Earth Sci. Rev., 2011, vol. 104, pp. 111–142.
Wynd, J., Biofacies of the Iranian consortium agreement area, IOOC., 1965, Report no. 1082.
Yazdi, M., Parvanenejad Shirazi, M., Rahiminejad, A.H., and Motavalipoo, R., Paleobathymetry and paleoecology of colonial corals from the Oligocene–early Miocene (?) Qom Formation (Dizlu area, central Iran), Carbonates Evaporites, 2012, vol. 27, pp. 395–405.
Yazdi-Moghaddam, M., Early Oligocene larger foraminiferal biostratigraphy of the Qom Formation, South of Uromieh (NW Iran), Turk. J. Earth Sci., 2011, vol. 20, pp. 847–856.
Yazdi-Moghadam, M., Sadeghi, A., Adabi, M.H., and Tahmasbi, A., Foraminiferal biostratigraphy of the lower Miocene Hamzian and Arashtanab sections (NW Iran), northern margin of the Tethyan Seaway, Geobios, 2018a, vol. 51, pp. 231–246.
Yazdi-Moghadam, M., Sadeghi, A., Adabi, M.H., and Tahmasbi, A., Stratigraphy of the Lower Oligocene nummulitic limestones, north of Sonqor (NW Iran), Riv. Ital. Paleontol. Strat., 2018b, vol. 124, no. 2, pp. 407–419.
Yazdi-Moghaddam, M., Sarfi, M., Ghasemi-Nejad, E., Sadeghi, A., and Sharifi, M., Early Miocene larger benthic foraminifera from the northwestern Tethyan Seaway (NW Iran): New findings on Shallow Benthic Zone 25, Int. J. Earth Sci., 2021, vol. 110, pp. 719–740.
Zhu, Y., Qi, Y., Zhang, B., Yang, H., He, Ch., Wang, Sh., Zhou, W., Zhu, Q., and Li, Zh., Revision of the age of the Qom Formation in the Central Iran Basin, Iran, J. Asian Earth Sci., 2007, vol. 29, pp. 715–729.
ACKNOWLEDGMENTS
This article benefited greatly from the collaboration of Hassan Mafi during the field work. We would like to extend our sincere thanks to Popov Sergey for his constructive comments.
Funding
The authors did not receive support from any organization for the submitted work.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The authors declare that they have no conflicts of interest.
Additional information
Publisher’s Note.
Pleiades Publishing remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Afsaneh Dehbozorgi, Saeed Maleki Palynostratigraphy and Foraminiferal Stratigraphy of the Early Miocene Qom Formation in the Northeastern Margin of the Tethyan Seaway. Paleontol. J. 57, 833–847 (2023). https://doi.org/10.1134/S0031030123070031
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0031030123070031