
Abstract
Qom Formation in the Sanandaj–Sirjan fore-arc and Central Iran back-arc basins was studied in order to determine its accurate age. Larger benthic foraminifera (mainly Lepidocyclinidae, Nummulitidae, and Neoalveolina) are biostratigraphically the main biota of the Qom Formation in the study areas. There are some debates concerning the age dating of the Qom Formation layers based on the presence or absence of Nummulites and Eulepidina. However, the last true Nummulites spp. became extinct at the top of Late Rupelian and all of the in situ Nummulites bearing layers of the Qom Formation belong to Rupelian in age. Lepidocyclinids, which originated in the Americas during the Middle Eocene, reached Iran in the Rupelian. The first appearance of Lepidocyclina spp. in our study sections, before the last occurrence of Nummulites spp., clearly indicates that the earliest Lepidocyclinidae reached Iran in the Rupelian. The Lepidocyclinids are widespread in the Chattian marine deposits of Iran. The presence of in situ Nummulites (with or without Eulepidina) in Qom Formation successions indicates an Early Oligocene (Rupelian) age, and the presence of Eulepidina without Nummulites suggests a Late Oligocene age (Chattian; possibly into Aquitanian). Moreover, Nummulites intermedius-Nummulites vascus Assemblage zone of Wynd (1965) and Eulepidina-Nephrolepidina-Nummulites Assemblage zone of Adams and Bourgeois (1967), which were proposed for Oligocene (Rupelian- Chattian), should be ascribed to Rupelian in age. On the basis of biostratigraphy data and foraminifera assemblages, the Qom Formation is Rupelian-Chattian, Rupelian, and Rupelian-Burdigalian in age in Bujan, Varkan, and Qom (Khurabad) areas, respectively.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The marine Qom Formation was deposited at the north-eastern coast of the Tethyan Seaway (Reuter et al. 2009b), in the Oligo-Miocene, during the final sea transgression (Rahimzadeh 1994; Daneshian and Ramezani Dana 2007; Khaksar and Maghfouri–Moghadam 2007), in the Sanandaj–Sirjan fore-arc, Urumieh–Dokhtar magmatic arc (Intra-arc), and Central Iran back-arc basins (Mohammadi et al. 2013; Fig. 1). It is essential and important to study different properties of the oil-bearing Qom Formation because of economic importance and communicative role between Eastern Tethys (the proto-Indian Ocean) and the Western Tethys region (the proto-Mediterranean Sea) in the Iranian Plate at the same time (Mohammadi et al. 2011, 2013).

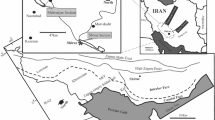
A map showing (1) distribution of Thetyan Seaway Iranian Plate Oligo-Miocene deposits (Qom Formation); (2) distribution of Urumieh-Dokhtar Magmatic Arc (UDMA); (3) distribution of the Qom Formation evaporate deposits: deposition of evaporate deposits of the Qom Formation took place in concave side of the red line; and (4) time and trend of opening of the Tethyan Seaway on the Iranian plate. As indicated on the map, the Rupelian deposits of the Qom Formation deposited below 34° 20′ N (blue line); the Chattian deposits continued to 35° N (green line) and above 35° N, deposition of the Qom Formation started in Miocene. The violent arrows show the direction of the transgression of the Tethyan Seaway on the Iranian plate; as indicated on the map transgression of the Tethyan Seaway on the Iranian plate started from southeast and continued northwestward gradually. The main cities and other places mentioned in the text and Table 1 are shown on this map (Mohammadi et al. 2013)


Co-occurrence of Lepidocyclinids and Nummulites spp. in the study sections clearly indicates that the Lepidocyclinids reached Iran in the Rupelian (Early Oligocene). a–d Khurabad section, e–f Varkan section
Biogenic components of the Oligo-Miocene Qom formation comprise of different biota including: larger benthic foraminifera (LBF) which are represented by nummulitids (Nummulites, Operculina, Heterostegina, Spiroclypeus), lepidocyclinids (Eulepidina, Nephrolepidina, Lepidocyclina), alveolinids (Borelis), miogypsinids (Miogypsinoides, Miogypsina), Amphistegina, Archaias, Austrotrillina, small benthic foraminifera (textulariids, rotaliids, miliolids), planktic foraminifera, corals, corallinacean algae, bryozoans, bivalves, echinoderms, gastropods, and polychaete worm tubes especially Ditrupa. Among the biogenic components, LBF, which are the main biota of the Qom Formation in the study areas, are biostratigraphically important fossils in biozonation because of these episodes of rapid diversification and abrupt extinction (Hallock 1985; Beavington-Penney and Racey 2004).
The Foraminifera is a class of microscopic shelled protists. Foraminifers are the most abundant shelled microorganisms in benthic marine environments (Koukousioura et al. 2011). Simple forms of foraminifera appeared in the Cambrian and were common in the Early Palaeozoic. They became abundant, with the evolutionary development of relatively large and complicated test architecture, by the Late Palaeozoic, which provide a model example of evolutionary diversity throughout the Mesozoic and Cenozoic. This long and well-recorded evolutionary record makes foraminifera of outstanding value in zonal stratigraphy and palaeoenvironmental, palaeobiological, and palaeoceanographic interpretation and analysis (BouDagher-Fadel 2008). Foraminifers that host algal endosymbionts are informally referred to as LBF (Baker et al. 2009). LBF are typically larger than 2 mm in diameter and 3 mm3 in volume and have complex internal structures which, when studied in thin section, are useful for the biostratigraphy of Tethyan and other tropical limestones (Armstrong and Brasier 2005). The tests of dead, larger foraminifera are important rock-forming materials, and foram-limestones are extensively developed in the Upper Palaeozoic, the Upper Cretaceous, and in the Cenozoic. From their structural complexity, and because of the diversity of the shelf environments that they inhabited, fossil larger foraminifera provide unique information on palaeoenvironments and biostratigraphy of shelf limestones around the world (BouDagher-Fadel 2008). These protists are important as biostratigraphical indicators in marine rocks of Late Palaeozoic, Mesozoic, and Cenozoic ages because they are abundant, diverse, and easy to study (Armstrong and Brasier 2005). The larger foraminifera assemblage zones could be identified by the presence of a few key taxa, usually with the use of a hand lens in the field. Some groups of larger foraminifera provide excellent biostratigraphic markers, sometimes the only ones which can be used to date carbonate successions (e.g., the fusulinines in the Upper Palaeozoic, orbitoidids in the Middle to Upper Cretaceous, nummulitids in the Palaeogene, and lepidocyclinids and miogypsinids in the Oligocene and Neogene (BouDagher-Fadel 2008). Besides, foraminifera are typically a significant component of marine benthic communities, and can provide sensitive indicators of environmental conditions because of their short reproductive cycles, great abundance in marine and estuarine habitats, good test preservation (Koukousioura et al. 2011), stress tolerance, small size, and high diversity (Martínez-Colón et al. 2009).
Eocene/Oligocene boundary belongs to the most significant extinction events in the Phanerozoic (Nebelsick et al. 2005). Oligocene carbonate and mixed siliciclastic–carbonate platforms are characterized by the re-establishment of shallow water marine benthic communities following major changes at the Eocene/Oligocene boundary. The Oligocene epoch represents a time span dominated by larger foraminiferal and coralline algal facies in carbonate environments (Bassi and Nebelsick 2010). As biostratigraphic markers, various LBF are also key organisms for dating shallow water successions using the shallow benthic zones which have also been introduced for the Oligocene and Miocene (Cahuzac and Poignant 1997; Bassi and Nebelsick 2010).
According to next part and as shown in Table 1, mostly all of the previous works on the Qom Formation are limited to the middle parts of Iran. However, surprisingly little information is available concerning the different geological and paleontological properties (biostratigraphy, paleoenvironment, etc.) of the Qom Formation in the southeast of Iran. Besides, mostly all of the previous works are limited to one of the Sanandaj-Sirjan, Urumieh–Dokhtar, and Central Iran basins, and there are a few works that have compared different properties of the Qom Formation in the different basins. The aim of this study is therefore to bridge this gap by study and comparison of some stratigraphic sections in the Sanandaj-Sirjan and Central Iran basins in order to analyze their biostratigraphy, on the basis of LBF.
