
Abstract
We performed a palynological study on Jurassic successions of the Hojedk Formation, Central Iran. The sampled material was recovered from well no. 233, South Kuchak-Ali area belonging to the Tabas Block. The lithology comprises 475.80 m of green–grey shales and sandstones. Palynological assemblages were recovered including spores, pollen grains, dinoflagellate cysts, woody debris and amorphous organic matter (AOM). Furthermore, 22 taxa of plant macrofossils were identified. We analysed 48 samples which resulted in identification of 13 spore species (belonging to 11 genera) and three pollen species (from three genera) with fair preservation quality. Trilete spores dominate while pollen grains only represent 11.5% of the total particulate organic matter. Representatives of the fern spore Klukisporites are particularly abundant and the K. variegatus acme zone, which is of biostratigraphical importance, is identified in the host strata. The abundance of K. variegatus and K. scaberis suggests a Middle Jurassic (Bajocian–Bathonian?) age for the host strata. This age designation corroborates previous stratigraphic interpretations, mainly based on plant macrofossils for this formation. The palynomorphs indicate deposition in an oxygenated environment close to the shoreline in a deltaic system. The presence of subordinate marine palynomorphs (proximate dinoflagellate cysts) is probably indicative of intermittent sea level transgression. The affinity of the palynological assemblages from the Hojedk Formation indicates a diverse parental flora of, in descending quantitative order, ferns, bryophytes and gymnosperms, the latter mainly represented by conifers and ginkgophytes. Comparisons with modern plant ecology indicate accumulation of palynomorphs under a moist warm climate during the Middle Jurassic (Bajocian–Bathonian?) time. The comparison of parental plants with previous palaeofloristic studies indicates that the Tabas Block was located in the Mid-Asian part of the Indo-European floristic province of the Northern Hemisphere.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
From a geological point of view, Iran has been subdivided into a number of distinctive tectonostratigraphic units (Stocklin 1968). The Central Iran Basin is located in the central part of the country with narrow extensions to the northeast and northwest resulting in a triangular outline. In the east, it is limited by Dasht-e-Lut; in the south, southwest and west by the Sanandaj-Sirjan Metamorphic Belt; and in the north by the Alborz Mountains (Fig. 1). The Tabas Block is sandwiched between the Yazd Block in the west and the Lut Block in the east, as a part of the so-called Central-East Iranian Microcontinent (CEIM; Takin 1972). The thick Upper Triassic–Jurassic succession within this area has been subdivided into several formations (Stocklin and Nabavi 1971) which were subsequently combined into two lithostratigraphic groups: the Upper Triassic–lower Middle Jurassic Shemshak Group and the Middle–Upper Jurassic Magu Group (northern Tabas Block) or Bidou Group (southern Tabas Block), respectively (Aghanabati 1977, 1998). The wide distribution of the Shemshak Group across the Iran Plate represents deposition of sediments in a foreland basin, which was part of the Cimmerian Orogeny, during the Upper Triassic–Middle Jurassic (Fürsich et al. 2003; Seyed-Emami et al. 2004; Wilmsen et al. 2009b). In the area of the Tabas block, the Shemshak group consists of the marine Nayband Formation (Norian–Rhaetian), the non-marine Ab-e-Haji Formation, which consists of coal-bearing sediments deposited around the Triassic–Jurassic boundary, the marine, Toarcian–Aalenian Badamu Formation, followed by the Bajocian (−early Bathonian), coal-bearing Hojedk Formation. This formation is unconformably overlain by the Upper Bajocian–Upper Jurassic Magu (or Bidou) Group (Wilmsen et al. 2009a). The Hojedk Formation comprises arcosic-quartzitic sandstones and green to grey shales with coal-bearing intervals. The presence of coal in grey shales is indicative of a temporary development of marshlands. Both, the lower and upper boundaries of the Hojedk Formation are defined by an erosional unconformity (Fürsich et al. 2009; Wilmsen et al. 2009a).

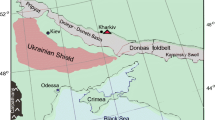
Sketch map showing the location of the exploration well no. 233 in South Kuchak-Ali area, Tabas Block, Central Iran
The Hojedk Formation is known for its rich assemblages of bivalves, ammonites (e.g. Seyed-Emami et al. 2004) and plant macrofossils (e.g. Vassiliev 1984; Poole and Ataabadi 2005; Vaez-Javadi and Mirzaei-Ataabadi 2006; Ameri et al. 2014), which generally indicate a late Bajocian–early Bathonian age.
Palaeogeographical reconstructions indicate that during the Callovian, Northern and Central Iran was located at the northeastern margin of the Neotethys Ocean, approx. 20° to 30° to the north (Thierry 2000; Seyed-Emami et al. 2008; Wilmsen et al. 2009a, 2009c, 2010). Vakhrameev (1991) suggested that during the Late Triassic and Middle Jurassic humidity increased, causing plants to thrive and develop fast and form widespread coal-bearing strata. The Middle Jurassic Hojedk Formation also includes abundant coal seams, with c. 30 beds identified in well no. 233. Many authors have studied the plant micro- (Kimyai 1968; Arjang 1975; Baharadwaj and Kumar 1986) and macrofossils (Barnard 1967; Vassiliev 1984; Schweitzer and Kirchner 1996, 1998; Poole and Ataabadi 2005; Vaez-Javadi and Mirzaei-Ataabadi 2006; Ameri et al. 2014) of the Hojedk Formation, in the Tabas Block. The purpose of the present study is to document the palynological and palaeobotanical content of the Hojedk Formation in the well no. 233, South Kuchak-Ali area, in the Tabas Block and to evaluate its stratigraphic, palaeoecological, palaeoenvironmental and palaeobiogeographical significance.
