
Abstract
Sediments of Maloe Olen’e Lake in the central part of the Kaliningrad oblast were subjected to analysis of the botanical composition of peat, pollen analysis, and radiocarbon dating. Four local pollen assemblage zones as reflecting the dominant plant communities of the surrounding territories during the Holocene were assigned. Analysis of plant macrofossils resulted in the identification of the stages of floating mat vegetation development in the Late Holocene. The atypical sequence of peat deposition, with transition mire peat located above raised bog peat, was identified. Sediment accumulation rate was calculated according to radiocarbon dating.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
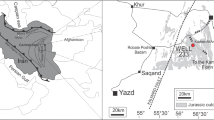
Mire ecosystems of lacustrine origin are quite scarce and poorly characterized elements of the Kaliningrad oblast landscape, even though analysis of the distinctive features of their formation can contribute to our understanding of the development of the environmental conditions in the South-Eastern Baltic area in the postglacial time. The authors have analyzed pollen assemblages and plant macrofossils in the organogenic Holocene deposits from the ecosystem of Maloe Olen’e Lake located in the central part of the Kaliningrad oblast and currently undergoing a transition from lake to mire due to terrestrialization (Figs. 1a–1c).

Schematic map of the study area: (a) geographic location of the Kaliningrad oblast, (b) scheme of the landscape areas in Kaliningrad oblast (Geografiya …, 2008) and location of Maloe Olen’e Lake, (c) surface structure of Maloe Olen’e Lake. Designations: (1) hilly terminal-moraine landscapes, (2) moraine plains, (3) glaciolacustrine plains, (4) ancient alluvial and glaciofluvial plains, (5) recent alluvial plains, (6) deltaic lowlands, (7) aeolian landscapes, (8) terrestrialized part of the lake (floating mat), (9) open water surface, (10) coring and sampling site.
METHODS
Coring and bottom sediment sampling for botanical, pollen, and radiocarbon analyses were performed on the floating mat of the lake-mire system (Fig. 1c; 54.56646° N; 21.70243° E). Botanical composition of the peat was analyzed according to the conventional approaches (Korotkina, 1939). Sample processing for pollen and spore preparation was performed as described by Faegri and Iversen (1989). The С2 software (Juggins, 2014) was used to construct the pollen diagram and the diagram of the botanical composition of the peat. The absolute age of the sediments was determined in the Radiocarbon Dating Laboratory of the Geography Institute, Russian Academy of Sciences.
Stratigraphy and botanical composition of the peat. Maloe Olen’e Lake is located in a 10.5-m deep closed kettle; the kettle bottom is covered by glaciolacustrine clays, with a thick gyttja layer located above the clay layer (Fig. 2). The peat bed of the terrestrialized part of the lake is represented by a thick (3.5–4 m) and broad floating mat surrounding the lake. A water layer 3–5 m deep is located between the peat bed and the gyttja (Fig. 2). The floating mat is replaced by open water surface near the middle of the lake. Fen peat is completely absent from the floating mat. The entire peat bed has distinct raised bog peat and transition mire peat layer. Raised bog peat constitutes the lower two-thirds of the floating mat (120–350 cm), and the thin transition layer is located above it (Fig. 2), in contrast to the conventional sequence of the different types of peat.

Stratigraphic composition scheme for Maloe Olen’e Lake: (1) transition mire peat, (2) raised bog peat, (3) gyttja, (4) glaciolacustrine clay, (5) water, (6) borehole path.
Two stages of vegetation development (Fig. 3): Sphagnum fuscum (stage I) and Eriophorum polystachyon – Sphagnum angustifolium + S. magellanicum (stage II) were identified in the diagram of botanical composition of peat according to the results of analysis of plant macrofossils complexes in floating mat peats.

Botanical composition diagrams for the peats from Maloe Olen’e Lake. Vegetation stages: (I) reed stretches: raised Sphagnum fuscum bog at an active growth stage; (II) transition mire communities: Eriophorum polystachyon–Sphagnum angustifolium + S. magellanicum.
Pollen assemblages. Four local pollen assemblage zones characteristic of the dominant plant communities of the surrounding territories in the Holocene are distinguished in the pollen diagram of Maloe Olen’e Lake (Fig. 4).