Previous works
Geological investigations of the Qom Formation started with the work of Loftus (1855) and continued by Tietze (1875), Stahl (1911), and Kuhn (1933). After oil was discovered in porous bioclastic limestones in 1934, the Qom Formation became the focus of scientific research as different properties of the Qom Formation have been studied by many authors (e.g., Loftus 1855; Tietze 1875; Stahl 1911; Kuhn 1933; Riben 1935; Furon and Marie 1939; Furon 1941; Dozy 1945, 1955; Furrer and Soder 1955; Gansser 1955; Abaie et al. 1963, 1964; Bozorgnia 1966; Rahaghi 1973, 1980). More recently, published researches on the Qom Formation were done by many authors which are listed in Table 1. Besides, Harzhauser (2000) and Mandic (2000) studied the gastropods and pectinid bivalves of the Qom Formation, respectively. However, both are unpublished PhD theses.
Mohammadi et al. (2013) have illustrated distribution of the Qom Formation on new maps showing the outcrops of the Qom Formation extended to east and southeast of the Sirjan depression in the southeast, northern, and southern of Great Kavir in the east, foothills of the Alborz Range in the north and northeast, north of Orumieh Lake (Khoy and Maku) in the northwest, and the southwest limit of the Qom Formation is coincident along with Orumieh–Mahabad–Saqqez–Hamadan–Esfahan–Abadeh–Sirjan–Jazmurian trend (Fig. 1).
Mohammadi et al. (2013) by scrutinizing (1) two stratigraphic sections of the Qom Formation in the southern and southwestern Kashan, (2) results of the study of more than 100 stratigraphic sections of the Qom Formation, (3) geological maps of Iran sheets, and (4) more than 30 geological maps of different parts of Iran, illustrated the distribution of Rupelian (Early Oligocene) and evaporate deposits as evidence for timing and trending of opening and closure of the Tethyan Seaway. Their study area encompasses more than 1,700 km (aerial distance) of the north-eastern coast of the Tethyan Seaway. They concluded that the Qom Formation is Rupelian–Burdigalian in age. Besides, Mohammadi et al. (2013) believed that evaporate deposits of the Qom Formation are deposited in a rather small area of Central Iran back-arc basin and are deposited totally in the Early Miocene (Aquitanian–Burdigalian). This indicates that due to the compressive tectonic regime in the Central Iran back-arc basin, the gates to the open ocean became restricted in the Early Miocene gradually (Reuter et al. 2009b).
Geological setting
Iran is divisible into eight geological provinces. These provinces are: (1) Zagros, (2) Sanandaj–Sirjan, (3) Urumieh–Dokhtar (Sahand–Bazman) magmatic arc, (4) Central Iran, (5) Alborz, (6) Kopeh Dagh, (7) Lut, and (8) Makran accretionary prism, each province having experienced distinct tectonic and sedimentary histories (Heydari et al. 2003). The subduction of the Neo-Tethyan ocean floor beneath Iran (Late Cretaceous–Early Paleocene) sutured Iran to Arabia (Berberian and King 1981; Alavi 2004; Ghasemi and Talbot 2006), and the subsequent continental convergence built the Zagros Orogenic Belt. This orogenic belt consists of four NW–SE trending parallel zones: (1) Urumieh–Dokhtar Magmatic Assemblage (UDMA), (2) Sanandaj–Sirjan Zone (SSZ), (3) High Zagros, and (4) Zagros Simply Folded Belt (Ghasemi and Talbot 2006; Shafiei et al. 2011). The SSZ extends for 1,500 km from Sanandaj in the NW to Sirjan (Shafiei et al. 2011). The UDMA forms a distinct linear intrusive–extrusive complex, which extends along the entire length of Zagros orogen, with a width of over 4 km (Ghasemi and Talbot 2006). Central Iran is separated from the SSZ by UDMA and a belt of steep and straight faults (Ghasemi and Talbot 2006).
The tectonic units of Central Iran originated during the subduction and final collision of the African/Arabian with the Iranian Plate (Schuster and Wielandt 1999). Northeastward subduction of the African–Arabian Plate underneath the Eurasian plates led to the formation of a volcanic arc (Urumieh–Dokhtar belt) during the Cretaceous to Eocene and compartmentalization of the region, creating the Esfahan–Sirjan (Sanandaj–Sirjan Zone) fore-arc and the Qom or Central Iran back-arc basins at the north-eastern margin of the Tethyan Seaway (Reuter et al. 2009b; Berning et al. 2009). The Eocene period of volcanism in Central Iran was followed by Late Eocene (37 Ma) movements, represented by a regional unconformity at the base of the Oligocene rocks. During this phase, the Lut zone in east-Central Iran underwent uplift (major Lut uplift), and no Oligocene–Miocene sediments were apparently deposited (Berberian and King 1981). Orogenic movements in the Late Eocene–Early Oligocene resulted in the creation of lagoonal to continental sedimentary regime characterized by detritic-evaporitic sediments of the Lower Red Formation (LRF; Khaksar and Maghfouri–Moghadam 2007). Following a short interval of intermediate to acid volcanic to subvolcanic activity and before final closure of the seaway during the latest Early Oligocene–latest Early Miocene, thick limestone and marl successions (the Qom Formation) were formed in three NW–SE-trending basins: Sanandaj–Sirjan (fore-arc basin), Central Iran (back-arc basin; Gilg et al. 2006; Berning et al. 2009), and Urumieh–Dokhtar magmatic arc (intra-arc basin (Mohammadi et al. 2013). The Qom limestones are followed by thick continental red beds [the Upper Red Formation (URF)] with intercalated marine (to lacustrine?) evaporate deposits (gypsum and salt; Gilg et al. 2006). Rahimpour-Bonab and Kalantarzadeh (2005) believed that in the Early–Middle Miocene, development of restricted marine conditions led to a facies change from shelf carbonates of the Qom Formation to the evaporate series of the M1 member of the overlying URF. The URF consists of sandstones, marls, and inter-layered evaporates which mostly are present in the lower parts.
Study area and methods
Three stratigraphic sections of the Qom Formation were measured and sampled bed by bed in order to analyze their biostratigraphic conditions. They included (1) the Bojan section with these coordinates: 29° 26′ 04″ N, 55° 59′ 27″ E, about 40 km eastern Sirjan City, (2) Varkan section with these coordinates: 33° 41′ 29″ N, 51° 04′ 54″ E, about 75 km southwestern Kashan City, and (3) Khurabad (Khourabad in Mohammadi et al. 2013) section with these coordinates: 34° 30′ 53″ N, 50° 56′ 58″ E, about 20 km southeastern Qom City (Fig. 1). Based on Schuster and Wielandt (1999), Reuter et al. (2009b), Berning et al. (2009), Mohammadi et al. (2013) geological maps of Sirjan (Soheili 1995), Kashan (Zahedi and Amidi 1991), and Qom (Hajian and Emammi 1991) and Geological Maps of Iran Sheets (no. 1, 2, and 5; NIOC 1969, 1978, 1977), Bojan and Varkan sections are located in the Sanandaj-Sirjan fore-arc basin and Qom section is located in Central Iran back-arc basin (Fig. 1). Outcrops of the Qom Formation in Bujan area with 156-m thickness mainly consist of medium to thick bedded and massive limestone, shale, and marl (Fig. 4). They lay unconformably on top of the polygenetic conglomerate with indeterminate age (Fig. 4), and their upper boundary is covered by quaternary alluvium. The Qom Formation Outcrops in the Varkan area, with 190-m thickness, consist mainly of medium to thick bedded and massive limestone, marly limestone, marl, and conglomerate (Fig. 6). They lay unconformably on top of the LRF, and their upper boundary is covered with polymictic Pliocene conglomerate (Fig. 6). Outcrops of the Qom Formation in Qom area (Khurabad), with 260-m thickness, consist mainly of medium to thick bedded and massive limestone, marly limestone, nodular, limestone, shale and marl, sandstone, conglomerate, and evaporate deposits (Fig. 9). They lay on top of the Eocene volcanic rocks, and their upper boundary is covered with URF (Fig. 9). Field and microscopic studies were carried out for biostratigraphical analysis of the studied sections. Up to 330 samples were collected based on field evidences and lithofacies changes. Thin sections were obtained from harder samples, while softer samples were disaggregated and their microfauna were processed, and finally thin sections and released specimens were analyzed. All samples were studied in detail, and particular attention has been paid to the larger benthic foraminiferal associations, since they are excellent bio-indicators for age dating and paleoenvironmental interpretation. Obtained foraminifera were identified based on different studies by Stainforth et al. (1975), Rahaghi (1980), Loeblich and Tappan (1988), Boudagher-Fadel and Lord (2000), Boudagher-Fadel and Lokier (2005), Iaccarino and Premoli Silva (2005), Sharaf et al. (2005), BouDagher-Fadel (2008), Kuss and Boukhary (2008), Özcan et al. (2009), and BouDagher-Fadel and Price (2010). The fossil contents of the study section are so well preserved, abundant, and diverse that these conditions improved this investigation’s process. The last occurrence of the Nummulites, the time range of Lepidocyclinids (genera/subgenus), the first appearance, and last occurrence of Lepidocyclinids in Iran are studied and analyzed based on several previous researches and are shown in different Tables in this paper. Besides, age of the study sections is determined according to several authors including: Wynd (1965), Adams and Bourgeois (1967), Rahaghi (1980), Adams (1984), Loeblich and Tappan (1988), Racey (1995), Cahuzac and Poignant (1997), Beavington-Penney and Racey (2004), Harzhauser (2004), Ehrenberg et al. (2007), BouDagher-Fadel (2008), Laursen et al. (2009), BouDagher-Fadel and Price (2010), Van Buchem et al. (2010), and Mohammadi et al. (2013).