Material and methods
Forty-eight samples were collected from exploration well no. 233, which is located at N33° 15′ 39″ and E56° 22′ 44″, 60 km southwest of Tabas city in South Kuchak-Ali area, South Khorasan Province, Iran (Figs. 1 and 2). Standard palynological procedures (e.g. Phipps and Playford 1984; Wood et al. 1996) were utilised for extraction and concentration of the palynomorphs. Samples were mechanically crushed into small fragments, 1–2 mm in diameter, and soaked in HCl (30%) for 24 h to dissolve carbonate contents. The silicate content was dissolved using HF (33%). After that, samples were boiled in HCl (10%) for 10 min to prevent silicate gel formation. The organic residue was sieved via a 20 μm mesh. Finally, samples were mixed with zinc chloride (ZnCl2, 2 gr/cm3) and centrifuged to separate palynomorphs from heavy minerals and other unwanted material. Due to their lower specific weight, palynomorphs form a thin layer at the top of all material when centrifuged.

Lithostratigraphic column and distribution of miospore taxa and dinoflagellates through the sampled succession, and proposed palynostratigraphic zone of the Hojedk Formation in the exploration well no. 233, in South Kuchak-Ali area, Tabas Block, Central Iran
This layer was separated and washed through a 20 μm sieve and used to prepare palynological slides. In general, more than 130 palynomorphs were counted per sample. Spores comprise 87.1%, pollen grains 11.5%, bisaccate pollen 1.6% and dinoflagellates 1.6% of the total assemblage (Fig. 3). The proportions of palynomorphs after counting were calculated in percentages for all samples (Table 1). To supplement palaeoecological data derived from the botanical affinities of pollen grains and spores, a palynofacies analysis was performed. To do this, three microscopic fields of view were randomly selected, in which the organic particles were counted. Based on these, selected ratios were calculated, which are presented in Table 1. All rocks samples, residues, and strew slides used in this investigation are permanently housed in the Paleontology Collection at the School of Geology in the University of Tehran, Iran.

Relative abundances (average percentages) of palynomorphs (spores, pollen grains, bisaccate pollen and dinoflagellates) from Hojedk Formation samples
Microfloristic assemblage and palynostratigraphy
Shales and sandstones of the Hojedk Formation in the exploration well no. 233 follow unconformably on limestones of the Badamu Formation. The upper boundary of the Hojedk Formation in the well no. 233 is located at surface level. Most of the studied samples comprise palynological assemblages of fair preservation, including spores, pollen grains and, as minor constituents, dinocysts. The palynoflora includes 13 spore species (assigned to 11 genera) and three pollen species (assigned to three genera) (Fig. 4). Trilete spores dominate in terms of abundance, and among these the fern spore Klukisporites variegatus is of greatest abundance, allowing the identification of the Klukisporites variegatus acme zone (Fig. 2). The palynozonation described by Arjang (1975) for the Hojedk Formation comprises the Klukisporites Zone (Arjang 1975) and the Klukisporites variegatus Subzone (Arjang 1975), which is dated as Middle Jurassic.

Palynomorphs retrieved from the Hojedk Formation. aCyathidites australis Couper, 1953, proximal focus. bDeltoidospora sp., proximal focus. cDictyophyllidites harrisii Couper, 1958, proximal focus. dOsmundacidites wellmanii Couper, 1953, lateral view. eConverucosisporites sp., median focus. fConcavissimisporites punctatus (Delcourt and Sprumont) Brenner, 1963, proximal focus. gAnapiculatisporites sp., proximal focus. h, iKlukisporites scaberis (Cookson and Dettmann) Dettmann, 1963, (h distal focus, i proximal focus). j, kKlukisporites variegatus Couper, 1958, (j distal focus, k proximal focus). lBiretisporites spp., median focus. mVerrucosisporites varians Volkheimer, 1972, proximal focus. nGranulatisporites sp., median focus. oCallialasporites dampieri (Balme) Sukh Dev, 1961, polar view. pCycadopites sp., distal focus. qAraucariacites australis Cookson ex Couper, 1953, median focus. r Indeterminate dinoflagellate cyst, Scale bar 20 μm
Other co-occurring spores represent Anapiculatisporites sp., Concavissimisporites punctatus, Converrucosisporites sp., Cyathidites australis, Deltoidospora sp., Dictyophyllidites harrisii, Granulatisporites sp., Klukisporites scaberis, Biretisporites spp., Osmundacidites wellmanii and Verrucosisporites varians and the pollen taxa Araucariacites australis, Callialasporites dampieri and Cycadopites sp. (Fig. 4)
Arjang (1975) states that the palaeoflora of the Kerman Block changed greatly at the beginning of the Middle Jurassic, that is when Klukia exilis (the Klukisporites variegatus parental plant) became abundant in the region (Fig. 5), an interpretation also supported by observations on plant macroremains (e.g. Popa et al. 2012).

Comparison of the palynostratigraphic scheme proposed herein for the Middle Jurassic Hojedk Formation with those established from coeval strata from Iran and elsewhere (modified after Dehbozorgi 2014)
Klukisporites variegatus is reported from the Middle Jurassic by many authors (e.g. Arjang 1975; Achilles et al. 1984; Bharadwaj and Kumar 1986; Kimyai 1975, 1977; Vajda 2001; Sabbaghiyan 2009; Hashemi-Yazdi 2015; Dehbozorgi 2014; Shevchuk et al. 2018; Slater et al. 2018). This species is also reported widely from Jurassic-Cretaceous sediments of Australia (e.g. Filatoff 1975; Backhouse 1988; McKellar 1998; Sajjadi and Playford 2002). Thus, due to the high abundance of K. variegatus in the Hojedk Formation observed in this study, the succession can confidently be dated as Middle Jurassic. In addition, the occurrence of Klukisporites scaberis, indicative of Middle Jurassic (McKellar 1998), in the lower parts of the formation, further supports the age determination (samples 8–29). Filatoff and Price (1988) stated that the Bajocian–Bathonian boundary is marked by the presence of Steriatella spp. and of Contignisporites burgeri, but these miospores were not found in the current study. Marine palynomorphs are very rare (1.6%) in the Hojedk Formation and cannot be identified taxonomically due to poor preservation.