Pollen diagram of Maloe Olen’e Lake: (1) transition mire peat, (2) Sphagnum raised-bog peat, (3) gyttja, (4) water, (5) glacio-lacustrine clay.
Local pollen assemblage zone 1 Betula–Pinus (1175–920 cm) is characterized by low species diversity with Betula (55–67%) and Pinus (27–40%) pollen predominating (Figs. 4; 5е, 5l, 5m) and insignificant amounts of pollen from Ulmus, Alnus, Picea, Corylus, Salix (Figs. 5b–5e, 5p, 5q), and Artemisia and Poaceae (Fig. 5n). Pediastrum boryanum remnants constitute a substantial part of the gyttja deposits (Figs. 6a, 6b). We believe that the pollen assemblage zone corresponds to the early Holocene (the Preboreal and first half of the Boreal). The development of microphyte communities and accumulation of thick gyttja layers at the average rate of 1.5 mm/year occurred in the lake at that time. The replacement of cryophilic vegetation by pine and birch communities took place in the neighboring areas.

Pollen grains and spores from the Holocene sediments of Maloe Olen’e Lake (magnification ×600): (a) Quercus Linnaeus, 1753; (b) Ulmus Linnaeus, 1753; (c) Corylus Linnaeus, 1753; (d) Alnus Miller, 1754 (with five pores); (e) Alnus Miller, 1754 (with four pores); (f) Betula Linnaeus sect. Albae Regel, 1865; (g) Carpinus Linnaeus, 1753; (h, i) Tilia Linnaeus, 1753; (j, k) Ericaceae Jussieu, 1789 (microspore tetrads); (l) Pinus Linnaeus, 1753 (view from below); (m) Pinus Linnaeus, 1753 (side view); (n) Poaceae Barnhart, 1895; (o) Polypodiaceae Jan Presl et Carl Presl, 1822 (spore); (p, q) Picea Albert Dietrich, 1824.