Biostratigraphy of the Qom Formation at the Bujan section (S Sirjan) based on key LBF

A figure showing abundance of N. fitchteli/N. intermedius in hand sample and thin section of the Bujan section

Biostratigraphy of the Qom Formation at the Varkan section (SW Kashan) based on key LBF

Selected key LBF of the Qom Formation in study sections. a–d N. fichteli; Bujan section, e–g N. vascus; e, g Varkan section; f Khurabad section; h–j Eulepidina sp., Bujan section; k, l Lepidocyclina (Nephrolepidina) sp., Khurabad section

Selected key LBF of the Qom Formation in study sections. a–b Miogypsina sp, Khurabad section; c (Nu) N. vascus (Am) Amphistegina, Khurabad section; d Operculina sp., Bujan section; e Heterostegina sp., Bujan section; f Borelis pygmaea, Bujan section; g–j Borelis melo curdica, Khurabad section

Biostratigraphy of the Qom Formation at the Khurabad section (SE Qom) based on key LBF
Discussion
There is no formal biozonation for the Qom Formation. However, conspicuous similarity is observed between the Qom Formation foraminifera and those of the Asmari Formation (Zagros region, SW Iran) by many authors correlating the Qom Formation with the Asmari Formation (Bozorgnia 1965; Daneshian and Dana 2007; Mohammadi et al. 2011). LBF, which are the main biota of the Qom Formation in the study sections, are biostratigraphically important fossils in biozonation.
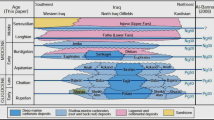
Biostratigraphy criteria of the Asmari Formation was established by Wynd (1965) and reviewed by (Adams and Bourgeois 1967); however, both are unpublished reports. Adams and Bourgeois (1967) designed four assemblage zones for the Asmari Formation (Fig. 2). They could not recognize the stages of the Oligocene as individual.
Biostratigraphically, Nummulites and Lepidocyclinids are more important for age dating of Rupelian, Chattian (and Aquitanian) deposits of the Qom, and Asmari formations. Racey (1994) believed that the presence of Nummulites indicates an Early Oligocene age, and the presence of Eulepidina without Nummulites suggests a late Oligocene age. But Eulepidina has been reported with Nummulites in Late Oligocene and without Nummulites in Early Miocene by some researchers (e.g., Bozorgnia 1966; Adams and Bourgeois 1967). Besides, using Sr isotope dating, Ehrenberg et al. (2007) and Laursen et al. (2009) believed that the last occurrence of Nummulites in the Asmari Formation was recorded before the end of Rupelian time. As cited above, there are some debates concerning the age dating of the Qom Formation layers based on the presence or absence of Nummulites and Eulepidina. Therefore, their ranges are discussed in the following sections (Tables 2, 3, and 4).
Nummulites range
The Nummulitidae are common throughout the Cenozoic, and most of them are extant. They are in fact the largest extant calcareous foraminifera (BouDagher-Fadel 2008). Living nummulitids are widely distributed in modern tropical and subtropical shallow-water seas and achieve their highest diversity in the subtropical and tropical West Pacific (BouDagher-Fadel 2008). Several nummulitids genera, especially Assilina and Nummulites, are particularly useful for zoning Tethyan shelf sequences (Haynes et al. 2010). Nummulites are good markers for Early Tertiary (Eocene, Oligocene) sedimentary sequences in the Tethyan realm. The species of this genus have evolved very rapidly and are classified into evolutionary groups which have common characteristic morphologic features that undergo changes during evolution (Boukhary and Scheibner 2009). In the Eocene, Nummulites and Assilina witnessed a major radiation and increase in test size, which persisted up to the major extinction of the last large species at the Middle–Late Eocene boundary. Few small species of Nummulites, but no Assilina, survive into the Late Eocene, and Nummulites finally became extinct in the end of Early Oligocene (BouDagher-Fadel 2008).
As shown in Table 2, all of the recent works (published since 2000) plus most of the older ones attribute the last occurrence of Nummulites to the Late Rupelian. For example, Beavington-Penney and Racey (2004) believed that from among the principal nummulitid genera, only Operculina, Palaeonummulites, Heterostegina, and Cycloclypeus have extant descendants; Nummulites became extinct in the Early Oligocene, Assilina in the late Middle Eocene, Ranikothalia in the late Palaeocene, and Spiroclypeus in the Early Miocene. It is worth mentioning that, some researchers (e.g., Hohenegger 2000) refer to one living species of Nummulites (Nummulites venosus) from the Pacific, although this has closer affinities with Operculinella, and following the more recent revised classification of Haynes et al. (2010) should be reassigned to Palaeonummulites. Besides, BouDagher-Fadel (personal communication 2011) believed that all megalospheric large Nummulites with large proloculus disappeared in the Late Rupelian. And that the small Nummulites (or Nummulites-like foraminifera) which are mainly called Paleonummulites/Operculinoides, etc., are still living today (BouDagher-Fadel, personal communication 2011; Mohammadi et al. 2013).
BouDagher-Fadel (2008) in her great book “Evolution and Geological Significance of Larger Benthic Foraminifera” noted that the range of Nummulites is from Middle Palaeocene to Early Oligocene. She believed that the extinction of Nummulites can be correlated with the Rupelian stage and planktonic zones P18-P21a. The last true Nummulites spp. became extinct at the top of the Td “Letter Stage” (Late Rupelian) with Nummulites fichteli from the upper Early Oligocene of Italy (BouDagher-Fadel 2008).
Lepidocyclinids ranges
Lepidocyclinids are important elements of Tertiary larger foraminiferal assemblages in Iran and in many other parts of the world. They originated in the Americas during the Middle Eocene and migrated eastward through the Tethyan/Mediterranean corridor and reached Iran in the Rupelian (BouDagher-Fadel 2008; BouDagher-Fadel and Price 2010). In Iran they have been recorded from sedimentary sequences of Early Oligocene (Rupelian) to Early Miocene times of the Qom and Asmari formations.
The LBF family Lepidocyclinidae has been described extensively in the literature, with occurrences ranging from the Middle Eocene in the Americas to the Late Miocene (or possibly Early Pliocene) in the Indo-Pacific Tethyan sub-province (BouDagher-Fadel and Price 2010; Table 3). Lepidocyclina (Lepidocyclina) of Tethys was almost certainly the direct descendant of Lepidocyclina (L.) of the Eocene and Oligocene to Early Miocene of Central America (BouDagher-Fadel 2008). By the study of new samples from 37 wells drilled off South America, and systematic comparisons of the Lepidocyclinidae from the South America and the Tethyan sub-provinces of the Mediterranean–West Africa and the Indo-Pacific, BouDagher-Fadel and Price (2010) traced the timing and route of the paleogeographic migration of the lepidocyclinids from the Americas, through the Tethyan/Mediterranean corridor, and into the Indo-Pacific region. These authors believed that Lepidocyclinids originated in the Americas during the Middle Eocene, Lutetian; but related forms are reported from West Africa from the Middle Eocene–Early Oligocene pointing to an eastward migration. They only reached the Mediterranean during the Early Oligocene, filling ecological niches that had previously been occupied in the Eocene by Nummulites. This migration continued within the Tethys until the lepidocyclinids finally colonized the Indo-Pacific in the late Early Oligocene (BouDagher-Fadel and Price 2010).