Palaeoenvironmental interpretations
Palynofacies
Palynofacies analysis can be used to determine certain palaeoenvironmental aspects and sea level changes (e.g. Tyson 1993, 1995; Zavattieri et al. 2008; Batten and Stead 2005). In this study the palaeoenvironment is interpreted based on AOM ratio and phytoclasts (Fig. 6).

Proxies used for palaeoenvironmental interpretation of the Hojedk Formation in well no. 233. a The ratio of PH/AOM/MP shows that PH is more than 70% throughout the formation. b The ratio of AOM(T)/AOM(O) indicates a high oxygen content and low sedimentation rate. c The ratio of P1/ P2 indicates that the sediments were deposited in a shallow nearshore environment. d The ratio of BP/OP shows a high level of oxygen in the depositional environment. PH phytoclasts, AOM amorphous organic matter, MP marine palynomorph, AOM(T) translucent amorphous organic matter, AOM(O) opaque amorphous organic matter, P1 dark equidimensional phytoclasts, P2 dark elongated phytoclasts, BP brown palynomorphs, OP opaque palynomorphs
AOM and ratio of brown to opaque AOM (BP/OP)
The relative amount of transparent AOM and opaque AOM can be used to determine oxygen content. Oxygen is one of the most important factors in environmental interpretation (Bombardiere and Gorin 2000). Transparent AOM is influenced by anaerobic bacteria which are active slightly below the sediment surface, and is formed in non- to low-oxygen environments. These bacteria decompose organic matters, producing nitrate and sulphate (e.g. van Waveren and Visscher 1994; Tyson 1993). High sedimentation rates do not allow these anaerobic bacteria to decay organic matter (Zonneveld et al. 1997), therefore, in this situation, the proportion of transparent AOM is higher than of opaque AOM. The ratio of transparent AOM to opaque AOM is less than one for the sediments in the present study, indicating that the oxygen content was high and sedimentation rate was low (Batten 1996; Oboh-Ikuenobe 1996; Bombardiere and Gorin 2000) (Fig. 6).
The ratio of brown to opaque macerals is another useful proxy for palaeoenvironmental interpretations (Fig. 6 and Table 2). For the material studied in the present study, this ratio is less than one, confirming that during sedimentation, the oxygen content was relatively high in the depositional environment (e.g. van Waveren and Visscher 1994; Bombardiere and Gorin 2000).
Phytoclast ratio (P1/P2)
Phytoclasts originate from plants and are divided into opaque and brown clasts. Higher abundance of phytoclasts indicates that sedimentation occurred in a proximal facies (e.g. van Waveren and Visscher 1994; Schiøler et al. 2002) (Fig. 6). Also the ratio of dark, equidimensional phytoclasts (P1) to elongated phytoclasts (P2) can be used as additional evidence that the sediments of the Hojedk Formation were deposited in a proximal condition (Fig. 6).
Palaeoecology and palaeoclimatic interpretations
Plant macrofossils are useful proxies in palaeogeographical and palaeoecological studies. By determining the parental plants of miospores, it is possible to provide palaeoecological interpretations based on palynomorphs (Table 3).
A major component of the palynomorphs recovered from the Hojedk Formation is miospores. Most miospores recognised in this study are related to ferns indicating that this group was a dominant component of the source vegetation (Figs. 7 and 8; Table 3). A small number of miospores is related to conifers and ginkgophytes and some to bryophytes (Figs. 7 and 8). Based on the identified parental plants and their inferred palaeoecology and also the presence of coal-bearing layers with woody debris, it can be concluded that the Hojedk Formation was deposited under a relatively warm and humid climate. The composition of the macrofossils (including remains of 22 taxa) in the studied deposits is in agreement with previous interpretations based on plant macroremains occurring in this formation at other localities (e.g. Ameri et al. 2014) (Figs. 9 and 10).

The abundance per sample of the miospore species observed in the Hojedk Formation (well no. 233)

Schematic diagram showing the relative frequency of the parent plants of the miospores identified in this study

Plant macroremains from well no. 233 in southern Kuchak-Ali area (Tabas block). aAnnulariopsis simpsoni (Phillips) Harris, 1946. bEquisetites lateralis Phillips, 1829 emend. Gould, 1968. c Diaphragm of Equisetites lateralis Phillips emend. Gould, 1968 (= Equisetum laterale Phillips 1829). dSpiropteris sp. eDictyophyllum sp. f Sterile frond of Klukia westii Jacob and Shukla, 1955. g Fertile frond of Klukia exilis (Phillips) Raciborski, 1890, h Sterile frond of Klukia exilis (Phillips) Raciborski, 1890. iMarattiopsis munsteri (Geoppert) Shimper, 1869. j.Cladophlebis denticulata (Brongniart) Fontaine, 1889. kFerizianopteris undulata Fakhr, 1975. l Sterile and fertile frond of Coniopteris hymenophylloides (Brongniart) Seward, 1900 emend. Harris, 1961. mSagenopteris iranica Fakhr, 1975. nSagenopteris colpodes Harris, 1940

Plant macroremains from well no. 233 in southern Kuchak-Ali area (Tabas Block). a Fertile pinnules of Todites williamsoni (Brongniart) Seward, 1900. b Sterile pinnules of Todites williamsoni (Brongniart) Seward, 1900. c 1.Pterophyllum schenki Zeiller, 1903, 2.Anomozamites thomasi Harris, 1969. dPtilophyllum pectinoides (Phillips) emend. Harris, 1969. ePterophyllum sp. fNilssonia bargi-bidi Barnard and Miller, 1976. g.Nilssonia herriesi (Harris) Schweitzer et al. 2000. hNilssonia sp. i Male cone of Elatides thomasii Harris, 1979. jPodozamites distans (Presl 1838) Braun, 1843
The coals in the sediments of the Hojedk Formation are derived from carbonised plant macrofossils, which are indicative of a moist nearshore environment (e.g. Ameri et al. 2014). In addition, higher amounts of spores in comparison to pollen grains can be seen as further evidence for a nearshore source (e.g. Tyson 1993, 1995; Zavattieri et al. 2008). Additionally, the higher amount of terrestrial palynomorphs compared to marine palynomorphs indicates that the palaeoenvironment of the succession was a deltaic system (Tyson 1987; Van der Zwan 1990; Zavattieri et al. 2008). The presence of dinocysts in two samples (41 and 42) (Table 2) may point to a temporary sea level rise during the deposition of the formation (Fig. 6).