Plant macrofossils from the Holocene deposits of Maloe Olen’e Lake (magnification: c–d, f–g, i–j, l ×150; a–b, e, h, k ×600): (a, b) Pediastrum boryanum (Turpin) Meneghini, 1840; (c) Sphagnum fuscum (Schimper) Klinggraeff, 1872 (branch leaf); (d, e) Betula Linnaeus, 1753 (bark cell layer); (f) Sphagnum fuscum (Schimper) Klinggraeff, 1872 (stem leaf); (g, h) Pinus Linnaeus, 1753 (bark cell layer); (i) Sphagnum magellanicum Bridel, 1798 (branch leaf); (j, k) Eriophorum polystachyon Linnaeus, 1753 (leaf epidermis with fibers); (l) Eriophorum polystachyon Linnaeus, 1753 (leaf sheath epidermis).
Local pollen assemblage zone 2 Corylus–Ulmus (920–800 cm). The lower boundary of the palynozone corresponds to an increase of pollen percentage of thermophilic species (Corylus pollen value increases to 23%, and Ulmus pollen value, to 4.6%) accompanied by a decrease of Pinus pollen value (Figs. 4; 5b, 5c, 5l–5m). The character of the pollen spectrum corresponds to the second half of the Boreal and beginning of the Atlantic, this being in good agreement with the data for the neighboring areas (Neustadt, 1957; Lang, 1994; Palaeoecological …, 1996) and the results of the radiocarbon dating. Pine and birch forests predominated in the study area during that period, but thermophilic species already played an important role in the communities.
The absence of sediment material (Figs. 2, 4) due to interruption of sedimentation prevented us from drawing conclusions on the development of vegetation in the study area during the Middle Holocene and the first half of Late Holocene.
Local pollen assemblage zone 3 Carpinus (350–80 cm). Arboreal plant pollen predominates (97–99%); these plants are represented by Betula, Alnus, Pinus, Carpinus, Picea, Quercus, Tilia, Corylus, and Alnus (Figs. 4; 5a–5i, 5l, 5m, 5p, 5q), whereas Ulmus and Fagus pollen is present in small amounts. Radiocarbon dating indicated that the palynozone corresponded to the vegetation of the second half of the Sub-Atlantic period represented by broadleaf and spruce–broadleaf forests similar to the contemporary zonal communities of the South-Eastern Baltic area (Rastitel’nost’…, 1980). The two subzones (Fig. 4), Alnus (340–210 cm) and Picea (210–80 cm), identified within the pollen assemblage zone might reflect the centennial-scale climate variability.
Raised bog peat in the lower part of the floating mat is largely composed of Sphagnum fuscum (Figs. 3; 6c–6f), this being indicative of intensive expansion of raised bog communities during this period.
Local pollen assemblage zone 4 Pinus—NAP (the pollen of non-arboreal species) (80–5 cm). Tree pollen predominates, but herbaceous plant pollen makes a substantial contribution (up to 7.5%) (Figs. 4; 5j, 5k, 5n), with Poaceae, Artemisia, Chenopodiaceae, Compositae (Cichorioideae), and Ericaceae being the predominant sources of herbaceous plant pollen. The share of Pinus pollen among the arboreal species increases significantly (to 74.3%). The value of Betula, Carpinus, Picea, Quercus, Alnus, and Corylus pollen decreases significantly, and Fagus and Tilia pollen is nearly absent.
The hosting peat layer also changes: rather abrupt replacement of raised bog peat by transition mire peat with predominating remnants of Eriophorum polystachyon (Figs. 3; 6j–6l) and more mesotrophic Sphagnum species (Sphagnum angustifolium, S. capillifolium, and S. magellanicum), along with pine and birch bark (Figs. 3; 6d, 6e, 6g–6i) occurs at the depth of 120 cm and above. These deposits are of a subrecent age: they were supposedly formed within the 100–120 most recent years. Thus, pollen spectra of the pollen assemblage 4 provides information on the character of pollen deposition in the contemporary epoch and the degree of correspondence between this process and the composition of contemporary vegetation.
Radiocarbon dating showed that peat accumulation rate in the floating mat of Maloe Olen’e Lake was 1.6–1.7 mm/year throughout the second half of the Sub-Atlantic period. The high rate of peat increment in the recent decades (up to 11 mm/year) is due to the incomplete decomposition and ongoing decay of the plant remnants in the upper horizons.
REFERENCES
Geografiya Yantarnogo kraya Rossii (Geography of the Amber Land of Russia), Orlenok, V.V., Ed., Kaliningrad: Yantarnyi skaz, 2008.
Korotkina, M.Ya., Botanical analysis of peat, in Metody issledovaniya torfyanykh bolot (Methods of Peat Bog Investigation), Neustadt, M.I., Ed., Moscow: Narodn. Komissariat Zemledeliya RSFSR, 1939, pt. 2, pp. 5–59.
Neustadt, M.I., Istoriya lesov i paleogeografiya SSSR v golotsene (Forest History and Paleogeography of the USSR during the Holocene). Moscow: Akad. Nauk SSSR, 1957. Rastitel’nost’ Evropeiskoi chasti SSSR (Vegetation of the European Part of the USSR), Gribova, S.S., Isachenko, T.I., and Lavrenko, E.M., Eds., Leningrad: Nauka, 1980. Faegri, K. and Iversen, J., Textbook of Pollen Analysis, Caldwell, N.J.: The Blackburn Press, 1989.
Juggins, S., C2 Version 1.7.6. [Electronic resource]. 2014. Mode of access: https://www.staff.ncl.ac.uk/stephen.juggins/software/C2Home.htm.
Lang, G., Quartäre Vegetationsgeschichte Europas, Jena: G. Fischer, 1994.
Palaeoecological Events during the Last 15000 Years: Regional Syntheses of Palaeoecological Studies of Lakes and Mires in Europe, Berglund, B.E., Birks, H.J.B., Ralska-Jasiewiczowa, M., and Wright, H.E., Eds., Chichester, West Sussex, England; New York: John Wiley& Sons Ltd., 1996.
Funding
Fieldwork and pollen analysis were performed under the state assignment of the Shirshov Institute of Oceanology RAS (research theme no. 0149-2019-0013), analysis of the botanical composition of peat was funded from the Russian Academic Excellence Project at the Immanuel Kant Baltic Federal University, and data interpretation and review were supported by Russian Science Foundation (project no. 18-77-10016).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Translated by S. Semenova
Rights and permissions
About this article

Cite this article
Napreenko-Dorokhova, T.V., Napreenko, M.G. Pollen Assemblages and Plant Macrofossils in Holocene Deposits of Maloe Olen’e Lake (Kaliningrad Oblast). Paleontol. J. 53, 780–786 (2019). https://doi.org/10.1134/S0031030119080100
Received:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0031030119080100