Iranian lepidocyclinids ranges
BouDagher-Fadel (2008) and BouDagher-Fadel and Price (2010) figured Lepidocyclina (Nephrolepidina) marginata (Michelotti) = L.(Nephrolepidina) tournoueri Lemoine, and Douvillé from Rupelian (Early Oligocene) deposits of Iran (Rahaghi NHM collection). Besides, there is clear evidence confirming that the first appearance of Lepidocyclina spp. in Iran occurred in the Rupelian (before the last occurrence of Nummulites spp.; see below).
The first appearance of Lepidocyclina spp. before the last occurrence of Nummulites spp. and co-occurrence of Lepidocyclinids and Nummulites spp. in the study sections (Bujan, Varkan, Khourabad) clearly indicate that the earliest Lepidocyclinidae reached Tethys (Iran) in the Rupelian (Fig. 3). This feature is corroborated by the senior author’s investigations from some other sections of the Qom Formation (e.g., Jazeh and Abadeh).
Schuster and Wielandt (1999) recorded the last occurrence of Nummulites at the end of the East Indian Letter Stage Td (in the Qom Formation deposits of Abadeh section). However, they recorded the first appearance of Lepidocyclina (Eulepidina) formosoides, Lepidocyclina (Eulepidina) favosa, and Lepidocyclina (Nephrolepidina) morgani) at the beginning of the East Indian Letter Stage Td. In other words, Schuster and Wielandt (1999) recorded the first appearance of Lepidocyclina before the last occurrence of Nummulites.
Similarly, in Oligo-Miocene Asmari Formation (SW Iran) the first appearance of Lepidocyclina spp. occurred before the last occurrence of Nummulites spp. in several stratigraphic sections (Table 4) including: (1) Lali section (N Ahwaz, Vaziri-Moghaddam et al. 2006), (2) Tang-e-Gurgdan (NW Gachsaran, Amirshahkarami et al. 2007a), (3) Tang-e Abolhayat (W Shiraz, Sadeghi et al. 2009), (4) Lali section (N Ahwaz, Sadeghi et al. 2009), (5) Firozabad section (SSW Shiraz, Kalanat et al. (2010), Vaziri-Moghaddam et al. 2011), (6) Rag-e-Safid oil field (SE Ahwaz, Amirshahkarami et al. 2010), and (7) Naura anticline (SE Shiraz, Sooltanian et al. 2011).
The first appearance of Lepidocyclinids before the last occurrence of Nummulites spp. in several stratigraphic sections of the Qom (Bujan, Jazeh, Khourabad, Varkan, Abadeh) and Asmari (Lali, Naura anticline, Tang-e Abolhayat, Tang-e-Gurgdan, Firozabad, Rag-e-Safid oil field) formations (Table 4) clearly indicates that the earliest Lepidocyclinidae reached Tethys (Iran) in the Rupelian (Early Oligocene).
Concerning the last occurrence of Lepidocyclina spp. in Iranian deposits (Table 5), it should be mentioned that Reuter et al. (2009b) recorded Lepidocyclina spp. from Aquitanian deposits of Qom, Zefreh, and Chalheghareh sections, and Burdigalian deposits of Zefreh section. Likewise, the last occurrence of Lepidocyclina spp. in Asmari Formation was recorded from Aquitanian deposits of Dehluran and Kabirkuh-Darrehshahr (by Vaziri-Moghaddam et al. 2010), and the Chattian deposits of several other stratigraphic sections (Khaviz Anticline, Rahmani et al. 2009; Lali section, Sadeghi et al. 2009; Tang-e Abolhayat, Sadeghi et al. 2009; Firozabad section, Vaziri-Moghaddam et al. 2011; Naura anticline, Sooltanian et al. 2011). It is worth mentioning that, Amirshahkarami et al. (2007a, b), recorded Lepidocyclinids from Aquitanian deposits of Tang-e-Gurgdan and Chaman-Bolbol sections, respectively (Asmari Formation), but considering new biozonation of the Asmari Formation, suggested by Laursen et al. (2009), their recorded Lepidocyclinids may be ascribed to Chattian in age.
As mentioned above, there are some debates concerning the age dating of the Qom Formation layers based on the presence/absence of Nummulites and Eulepidina. Racey (1994) believed that the presence of Nummulites indicates an Early Oligocene age, and the presence of Eulepidina without Nummulites suggests a Late Oligocene age. On the contrary, Eulepidina have been reported with Nummulites in Late Oligocene and without Nummulites in Early Miocene by many researchers (e.g., Bozorgnia 1966; Adams and Bourgeois 1967).
It must be mentioned that, Rahaghi (1980) believed that the Rupelian beds of the Qom Formation are characterized by the presence of Nummulites intermedius. He attributed the Qom Formation in Jovsheghan-e-ghali area (SW of Kashan) to Rupelian based on the presence of N. intermedius. It is worth mentioning that N. intermedius of the Jovsheghan-e-ghali area is co-occurred with Nephrolepidina sp. and Eulepidina aff. dilatata. Likewise, Harzhauser (2004) attributed the Nummulites bearing layers of the Qom Formation to Early Oligocene based on the occurrence of Nummulites sublaevigatus.
Concerning E. dilatata, it should be mentioned that Rahaghi (1980) recorded E. dilatata (without Nummulites) from Aquitanian deposits of Mashhad section (NW Kashan). Likewise, he believed that Borelis melo curdica is an index of Burdigalian age.
Besides, it is worth mentioning that Jones and Racey (1994) attributed the Early Oligocene age for Ashawq Formation (S Oman) based on occurrence of N. fichteli.
Biostratigraphy of study sections
In fact, Nummulites is an important Early Oligocene genus whose last occurrence marks the upper boundary of the Rupelian (Rahaghi 1980). Its last occurrence has been used (see Table 2) to mark the upper boundary of the Rupelian. Based on the extinction of the last true Nummulites spp. (Table 2) at the Late Rupelian, we believe that the presence of in situ Nummulites (with or without Eulepidina) in Qom Formation successions indicates Early Oligocene (Rupelian) age, and the presence of Eulepidina without Nummulites suggests a Late Oligocene age (Chattian; possibly into Aquitanian). Our claim is closely similar to Racey’s (1994) belief, but we strongly disagree with Bozorgnia (1966) and Adams and Bourgeois’ (1967) ideas regarding the last occurrence of Nummulites.
Concerning the Chattian, Aquitanian, and Burdigalian index LBFs, we would like to say that Borelis melo curdica/Borelis melo melo is an important Early Miocene species whose first occurrence marks the lower boundary of biozone-1 of Adams and Bourgeois (1967) (Heidari et al. 2012; Fig. 2). The first occurrence of B. melo curdica is used by many researchers to mark the lower boundary of the Burdigalian (Wynd 1965; Adams and Bourgeois 1967; Rahaghi 1980; Seyrafian et al. 1996; Laursen et al. 2009; Van Buchem et al. 2010; Heidari et al. 2012). In this case, there is full agreement between all researchers who worked on the Iranian Oligo-Miocene Biozonation (Wynd 1965; Adams and Bourgeois 1967; Laursen et al. 2009; Van Buchem et al. 2010). Besides, according to Adams et al. (1983), B. melo curdica indicates an age no older than Burdigalian, although Cahuzac and Poignant (1997) suggested that it may be no older than Langhian (Ehrenberg et al. 2007). However, the first occurrence of B. melo curdica is used by Cahuzac and Poignant (1997) to mark the lower boundary of the Burdigalian deposits of the European basin (see SB25 of Cahuzac and Poignant: Borelis melo group-Miogypsina).
As yet, the biostratigraphy of the Qom Formation relies mainly on biozonations of Wynd (1965) and Adams and Bourgeois (1967). For the first time, Mohammadi et al. (2013) dated the lower deposits of the Qom Formation based on BouDagher-Fadel (2008). Mohammadi et al. (2013) believed that future works on the Qom Formation must be based on this approach, and all of the in situ Nummulites bearing layers of the Qom Formation should be ascribed to Rupelian in age. Therefore, in this study methods of BouDagher-Fadel (2008) and Mohammadi et al. (2013) were used for dating the in situ Nummulites bearing layers of the Qom Formation. Here, we use B. melo curdica as index of Burdigalian age. Based on BouDagher-Fadel (2008) the age range of genus Miogypsina is from Latest Early Oligocene (Rupelian, P21, in America), Late Oligocene (Chattian, P22, in Mediterranean), Early Miocene (Aquitanian, N4, in the Far East) to Early Miocene (Burdigalian, in America) and Middle Miocene (Middle Serravallian, in the Indo-Pacific province). However, Miogypsina has been regarded as indicative of Early Miocene age (Adams et al. 1983) of Iran, but based on Ehrenberg et al. (2007), Laursen et al. (2009), and Van Buchem et al. (2010) its first appearance in Iran is in latest Chattian.