Palaeobiogeographical inferences
One of the most important events in the Jurassic of Iran is the Cimmerian Orogeny (Wilmsen et al. 2009a, 2009c). This event led to the closure of the Palaeotethys Ocean and uplifting as well as faulting in the sedimentary basin (Stocklin 1968). However, the folding also caused a great amount of sediments to be deposited during the Bajocian–Bathonian in central Iran (Berberian and King 1981; Wilmsen et al. 2009a). During the Permian, the Iranian microcontinent was separated from Gondwana and moved northward to eventually collide with Eurasia during the Triassic (Berberian and King 1981; Alavi et al. 1997; Stampfli and Borel 2002; Wilmsen et al. 2009b). During the Norian-Rhaetian, the subduction of the Neotethys Ocean under the Iranian Plate gave rise to the extensive volcanic area known as Sanandaj-Sirjan Zone (Arvin et al. 2007; Wilmsen et al. 2009b). The deposits from this time are formally named as Nayband Formation (Fürsich et al. 2005). The Cimmerian event caused a slab break-off, pushing the plate northward and giving rise to the Alborz Mountain Range, similar to the slab break-off, which gave rise to the North Alpine Foreland in the Middle Oligocene (Wilmsen et al. 2009b; Regard et al. 2008).
Two major Jurassic palaeofloral provinces are recognised in Eurasia. One, encompassing the north, is known as Siberian province and the other, encompassing mostly the south of Eurasia, is named Euro-Sinian province (Vakhrameev 1987). Subsequently Vakhrameev (1991) introduced the names Siberian and Indo-European for these Jurassic floristic provinces (Fig. 11). He also divided the Indo-European province into Europe, Mid-Asia (extending south from the Karagandar-Turgay region in the north to the coast of the Jurassic sea in the south), and Western Asia. This study follows the division of Vakhrameev (1991) utilising the term Indo-European province. The Middle Jurassic flora includes a variety of lycopsids, horsetails, ferns and gymnosperms (Gomolitzky and Khundayberdyev 1976). Palaeobotanical evidence reported from Georgia-Transcaucasia in Mid-Asia shows a comparable variety and congruence with the European province during the Middle Jurassic (Barale et al. 1991; Vakhrameev 1991). Previous palaeobotanical reports from the Middle Jurassic of the Kerman Block (e.g. Schweitzer and Kirchner 1996; Vaez-Javadi and Mirzaei-Ataabadi 2006) demonstrate that approximately 50% of the palaeoflora were similar to that of Eurasia (Delle 1967), the western Ural in Russia (Genkina 1963) and the Saighan in northern Afghanistan (Jacob and Shukla 1955), all of which are parts of the Indo-European province (Vakhrameev 1991).

The palaeofloristic divisions of the Early–Middle Jurassic showing the Central Iran Block as part of the Indo-European and Mid-Asian provinces, modified after Vakhrameev (1991)
Records of permineralized wood belonging to Prototaxoxylon feriziensis (Fakhr and Marguerier 1977) and Xenoxylon cf. latiporosum (Cramer) Gothan 1905 (Nadjafi 1982; Poole and Ataabadi 2005) in sediments of Early Jurassic and early Middle Jurassic age provide evidence that trees were part of the vegetation in Iran during this time. On a global scale Xenoxylon seems to be restricted to relatively high latitudes (> 35–40° N) in the Northern hemisphere during the Middle Jurassic (Philippe et al. 2017). The similarity between the palaeoflora of northeastern Iran and northern Afghanistan indicates a common terrestrial distribution area (such as an island) at the southern margin of Eurasia (Seyed-Emami 1971; Fakhr 1975; Vakhrameev 1991). However, Vozenin-Serra and Taugourdeau (1985) argue that the Kerman Block in Iran was part of Gondwana during this period and that there was a large body of water between this block and India.
The genus Coniopteris was abundant in the Mid-Asian province, whereas, Thaumatopteris and Dictyophyllum, which were abundant in Europe (Shevchuk et al. 2018), have only rarely been found in the Mid-Asian province. In the Middle Jurassic, the genus Cladophlebis was rare in this province, while Klukia was abundant. Cycadophytes such as Otozamites, Nilssonia and Ptilophyllum are notable in the Mid-Asian province (Vakhrameev 1991). Jacob and Shukla (1955) argued that Annulariopsis, Dictyophyllum and Clathropteris would only occur in the Northern Hemisphere and not in Gondwana. However, in the meantime these genera have also been reported from Argentina (Frenguelli 1941; Herbst 1966; Choo et al. 2016) and Antarctica (Bomfleur and Kerp 2010).
The following plant macrofossil taxa could be identified in the Hojedk Formation: Annulariopsis, Equisetites, Marattiopsis, Cladophlebis, Clathropteris, Dictyophyllum, Klukia, Ferizianopteris, Coniopteris, Phlebopteris, Sagenopteris, Pterophyllum, Anomozamites, Ptilophyllum, Nilssonia, Podozamites, Elatides and Elatocladus (Figs. 9 and 10). The abundance of Coniopteris, Ptilophyllum, Nilssonia and Klukia, which are indicators for the Mid-Asian province, together with the high variety of cycadophytes, indicates that the sediments of the Hojedk Formation in Central Iran were deposited in the Indo-European floristic province during the Middle Jurassic (Fig. 11).
Regional correlation
The Middle Jurassic macroflora of the Hojedk Formation in the Central Iran has first been studied by Barnard (1967). This early work was continued by a series of contributions by Vassiliev (1984), Schweitzer and Kirchner (1996, 1998), Poole and Mirzaie Ataabadi (2005) and Vaez-Javadi and Mirzaei-Ataabadi (2006) which contributed to the knowledge of the Middle Jurassic flora of this formation. In addition, the palynoflora from the Middle Jurassic of the Hojedk Formation has so far only been studied in the Kerman Block (Kimyai 1968; Arjang 1975; Bhardwaj and Kumar 1986; Hashemi-Yazdi et al. 2014; Hashemi-Yazdi 2015).
Kimyai (1968) reported a palynoflora from the upper–Middle Jurassic in the northwest of the Kerman area, and reported the following genera, which were also found in this study: Araucariacites, Cyathidites, Deltoidospora, Dictyophyllidites, Klukisporites and Osmundacidites.
Achilles et al. (1984) studied the microflora of the Norian–Middle Jurassic recovered from different parts of the Alborz Mountains. They reported the Leptolepidites argenteaeformis Zone (Achilles et al. 1984), which could not be seen in the samples studied here.
In similar reports, some authors positioned the Early–Middle Jurassic boundary at the FOO (first observed occurrence) of Ischyosporites (Klukisporites) variegatus and LOO (last observed occurrence) of Concavisporites (Dictyophyllidites) spp. (e.g. Achilles et al. 1984; Arjang 1975; Ashraf 1977).
Bharadwaj and Kumar (1986) reported three assemblage zones according to the abundance of (A) Dictyophyllidites (late Early Jurassic), (B) Cyathidites (early Middle Jurassic) and (C) Klukisporites (late Middle Jurassic). They argued that the FOO of Klukisporites variegatus is indicative for the late Middle Jurassic. Their palynozones B and C can be correlated with the current study on the Hojedk Formation in well no. 233. Common miospores between the two studies are Araucariacites, Cyathidites, Cycadopites and Dictyophyllidites.
Sajjadi et al. (2007) studied the Kashafrud Formation in the Senjedk section, Kopet Dagh Basin, and reported three assemblage zones (A–C) from the Bajocian–Bathonian. They reported the following microflora, which could also be seen in the current study: Concavissimisporites punctatus, Cyathidites australis, Deltoidospora sp., Dictyophyllidites harrisii, Klukisporites variegatus, Osmundacidites wellmanii, Araucariacites australis and Callialasporites dampieri.
Dehbozorgi et al. (2013) studied the Dalichai and the Baghamshah formations in the east of Semnan. They reported the Klukisporites Zone and the K. variegatus Subzone of Arjang (1975) for these strata, which are indicative for the late Bajocian-Callovian. Moreover, they defined the Callialasporites dampieri Assemblage Zone, which encompasses the Dictyosporites complex Oppel zone (late Bathonian) and the Klukisporites variegatus subzone (late Bathonian-Callovian) for the Baghamshah Formation. The Klukisporites variegatus subzone, introduced for the Baghamshah Formation, can be correlated with the Hojedk Formation in the well no. 233. Among the miospores reported by Dehbozorgi et al. (2013), the following could also be seen in this study: Concavissimisporites punctatus, Concavissimisporites verrucosus, Cyathidites australis, Dictyophyllidites harrisii and Osmundacidites wellmanii. Araucariacites australis, Callialasporites dampieri, and Cycadopites spp.
Hashemi-Yazdi et al. (2014) and Hashemi-Yazdi (2015) studied the Hojedk Formation in the Eshkeli section in the Kerman Block and found the Klukisporites Zone and the K. variegatus Subzone of Arjang (1975), which are indicative of late Bajocian–Bathonian. They reported some miospores among which the following are in common with the current study: Cyathidites australis, Concavissimisporites punctatus, Dictyophyllidites harrisii, Klukisporites scaberis, Limbosporites lundbladii, Osmundacidites wellmanii and Callialasporites dampieri.
Global correlation
Since the late 1950s, many studies were conducted on Jurassic sediments and a number of palynozones have been introduced for this period (cf. Hashemi-Yazdi et al. 2014). In this section, we compare our results with those of previous studies on Jurassic successions in other regions of the world.
Asia
Ashraf (1977) suggested that the Early–Middle Jurassic boundary in Afghanistan is concurrent with the genus Ischyosporites (Klukisporites) and the Middle–Late Jurassic is concurrent with the FOO of the genus Concavisporites (Dictyophyllidites). Furthermore, this author defined the Ischyosporites (Klukisporites) variegatus – Duplexisporites (Striatella) problematicus (seebergensis) palynozone for Middle Jurassic sediments which encompasses very few miospores shared with the present material.
Tripathi (2001) studied the microflora of the Rajmahal Basin in India and described 10 palynozones, reaching from the Early Permian up to the Early Cretaceous. Among all palynozones, only Assemblage 5, which indicates an Aalenian-Bajocian age, is comparable with the current study.