In Bujan section, on the basis of the distribution of foraminifera, two foraminiferal assemblages were identified. They are described in ascending order, following the stratigraphic development.
-
Assemblage 1
This assemblage extends from the base of the Qom Formation to height of 95 m. Nummulites spp. recorded from only a single layer of the Bujan section. Sample nos. B-59, B-59a, B-59b, and B-59c were collected from the mentioned layer. However, Nummulites spp. are so abundant that sometime rocks are totally formed by them (Fig. 5). Their sizes reach up to several millimeters as they are easily visible by the naked eye in the field (Fig. 5). Their abundance in thin section is so high that only remained spaces between grains are filled with carbonate mud so that their thin sections are characterized by packstone texture of grain-supported type (Fig. 5). It is worth mentioning that the mentioned Nummulites spp. bearing layer was recorded by Hosseinipour (2004) too. In Bujan section, the last occurrences of Nummulites spp. are in sample nos. B-59b and B-59c (thickness of 95 m from the base). Therefore, the first 95-m thickness of study section is confidently deposited in Rupelian and attributed to Rupelian because of the presence of Nummulites spp. (N. fichteli-intermedius, Nummulites sp; Figs. 4 and 5). However, we couldn’t say that these Nummulites spp. indicate the end of Rupelian because they were recorded from a single layer. Therefore, maybe deposition of the Rupelian deposits continued into the related layers of the assemblage 2.
The following faunal assemblages have been recognized in this interval: N. fichteli-intermedius, Nummulites sp. Eulepidina sp., Eulepidina dilatata, Eulepidina elephantina, Nephrolepidina sp., Neorotalia viennoti, Borelis pygmaea, Archaias kirkukensis, Dendritina rangi, Austrotrillina howchini, Austrotrillina sp., Quinqueloculina sp., Triloculina trigonula, Triloculina tricarinata, textularids, corallinacean algae (Lithotaminium, Lithophyllum), bryozoa, and echinoids. The presence of Nummulites spp. (N. fichteli, Nummulites vascus, Nummulites sp.) leads to the age of the lower part of the formation to be determined as Rupelian (Figs. 7 and 8).
-
Assemblage 2
This assemblage occurs from height 95 m to top of the section. Based on the absence of B. melo curdica (index of the lower boundary of the Burdigalian) and Miogypsina (index of Aquitanian and latest Chattian in Iran) deposition of the upper part of the Bujan section, in latest Chattian?, Aquitanian and Burdigalian ages is impossible. However, based on its position above Assemblage 1(Rupelian in age) and interpretations of Assemblage 1, it could be attributed to the Rupelian-Chattian. This assemblage is correlated with Lepidocyclina-Operculina-Ditrupa assemblage zone of Wynd (1965) and Laursen et al. (2009) (Fig. 2) which is attributed to Rupelian and Chattian. The following faunal assemblages have been recognized in this interval: Eulepidina sp., Eulepidina dilatata, E. elephantina, Lepidocyclina sp., Nephrolepidina sp., N. tournoueri, Neorotalia sp., N. viennoti, B. pygmaea, Sphaerogypsina sp., Borelis haueri, Archaias sp., Operculina complanata, Heterostegina sp., Amphistegina sp. D. rangi, A. howchini, Austrotrillina asmariensis, Quinqueloculina sp., T. trigonula, T. tricarinata, textularids, corallinacean algae (Lithotaminium, Lithophyllum), bryozoa, and echinoids.
In Varkan section, the last occurrence of Nummulites spp. in Varkan material is in the last samples which are therefore considered to mark the end of Rupelian. Thus, according to the presence of N. vascus and Nummulites sp., throughout Varkan section, the whole deposits of the section are attributed to the Rupelian. The following faunal assemblages have been recognized in this interval: N. vascus, Nummulites sp., N. fichteli, Eulepidina sp., Eulepidina elephantina, Lepidocyclina sp., Nephrolepidina sp., N. viennoti, Neorotalia sp., Nephrolepidina sp., O. complanata, Heterostegina sp., Amphistegina sp. B. pygmaea, Eulepidina sp., Sphaerogypsina globulus, Sphaerogypsina sp., B. haueri, Archaias sp., Peneroplis tomasi, D. rangi, A. howchini, Austrotrillina sp., Haplophragmium sp., Cibisides sp., Elphidium sp., Quinqueloculina sp., T. trigonula, T. tricarinata, textularids, Ditrupa sp., corallinacean algae (Lithotaminium, Lithophyllum, Subterraniphyllum thomasi), bryozoa (Tubucellaria), and echinoids (Figs. 6, 7, and 8).
In Khurabad (Qom) section, on the basis of the distribution of foraminifera, four foraminiferal assemblages were identified. They are described in ascending order, following the stratigraphic development.
-
Assemblage 1
This assemblage extends from the base of the Qom Formation to height of 60.5 m. The last occurrence of Nummulites spp. in Khurabad material is in sample no. K20 which is therefore considered to mark the end of Rupelian. Thus, based on the presence of N. vascus and Nummulites sp. the lower 60.5 m of the Qom section is attributed to the Rupelian. The following faunal assemblages have been recognized in this interval: N. vascus, Nummulites sp., Eulepidina sp., Lepidocyclina sp., Nephrolepidina sp., Nephrolepidina tournoueri, Neorotalia sp., Operculina sp., Amphistegina sp. textularids, Ditrupa sp., corallinacean algae (Lithotaminium, Lithophyllum), bryozoa, and echinoids (Figs. 7 and 8).
-
Assemblage 2
This assemblage appears from height 60.5 to 93.5 m. The absence of B. melo curdica (index of the lower boundary of the Burdigalian) and Miogypsina (index of Aquitanian and latest Chattian in Iran), its position above Assemblage 1(Rupelian in age) and below the first evaporate succession of the section (evaporate deposits of the Qom Formation are deposited totally in the Early Miocene (Aquitanian–Burdigalian), Mohammadi et al. 2013), and presence of Lepidocyclinids without Nummulites caused the deposition of this part to be defined as Chattian. The following faunal assemblages have been recognized in this interval: Eulepidina sp., Lepidocyclina sp., Nephrolepidina sp., N. tournoueri, Neorotalia sp., O. complanata, Amphistegina sp. Eulepidina sp., Austrotrillina sp., Elphidium sp., Quinqueloculina sp., Pyrgo sp., miliolids, textularids, corallinacean algae (Lithotaminium, Lithophyllum), bryozoa, and echinoids (Figs. 7 and 8).
-
Assemblage 3
This assemblage appears from height 93.5 to 209.5 m. The absence of B. melo curdica and Nummulites spp., the presence of Miogypsina and evaporate succession, its position between Assemblage 2 (Chattian in age) and Assemblage 4 (Burdigalian in age), and presence of Lepidocyclinids without Nummulites caused the deposition of the part including assemblage 3 to be attributed to Aquitanian. The following faunal assemblages have been recognized in this interval: Eulepidina sp., Lepidocyclina sp., Nephrolepidina sp., N. tournoueri, N. viennoti, Miogypsina sp., Neorotalia sp., O. complanata, Heterostegina sp., Amphistegina sp., B. pygmaea, Eulepidina sp., Sphaerogypsina sp., D. rangi, Austrotrillina sp., Cibisides sp., Elphidium sp., Pyrgo sp., miliolids, textularids, corallinacean algae (Lithotaminium, Lithophyllum), bryozoa (Tubucellaria), and echinoids (Figs. 7 and 8).
-
Assemblage 4
This assemblage occurs from height 209.5 m to the top of the section (Fig. 9). The first occurrence of B. melo curdica in our material is in sample no. K-72 which is therefore considered to mark the beginning of Burdigalian. Based on the presence of B. melo curdica, the last 50 m of the Qom section (From 209.5 to 260) is attributed to the Burdigalian. This assemblage is Burdigalian in age and is correlated with B. melo curdica zone of Wynd (1965), B. melo-Meandropsina iranica assemblage zone of Adams and Bourgeois (1967) and B. melo curdica-B. melo melo assemblage zone of Laursen et al. (2009) and Van Buchem et al. (2010) which are attributed to Burdigalian.