Tripathi (2004) described three assemblage zones for the Rajmahal Basin in India. This author argued that the high abundance of Callialasporites is indicative for the Middle Jurassic, from which only one species could be found in the current study. In addition, he described the palynozone Assemblage 1 for the Middle Jurassic including Concavissimisporites, Klukisporites and Callialasporites, which are present in the current study as well.
Jiang et al. (2008) studied the palynomorphs of the Tarim Basin in Northwest China. Among the miospores recovered by these authors, the following are found also in the present study: Callialasporites dampieri, Cyathidites australis, Dictyophyllidites harrisii, Klukisporites variegatus and Osmundacidites wellmanii.
Ashraf et al. (1999, 2010) studied palynomorphs from the Triassic and Jurassic in the Junggar Basin (NW China) and reported the Ischyosporites (Klukisporites) variegatus–Duplexisporites (Striatella) problematicus (seebergensis) zone, indicative for a Middle Jurassic age, from the uppermost part of the lower Sangonghe Formation, the upper Sangonghe Formation and the Xishanyiao Formation. They stated that these taxa are also known from other regions, like Afghanistan, Iran and Germany and can thus be used for long-range palynostratigraphical correlation. These, and more recent studies from the Junggar Basin (Sha et al. 2011, 2015) show that the miospore assemblage are comparable with those of this study, especially regarding the occurrence of Ischyosporites (Klukisporites) variegatus and Cycadopites sp.
Europe
Stefanowicz (2008) reported five palynozones (A–E) for the microflora of Scotland. Palynozone D, indicating middle Bajocian–early Bathonian, is comparable with the current study. Among the miospores recovered by Stefanowicz (2008), Araucariacites, Ischyosporites (Klukisporites) and Callialasporites were also found in the current study.
Ainsworth and Riley (2010) studied Triassic–Middle Jurassic (Bathonian) successions in Southern England. Among the miospores recovered by these authors, the following are found in the present study: Callialasporites and Deltoidospora.
Rostovtseva (2011) reported two palynozones for the Bathonian (Palynocomplex I) and the Callovian (Palynocomplex II) for the sediments of the Middle Jurassic northwest of Moscow. Based on the common occurrence of miospores such as Cyathidites australis, Klukisporites variegatus and Osmundacidites, the palynozone she described for the Bathonian can be correlated with the sediments of the Hojedk Formation in well no. 233.
Slater et al. (2017) investigated miospores assemblages from Middle Jurassic marine deposits of the Brent Group in the northern North Sea to evaluate vegetation and depositional processes. Among the miospores, only Deltoidospora is a common taxon between their and this study.
Shevchuk et al. (2018) described two assemblages zones for Jurassic (Bathonian) sediments of the Donbas, northeast Ukraine. Twenty-six spores and 19 pollen taxa were identified from assemblage A, which is dominated by the genera Cyathidites and Osmundacidites, and is dated as middle Bathonian. These two taxa are in common with this study. Also, 27 spores and 22 pollen taxa were identified from assemblage B. In this assemblage pollen grains are abundant.
Middle Jurassic spores and pollen from the Ravenscar Group of Northern Yorkshire from dinosaur foot prints were investigated by Slater et al. (2018) in order to reconstruct the vegetation and assess possible dinosaur-plant interactions. Common taxa are Deltoidospora spp., Dictyophyllidites harrisii, Araucariacites australis, Cycadopites spp. and Callialasporites spp.
Australia
Balme (1957) described three assemblage zones for the Jurassic-Early Cretaceous in Australia (I, IIa and IIb respectively). Later on, Balme (1964) modified the described palynozones to the Exesipollenites Assemblage (for the Early–Middle Jurassic), the Dampieri Assemblage Zone (for the Middle–Late Jurassic) and the Microcachryidites Assemblage for the Early Cretaceous. He found that pollen grains are most abundant in the Middle Jurassic, which is in contrast to the current study. Among the miospores reported by Balme (1964), Cyathidites and Araucariacites australis are also present in this study.
Reiser and Williams (1969) defined an assemblage zone for the Surat Basin in Queensland, Australia, which was named Tsugaepollenites (Callialasporites) segmentatus–Tsugaepollenites (Callialasporites) dampieri zone. This palynozone indicates Toarcian-Tithonian. Among all the species reported by these authors, only Callialasporites dampieri was seen in the sediments of the Hojedk Formation in well no. 233.
Filatoff (1975) studied Jurassic sediments in the west of the Perth Basin in Western Australia. This author reported two palynozones indicating Hettangian-Bajocian and Bajocian-Kimmeridgian [i.e. Exesipollenites tumulus Assemblage Zone (Filatoff 1975) and Callialasporites dampieri Assemblage Zone (Filatoff 1975) respectively]. The following miospores are shared between this report and the present study: Converrucosisporites variverrucatus (verrucosus), Klukisporites neovariegatus (variegatus), Klukisporites scaberis and Callialasporites dampieri.
Helby et al. (1987) introduced two superzones in Australia: the Callialasporites dampieri superzone indicating Hettangian-Kimmeridgian and the Microcachryidites superzone indicating Tithonian-Albian. There is one species, Callialasporites dampieri, which is shared between this study and the current study on the Hojedk Formation.
McKellar (1998) studied Middle Jurassic miospores in the west of Australia and reported eight palynozones ranging from Hettangian to Tithonian. Due to the lack of the key species Aequitriradites norrisii in the present study, his reported biozone (Retitriletes circolumenus Association Zone) which indicates Bajocian–early Bathonian, could only partially be correlated with the Hojedk palynoflora. However, Klukisporites scaberis, Klukisporites variegatus and Callialasporites spp. could also be found in this study.
Mantle and Riding (2012) studied the palynomorphs of the Middle Jurassic in multiple basins in Australia and reported some miospores among which the following could be found in this study as well: Concavissimisporites, Cyathidites, Dictyophyllidites, Klukisporites, Osmundacidites and Araucariacites.
Argentina
Quattrocchio et al. (2001) studied the miospores of the Middle Jurassic in the Guyo Group (Lajas and Callaco formations) from Argentina. Based on the presence of Deltoidospora, Biretisporites, Osmundacidites, Dictyophyllidites, Klukisporites, Araucariacites, Callialasporites and Cycadopites, their study can be correlated with this study.
Volkheimer et al. (2008) studied the palynoflora recovered from the North of Argentina and reported some miospores from the Triassic–Middle Jurassic, among which Araucariacites, Callialasporites and Dictyophyllidites, were also found in the present study.
Stukins et al. (2013) investigated the Middle Jurassic of the Lajas Formation, Neuquen Basin in Argentina. On the basis of the occurrence of the common taxa Deltoidospora, Araucariacites australis and Callialasporites spp. their study could only partially be correlated with the Hojedk Formation palynoflora.
Other localities
Ibrahim et al. (2001) studied Jurassic sediments in the Eastern Desert of Egypt. They introduced the Converrucosisporites spp., Verrucosisporites spp., Trilobosporites spp. assemblage zone (Ibrahim et al. 2001) indicating a Bajocian-Callovian age. This palynozone starts with Classopollis (Corollina) torosa, Concavissimisporites variverrucatus (verrucosus), Dictyophyllidites harrisii, Camarozonosporites sp. (ramosus), Verrucosisporites varians and Klukisporites variegatus and ends at the FOO of Cicatrocosisporites spp. Within this palynozone Concavissimisporites, Converrucosisporites, Klukisporites and Verrucosisporites are most abundant. Except the three genera Trilobosporites, Cicatricosisporites and Classopollis none of the miospores mentioned above have been found in the current study.
Ahmad et al. (2014) defined a Middle Jurassic (Bathonian) Callialasporites dampieri–Murospora florida assemblage for the Hamam Formation in the western part of the Wadi Zarqa region in northwestern Jordan. This assemblage is characterised by the occurrence of Callialasporites dampieri, Callialasporites trilobatus, Murospora florida, Corollina torosa, Osmundacidites wellmanii, Granulatisporites jurassicus, Cyathidites australis, Cyathidites minor, Protopinus scanicus, Klukisporites variegatus, as well as a number of other taxa diagnostic for a Jurassic age. Callialasporites dampieri, Osmundacidites wellmanii, Cyathidites australis and Klukisporites variegatus are also common miospores in the current study.
Conclusions
Thirteen spore species belonging to 11 genera and three pollen species belonging to three genera were recognised from the Hojedk Formation in the well no. 233 located in the Kuchak-Ali area, south of Tabas Block, Iran. Due to the high abundance of this spore type, a Klukisporites variegatus acme zone is introduced. According to the biostratigraphic age of relevant miospores (i.e. Klukisporites scaberis and Klukisporites variegatus), the formation is dated as Middle Jurassic, which is in agreement with data derived from plant macrofossils. Terrestrial palynomorphs are more abundant, when compared to the marine palynomorphs, pointing to a shallow nearshore environment. Palynological data gained from the formation show that (A) equidimensional phytoclasts are dominant, (B) AOM is the main contributor to organic matter and the ratio of transparent AOM/dark AOM is more than one and (C) the ratio of BP/OP is less than one throughout the whole section. These data indicate that the Hojedk Formation was probably deposited in an oxygenated deltaic system in which the sedimentation rate was low and the preservation of organic matter was poor. In addition, the presence of marine palynomorphs in very short intervals indicates that probably the sea level transgressed temporarily during the deposition of the formation. Parental plants of miospores are ferns, ginkgophytes, conifers, and bryophytes, which are usually regarded as representatives of a relatively warm and humid climate. Also, the correlation of the parental plants with previous palaeofloristic reports shows that the formation was deposited in the Mid-Asian part of the surrounding Indo-European province of the Northern Hemisphere.
References
Abbink, O. A. (1998). Palynological investigations in the Jurassic of the North Sea region. Laboratory of Palaeobotany and Palynology, Contributions Series, 8, 192.
Achilles, H., Kaiser, H., & Schweitzer, H. J. (1984). Die räto-jurassischen Floren des Iran und Afghanistans: 7. Die Mikroflora der obertriadisch-jurassischen Ablagerungen des Alborz-Gebirges (Nord-Iran). Palaeontographica Abteilung B, 14–95.
Aghanabati, A. (1977). Etude géologique de la région de Kalmard (W. Tabas). Stratigraphie et tectonique. Geological Survey of Iran, Report, 35.
Aghanabati, A. (1998). Jurassic stratigraphy of Iran, volumes 1 and 2. Tehran: Geological Survey of Iran. [in Persian].
Ahmad, F., Farouk, S., & Ziko, A. (2014). Facies associations of the Bathonian Hamam Formation from northwestern Jordan. Arabian Journal of Geosciences, 7(11), 4861–4875.
Ainsworth, N. R., & Riley, L. A. (2010). Triassic to Middle Jurassic stratigraphy of the Kerr McGee 97/12-1 exploration well, offshore southern England. Marine and Petroleum Geology, 27, 853–894.
Alavi, M., Vaziri, H., Seyed-Emami, K., & Lasemi, Y. (1997). The Triassic and associated rocks of the Nakhlak and Aghdarband areas in central and northeastern Iran as remnants of the southern Turanian active continental margin. Geological Society of America Bulletin, 109(12), 1563–1575.