The following faunal assemblages have been recognized in this interval: B. melo curdica, Miogypsina sp., B. pygmaea, Borelis sp., Archaias sp., Peneroplis sp., D. rangi, A. howchini, Austrotrillina sp., Haplophragmium sp., Elphidium sp., Quinqueloculina sp., T. trigonula, T. tricarinata, Pyrgo sp., textularids, Ditrupa sp., corallinacean algae (Lithotaminium, Lithophyllum), bryozoa (Tubucellaria), and echinoids (Figs. 7 and 8).
In summary, on the basis of biostratigraphy data and foraminifera assemblages, the Qom Formation is Rupelian, Rupelian-Chattian, and Rupelian-Burdigalian in age in Varkan, Bujan, and Qom (Khurabad) areas, respectively. The obtained results are compatible with the common trend of the transgression of the Tethyan Seaway on the Iranian Plate.
Conclusions
Three stratigraphic sections of the Qom Formation in Sanandaj–Sirjan fore-arc and Central Iran back-arc basins were studied in order to determine their accurate age. LBF (mainly Nummulites, lepidocyclinids,miogypsinids, and Borelis) biostratigraphically are the main biota of the Qom Formation in the study areas and therefore used for age dating of the study sections. This study led to the following conclusions:
-
1.
The last true Nummulites spp. became extinct at the top of Late Rupelian and all of the in situ Nummulites bearing layers of the Qom Formation belong to Rupelian in age.
-
2.
The presence of in situ Nummulites (with or without Eulepidina) in Qom Formation successions indicates an Early Oligocene (Rupelian) age.
-
3.
Lepidocyclinids which originated in the Americas during the Middle Eocene and migrated eastward through the Tethyan/Mediterranean corridor reached Iran in the Rupelian.
-
4.
The first appearance of Lepidocyclina spp., in our study sections and in several others stratigraphic sections of the Qom and Asmari formations, before the last occurrence of Nummulites spp., clearly indicates that the earliest Lepidocyclinidae reached Iran in the Rupelian (Early Oligocene).
-
5.
The Lepidocyclinids are widespread in the Chattian marine deposits of Iran.
-
6.
The presence of Eulepidina without Nummulites suggests a Late Oligocene age (Chattian; possibly into Aquitanian).
-
7.
N. intermedius-N. vascus Assemblage zone of Wynd (1965) and Eulepidina-Nephrolepidina-Nummulites Assemblage zone of Adams and Bourgeois (1967), which are proposed for Oligocene (Rupelian-Chattian), should be ascribed to Rupelian in age.
-
8.
On the basis of biostratigraphic data and foraminifera assemblages, the Qom Formation is Rupelian, Rupelian-Chattian, and Rupelian-Burdigalian in age in Varkan, Bujan, and Qom (Khurabad) areas, respectively.
-
9.
The obtained results are compatible with the common trend of the transgression of the Tethyan Seaway on the Iranian Plate.
References
Abaie I, Ansari H, Badakhshan A, Jaafaki A (1963) History and development of the Alborz and Seraih fields, central Iran. In: Proc 6th World Petrol Conf, Frankfurt, pp. 1–111
Abaie I, Ansari H, Badakhshan A, Jaafari A (1964) History and development of the Alborz and Sarajeh fields of central Iran. Bull Iran Petrol Inst 15:561–574
Adams CG (1984) Neogene larger foraminifera, evolutionary and geological events in the context of datum planes. In: Ikebe N, Tsuchi R (eds) Pacific Neogene datum planes. University of Tokyo Press, Tokyo, pp 47–67
Adams T, Bourgeois F (1967) Asmari biostratigraphy. Iranian Oil Operating Companies, Geological and Exploration Division, Report 1074
Adams CG, Gentry AW, Whybrow PJ (1983) Dating the terminal Tethyan event. Utrecht Micropaleontol Bull 30:273–298
Alavi M (2004) Regional stratigraphy of the Zagros fold-thrust belt of Iran and its proforeland evolution. Am J Sci 304:1–20
Amirshahkarami M, Vaziri-Moghaddam H, Taheri A (2007a) Paleoenvironmental model and sequence stratigraphy of the Asmari Formation in southwest Iran. Hist Biol 19:173–183
Amirshahkarami M, Vaziri-Moghaddam H, Taheri A (2007b) Sedimentary facies and sequence stratigraphy of the Asmari Formation at chaman-Bolbol, Zagros Basin, Iran. J Asian Earth Sci 29:947–959
Amirshahkarami M, Ghabishavi A, Rahmani A (2010) Biostratigraphy and paleoenvironment of the larger benthic foraminifera in wells sections of the Asmari Formation from the Rag-e-Safid oil field, Zagros Basin, southwest Iran. Stratigr Sedimentol 40:63–84
Armstrong H, Brasier MD (2005) Microfossils. Blackwell, Oxford
Baker RD, Hallock P, Moses EF, Williams DE, Ramirez A (2009) Larger foraminifers of the Florida Reef Tract, USA: distribution patterns on reef-rubble habitats. J Foraminifer Res 39:267–277
Bassi D, Nebelsick JH (2010) Components, facies and ramps: redefining Upper Oligocene shallow water carbonates using coralline red algae and larger foraminifera (Venetian area, northeast Italy). Palaeogeogr Palaeocl 295:258–280
Beavington-Penney SJ, Racey A (2004) Ecology of extant nummulitids and other larger benthic foraminifera: applications in palaeoenvironmental analysis. Earth Sci Rev 67:219–265
Behforouzi E, Safari A (2011) Biostratigraphy and paleoecology of the Qom Formation in the Chenar area (northwestern Kashan), Iran. Rev Mex Cienc Geol 28:555–565
Berberian M, King G (1981) Towards a paleogeography and tectonic evolution of Iran. Can J Earth Sci 18:210–265
Berning B, Reuter M, Piller W, Harzhauser M, Kroh A (2009) Larger foraminifera as a substratum for encrusting bryozoans (Late Oligocene, Tethyan Seaway, Iran). Facies 55:227–241
BouDagher-Fadel MK (2008) Evolution and geological significance of larger benthic foraminifera. Developments in palaeontology and stratigraphy, 21. Elsevier, Amsterdam
Boudagher-Fadel MK, Lokier S (2005) Significant Miocene larger foraminifera from south central Java. Rev Micropaleontol 24:291–309
Boudagher-Fadel MK, Lord AR (2000) The evolution of Lepidocyclina (L.) Isolepidinoides, L. (Nephrolepidina) Nephrolepidinoides sp. nov., L. (N.) brouweri and L. (N.) ferreroi in the Late Oligocene–Miocene of the Far East. J Foram Res 30:71–76
BouDagher-Fadel MK, Price GD (2010) Evolution and paleogeographic distribution of the lepidocyclinids. J Foram Res 40:79–108
Boukhary M, Scheibner C (2009) On the origin of nummulites: Urnummulites schaubi n. gen. n. sp., from the Late Paleocene of Egypt. Micropaleontology 55:413
Bozorgnia F (1965) Qum Formation stratigraphy of the Central Basin of Iran and its intercontinental position. ECAFE. In: Symp Dev Petr Res Asia and Far East, third session, Tokyo, 1–9
Bozorgnia F (1966) Qum formation stratigraphy of the Central Basin of Iran and its intercontinental position. Bull Iran Petrol Inst 24:69–76
Cahuzac B, Poignant A (1997) An attempt of biozonation of the Oligo-Miocene in the European basins by means of larger neritic foraminifera. Bull Soc Geol Fr 168:155–170
Daneshian J, Ramezani Dana L (2007) Early Miocene benthic foraminifera and biostratigraphy of the Qom Formation, Deh Namak, central Iran. J Asian Earth Sci 29:844–858
Dozy J (1945) A geological reconnaissance of the area of Veramin and the Siah Kuh (Central Persia). Geological Report 6 (Tehran file)
Dozy J (1955) A sketch of post-Cretaceous volcanism in central Iran. Leidse Geol Meded 20:48–57
Ehrenberg S, Pickard N, Laursen G, Monibi S, Mossadegh Z, Svånå T, Aqrawi A, McArthur J, Thirlwall M (2007) Strontium isotope stratigraphy of the Asmari Formation (Oligocene–Lower Miocene), Sw Iran. J Pet Geol 30:107–128
Furon R (1941) Geologie du plateau Iranien:(Perse-Afghanistan-Beloutchistan). Museum National d’histoire naturelle
Furon R, Marie P (1939) Sur la microfaune des marnes aquitaniennes Lepidocyclines de Qoum (Perse). Compte Rendu Sommaire des Séances de la Société Géologique de France: 79–80
Furrer M, Soder P (1955) The Oligo-Miocene marine formation in the Qum region (Central Iran). In: Proceedings of the 4th World Petroleum Congress, Roma, Italia, Section I/A/5, pp. 267–277
Gansser A (1955) New aspects of the geology in Central Iran. In: Proceedings of the 4th World Petroleum Congress, Roma, Italia, section I/A/5. Reprint, 280–300
Ghasemi A, Talbot C (2006) A new tectonic scenario for the Sanandaj–Sirjan zone (Iran). J Asian Earth Sci 26:683–693
Gilg HA, Boni M, Balassone G, Allen CR, Banks D, Moore F (2006) Marble-hosted sulfide ores in the angouran Zn-(Pb–Ag) deposit, NW Iran: interaction of sedimentary brines with a metamorphic core complex. Miner Deposita 41:1–16
Gradstein FM, Ogg JG, Smith AG (2005) A geologic time scale 2004. Cambridge University Press, Cambridge
Hadavi F, Notghi Moghaddam M, Mousazadeh H (2010) Burdigalian–serravalian calcareous nannoplanktons from Qom Formation, north-center Iran. Arab J Geosci 3:133–139
Heidari A, Mahboubi A, Moussavi-Harami R, Gonzalez L, Moalemi SA (2012) Biostratigraphy, sequence stratigraphy, and paleoecology of the Lower-Middle Miocene of Northern Bandar Abbas, Southeast Zagros basin in south of Iran. Arab J Geosci doi:10.1007/s12517-012-0803-3
Hajian J, Emammi MH (1991) Geological quadrangle map of Qom. Geological Survey of Iran, Tehran
Hallock P (1985) Why are larger foraminifera large? Paleobiology 11:195–208
Harzhauser M (2000) Paleobiogeography and palecology of Oligocene and Lower Miocene gastropods in the eastern Mediterranean and the western Indo-Pacific. PhD Dissertation, University of Vienna
Harzhauser M (2004) Oligocene gastropod faunas of the eastern Mediterranean (mesohellenic trough/Greece and Esfahan-Sirjan Basin/central Iran). Cour Forsch Inst Senckenberg 248:93–181
Hasani MJ, Vaziri MR (2011) Early-Miocene gastropods from Khavich area, south of Sirjan, (Kerman, Iran): biostratigraphy, paleogeography and paleoecology. J Sci Islam Repub Iran 22:125–133
Haynes JR, Racey A, Whittaker JE (2010) A revision of the Early Palaeogene nummulitids (Foraminifera) from northern Oman, with implications for their classification. In: Whittaker JE, Hart MB (ed) Micropalaeontology, sedimentary environments and stratigraphy: a tribute to Dennis Curry (1912–2001). The Micropalaeontological Society, Special Publications, Geological Society, pp. 29–89
Heydari E, Hassanzadeh J, Wade W, Ghazi A (2003) Permian–Triassic boundary interval in the Abadeh section of Iran with implications for mass extinction: part 1–sedimentology. Palaeogeogr Palaeoclimatol Palaeoecol 193:405–423
Hohenegger J (2000) Coenoclines of larger foraminifera. Micropaleontology 46:127–151
Hosseinipour F (2004) Study of biostratigraphy and paleoecology of Oligo-Miocene sediments at north and northwest Sirjan. MSc. Dissertation, Shahid Bahonar University of Kerman
Iaccarino S, Premoli Silva I (2005) Practical manual of Oligocene to Middle Miocene planktonic foraminifera. In: Biolzi M, Iaccarino S, Rettori R (eds) International school on planktonic foraminifera 4 course. University of Perugia Press, Perugia, pp 14–18
Jones R, Racey A (1994) Cenozoic stratigraphy of the Arabian Peninsula and Gulf. In: Simmons MD (ed) Micropaleontology and hyrocarbon exploration in the Middle East. Chapman and Hall, London, pp 273–307
Kalanat B, Vaziri-Moghaddam H, Taheri A (2010) Biostratigraphy and paleoecology of the Asmari Formation, SW Firozabad. Sediment Facies 3:71–84
Khaksar K, Maghfouri–Moghadam I (2007) Paleontological study of the echinoderms in the Qom Formation (Central Iran). Earth Sci Res J 11:57–79
Khalili M, Beavers R, Torabi H (2007) Depositional environment of the evaporitic unit (D-member) of the Qom Formation (Central Iran). Carbonate Evaporite 22:101–112
Koukousioura O, Dimiza MD, Triantaphyllou MV, Hallock P (2011) Living benthic foraminifera as an environmental proxy in coastal ecosystems: a case study from the Aegean Sea (Greece, NE Mediterranean). J Mar Syst 88:489–501
Kuhn O (1933) Das becken von Isfahan-Saidabad und seine altmiocane korallenfauna, palaeontographica. Abt A 79:173–218
Kuss J, Boukhary MA (2008) A new upper Oligocene marine record from northern Sinai (Egypt) and its paleogeographic context. Geoarabia-Manama 13:59–84
Laursen G, Monibi S, Allan T, Pickard N, Hosseiney A, Vincent B, Hamon Y, Van–Buchem F, Moallemi A, Druillion G (2009) The Asmari Formation revisited: changed stratigraphic allocation and new biozonation: In: First International Petroleum Conference & Exhibition, Shiraz, Iran
Loeblich AR, Tappan H (1988) Foraminiferal genera and their classification. Van Nostrand Reinhold, New York, two volumes
Loftus WK (1855) On the geology of portions of the Turko-Persian frontier, and of the districts adjoining. Q J Geol Soc 11:247–344
Mandic O (2000) Oligocene to Early Miocene pectinid bivalves of the Western Tethys (N-Greece, S-Turkey, Central Iran and NE-Egypt)-taxonomy and paleobiogeography. PhD Dissertation, University Vienna
Martínez-Colón M, Hallock P, Green-Ruíz C (2009) Strategies for using shallow-water benthic foraminifers as bioindicators of potentially toxic elements: a review. J Foram Res 39:278–299
Mohammadi E, Safari A, Vaziri-Moghaddam H, Vaziri MR, Ghaedi M (2011) Microfacies analysis and paleoenviornmental interpretation of the Qom Formation, south of the Kashan, central Iran. Carbonate Evaporite 26:255–271
Mohammadi E, Hasanzadeh-Dastgerdi M, Ghaedi M, Dehghan R, Safari A, Vaziri-Moghaddam H, Baizidi C, Vaziri M, Sfidari E (2013) The Tethyan Seaway Iranian Plate Oligo-Miocene deposits (the Qom Formation): distribution of Rupelian (Early Oligocene) and evaporate deposits as evidences for timing and trending of opening and closure of the Tethyan Seaway. Carbonate Evaporite 28:321–345
Nebelsick JH, Rasser MW, Bassi D (2005) Facies dynamics in Eocene to Oligocene circumalpine carbonates. Facies 51:197–217
NIOC (1969) Geological Maps of Iran Sheet No. 5 (South Central Iran). National Iranian Oil company
NIOC (1977) Geological Maps of Iran Sheet No. 2 (North Central Iran). National Iranian Oil company
NIOC (1978) Geological Maps of Iran Sheet No. 1 (North West Iran). National Iranian Oil company
Okhravi R (1998) Synsedimentary cementation in the Lower Miocene reefal carbonates of the central basin, Iran. Carbonate Evaporite 13:136–144
Okhravi R, Amini A (1998) An example of mixed carbonate–pyroclastic sedimentation (Miocene, central basin, Iran). Sediment Geol 118:37–54
Özcan E, Less G, Báldi-Beke M, Kollányi K, Acar F (2009) Oligo-Miocene foraminiferal record (Miogypsinidae, Lepidocyclinidae and Nummulitidae) from the Western Taurides (SW Turkey): biometry and implications for the regional geology. J Asian Earth Sci 34:740–760
Racey A (1994) Biostratigraphy and palaeobiogeographic significance of Tertiary nummulitids (foraminifera) from northern Oman. In: Simmons MD (ed) Micropaleontology and hyrocarbon exploration in the Middle East. Chapman and Hall, London, pp 343–370
Racey A (1995) Lithostratigraphy and larger foraminiferal (nummulitid) biostratigraphy of the Tertiary of northern Oman. Micropaleontology 41:1–123
Rahaghi A (1973) Étude de quelques grands foraminifères de la formation de Qum (Iran central). Rev Micropal 16:23–38
Rahaghi A (1980) Tertiary faunal assemblage of Qum-Kashan, Sabzewar and Jahrum areas. NIOC 8(8):1–64
Rahimpour-Bonab H, Kalantarzadeh Z (2005) Origin of secondary potash deposits; a case from Miocene evaporites of NW central Iran. J Asian Earth Sci 25:157–166
Rahimzadeh F (1994) Geology of Iran: Oligocene–Miocene, Pliocene. Geological Survey of Iran, Tehran
Rahmani A, Vaziri-Moghaddam H, Taheri A, Ghabeishavi A (2009) A model for the paleoenvironmental distribution of larger foraminifera of Oligocene–Miocene carbonate rocks at Khaviz Anticline, Zagros Basin, SW Iran. Hist Biol 21:215–227
Reuter M, Piller WE, Harzhauser M (2009a) Comment on? Revision of the age of the Qom Formation in the Central Iran Basin, Iran? by Zhu et al. (Journal of Asian Earth Sciences, 2007, vol. 29, 715? 721). J Asian Earth Sci 34:699–701
Reuter M, Piller W, Harzhauser M, Mandic O, Berning B, Rögl F, Kroh A, Aubry MP, Wielandt-Schuster U, Hamedani A (2009b) The Oligo-/Miocene Qom Formation (Iran): evidence for an Early Burdigalian restriction of the Tethyan Seaway and closure of its Iranian gateways. Int J Earth Sci 98:627–650
Riben H (1935) Contribution a la ge’ologie de l’Azerbeidjan Persan. Bulletin de la Socie‘te’ Neuchateloise des Sciences Naturelles. Neuchatel 59:20–144
Sadeghi R, Vaziri-Moghaddam H, Taheri A (2009) Biostratigraphy and paleoecology of the Oligo-Miocene succession in Fars and Khuzestan areas (Zagros Basin, SW Iran). Hist Biol 20:1–15
Schuster F, Wielandt U (1999) Oligocene and Early Miocene coral faunas from Iran: palaeoecology and palaeobiogeography. Int J Earth Sci 88:571–581
Seddighi M, Vaziri-Moghaddam H, Taheri A, Ghabeishavi A (2011) Depositional environment and constraining factors on the facies architecture of the Qom Formation, Central Basin, Iran. Hist Biol 24:91–100
Seyrafian A, Torabi H (2005) Petrofacies and sequence stratigraphy of the Qom Formation (Late Oligocene-Early Miocene?), north of Nain, southern trend of central Iranian Basin. Carbonate Evaporite 20:82–90
Seyrafian A, Vaziri-Moghaddam H, Torabi H (1996) Biostratigraphy of the Asmari Formation, Burujen area, Iran. J Sci Islam Repub Iran 7:31–48
Shafiei SH, Alavi SA, Mohajjel M (2011) Calcite twinning constraints on paleostress patterns and tectonic evolution of the Zagros hinterland: the Sargaz complex, Sanandaj–Sirjan zone, SE Iran. Arab J Geosci 4:1189–1205
Sharaf EF, BouDagher-Fadel MK, Simo J, Carroll A (2005) Biostratigraphy and strontium isotope dating of Oligocene–Miocene strata, East Java, Indonesia. Stratigraphy 2:1–19
Soheili M (1995) Geological quadrangle map of Sirjan. Geological Survey of Iran, Tehran
Sooltanian N, Seyrafian A, Vaziri-Moghaddam H (2011) Biostratigraphy and paleo-ecological implications in microfacies of the Asmari Formation (Oligocene), Naura anticline (Interior Fars of the Zagros Basin), Iran. Carbonate Evaporite 26:167–180
Stahl A (1911) Persien. In: Handbuch der Regionalen Geologie Heidelberg (Winter) Hft 8 (5):6
Stainforth RM, Lamb J, Luterbacher H, Beard J, Jeffords R (1975) Cenozoic planktonic foraminiferal zonation and characteristics of index forms
Tietze E (1875) Ein ausflug nach dem siahkuh (schwarzer berg) in persien. Mitteilungen der Geographischen Geographischen Gesellschaft Wien 18(8):257–267
Van Buchem F, Allan T, Laursen G, Lotfpour M, Moallemi A, Monibi S, Motiei H, Pickard N, Tahmasbi A, Vedrenne V (2010) Regional stratigraphic architecture and reservoir types of the Oligo-Miocene deposits in the dezful Embayment (Asmari and Pabdeh Formations) SW Iran. Geol Soc Lond Spec Publ 329:219–263
Vaziri-Moghaddam H, Torabi H (2004) Biofacies and sequance strayigraphy of the Oligocene succession, central basin, Iran. N Jb Geol Palaont Stuttgart 6:321–344
Vaziri-Moghaddam H, Kimiagari M, Taheri A (2006) Depositional environment and sequence stratigraphy of the Oligo–Miocene Asmari Formation is SW Iran. Facies 52:41–51
Vaziri-Moghaddam H, Seyrafian A, Taheri A, Motiei H (2010) Oligocene-Miocene ramp system (Asmari Formation) in the NW of the Zagros basin, Iran: microfacies, paleoenvironment and depositional sequence. Rev Mex Cienc Geol 27:56–71
Vaziri-Moghaddam H, Kalanat B, Taheri A (2011) Sequence stratigraphy and depositional environment of the Oligocene deposits at Firozabad section, southwest of Iran based on microfacies analysis. Geophys J R Astron Soc 1:71–152
Wynd J (1965) Biofacies of the Iranian consortium agreement area: Iranian Oil Offshore Company Report 1082
Yazdi M, Shirazi MP, Rahiminejad AH, Motavalipoor R (2012) Paleobathymetry and paleoecology of colonial corals from the Oligocene–Early Miocene (?) Qom Formation (Dizlu area, central Iran). Carbonate Evaporite 27:395–405
Yazdi-Moghadam M (2011) Early Oligocene larger foraminiferal biostratigraphy of the Qom Formation, south of Uromieh (NW Iran). Turk J Earth Sci 20:847–856
Zahedi M, Amidi SM (1991) Geological quadrangle map of Kashan. Geological Survey of Iran, Tehran
Zhu Y, Qi Y, Zhang B, Yang H, He C, Wang S, Zhou W, Zhu Q, Li Z (2007) Revision of the age of the Qom Formation in the central Iran basin, Iran. J Asian Earth Sci 29:715–721
Acknowledgments
This paper has benefited from the discussions with many colleagues. The authors are indebted to Prof. Guillem Mateu-Vicens and another anonymous reviewer; their constructive comments, criticisms and suggestions significantly improved the quality of this paper. Comments and suggestions made by Associate Editor Prof. Biswajeet Pradhan and by Chief Editor Abdullah M. Al-Amri, which significantly improved this manuscript, are much appreciated. Assistance in the field and useful discussion and suggestions on a first draft of the manuscript by Dr. Mehdi Ghaedi are very much appreciated. Prof. Andrew Racey is gratefully acknowledged for providing very useful discussions and articles about Nummulites. Prof. Marcelle K. BouDagher-Fadel, Prof. Pamela Hallock, Prof. Marco Brandano, Prof. Guillem Mateu-Vicens, Prof. Luis Pomar, Prof. Willem Renema, Prof. Alan Lord, Prof. Mathias Harzhauser, and Dr. Ricardo Meireles are gratefully appreciated for providing very useful articles and suggestions. Dr. Mohammad Allahkarampour Dill, Dr. Shabafrooz, Mr. Ali Opera, Dr. Ali Rahmani, Dr. Ali Ghabeishavi, and National Iranian Oil Company are much appreciated for their inestimable assistance in taking thin section photomicrographs and Laboratory supports. Mr. Mohammad Shrifi and Mr. Saeid Latifian are thanked for their assistance in the field. Mrs. Chavare Baizidi is gratefully acknowledged for many hours of assistance for disaggregation softer samples and Figure preparation.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mohammadi, E., Vaziri, MR. & Dastanpour, M. Biostratigraphy of the nummulitids and lepidocyclinids bearing Qom Formation based on larger benthic foraminifera (Sanandaj–Sirjan fore-arc basin and Central Iran back-arc basin, Iran). Arab J Geosci 8, 403–423 (2015). https://doi.org/10.1007/s12517-013-1136-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12517-013-1136-6