Ameri, H., Dastanpour, M., Khalilizade, H., & Zamani, F. (2014). Plant fossil remains from the Bajocian–Bathonian of Hojedk Formation, Babhutk area, Kerman, Iran. Arabian Journal of Geosciences, 7(6), 2293–2302.
Arjang, B. (1975). Die räto-jurassischen Floren des Iran und Afghanistans. 1. Die Mikroflora der räto-jurassischen Ablagerungen des Kermaner Beckens (Zentral-Iran). Palaeontographica Abteilung B, 85–148.
Arvin, M., Pan, Y., Dargahi, S., Malekizadeh, A., & Babaei, A. (2007). Petrochemistry of the Siah-Kuh granitoid stock southwest of Kerman, Iran: implications for initiation of Neotethys subduction. Journal of Asian Earth Sciences, 30(3–4), 474–489.
Ashraf, A. R. (1977). Die räto-jurassischen Floren des Iran und Afghanistans. 3. Die Mikrofloren der rätischen bis unterkretazischen Ablagerungen Nordafghanistans. Palaeontographica Abteilung B, 1–97.
Ashraf, A. R., Sun, G., Wang, X., Uhl, D., Li, C., & Mosbrugger, V. (1999). The Triassic-Jurassic boundary in the Junggar basin (NW-China)-preliminary palynostratigraphic results. Acta Palaeobotanica. Supplementum, 2, 85–91.
Ashraf, A. R., Sun, Y., Sun, G., Uhl, D., Mosbrugger, V., Li, J., & Herrmann, M. (2010). Triassic and Jurassic palaeoclimate development in the Junggar Basin, Xinjiang, Northwest China—a review and additional lithological data. In T. Martin, G. Sun & V. Mosbrugger (Eds.), Triassic-Jurassic biodiversity, ecosystems, and climate in the Junggar Basin, Xinjiang, Northwest China. Palaeobiodiversity and Palaeoenvironments, 90(3), 187–201.
Backhouse, J. (1988). Late Jurassic and Early Cretaceous palynology of the Perth Basin, Western Australia (Vol. 135). State Print. Division.
Balme, B. E. (1957). Spores and pollen grains from the Mesozoic of Western Australia. Commonwealth Scientific and Industrial Research Organization, Coal Research Section, 25, 1–48.
Balme, B. E. (1964). The palynological record of Australian pre-Tertiary floras. B Geology Department, University of Western Australia.
Balme, B. E. (1995). Fossil in situ spores and pollen grains: an annotated catalogue. Review of Palaeobotany and Palynology, 87(2–4), 81–323.
Barale, G., Iacobidze, E., Lebanidze, Z., & Philippe, M. (1991). Podocarpoxylon svanidzei n. sp. (Coniferae) du Bathonien de Tkibouli en Géorgie occidentale (URSS). Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, 1991, 443–457.
Barnard, P. D. (1967). Flora of the Shemshak Formation. Part 1. Liassic plants from Dorud. Rivista Italiana di Paleontologia e Stratigrafia, 71, 1123–1168.
Batten, D. J. (1996). Upper Jurassic and Cretaceous miospores. Chapter 26A, Palynofacies and palaeoenvironmental interpretation. In J. Jansonius & D. C. McGregor (Eds.), Palynology: Principles and Application (vol. 3, pp. 1011–1064). American Association Stratigraphic Palynologists Foundation.
Batten, D. J., & Stead, D. T. (2005). Palynofacies analysis and its stratigraphic application. In E. A. M. Koutsoukos (Ed.), Applied stratigraphy (pp. 203–226). Dordrecht: Springer.
Berberian, M., & King, G. C. P. (1981). Towards a paleogeography and tectonic evolution of Iran. Canadian Journal of Earth Sciences, 18(2), 210–265.
Bharadwaj, D. C., & Kumar, P. (1986). Palynology of Jurassic sediments from Iran. I: Kerman area. Biological Memoirs, 12(2), 146–172.
Bombardiere, L., & Gorin, G. E. (2000). Stratigraphical and lateral distribution of sedimentary organic matter in Upper Jurassic carbonates of SE France. Sedimentary Geology, 132(3–4), 177–203.
Bomfleur, B., & Kerp, H. (2010). The first record of the dipterid fern leaf Clathropteris Brongniart from Antarctica and its relation to Polyphacelus stormensis Yao, Taylor et Taylor nov. emend. Review of Palaeobotany and Palynology, 160(3–4), 143–153.
Choo, T., Escapa, I., & Benjamin, B. (2016). Monotypic colonies of Clathropteris meniscioides (Dipteridaceae) from the Early Jurassic of central Patagonia, Argentina: implications for taxonomy and palaeoecology. Palaeontographica. Abteilung B, Palaophytologie, 294, 85–109.
Couper, R. A. (1953). Upper Mesozoic and Cainozoic spores and pollen grains from New Zealand (Vol. 22). Alexander Doweld.
Couper, R. A. (1958). British Mesozoic microspores and pollen grains, a systematic and stratigraphic study. Palaeontographica, Abteilung B, 103, 75–179.
Dehbozorgi A. (2014). Palynology and palaeoecology of the Middle Jurassic (Dalichai and Baghamshah formations) east of Semnan (Jam area), PhD thesis. University of Tehran, Iran (unpublished).
Dehbozorgi, A., Sajjadi, F., & Hashemi, H. (2013). Middle Jurassic palynomorphs of the Dalichai Formation, central Alborz Ranges, northeastern Iran: Paleoecological inferences. Science China Earth Sciences, 56(12), 2107–2115.
Delle, G. V. (1967). The Middle Jurassic flora of the Tkvarchelian Coal-Basin (Transcaucasia). Paleobotanika, 6, 53–132. [in Russian with English abstract].
Dettmann, M. E. (1963). Upper Mesozoic microfloras from southeastern Australia. Proceedings of the Royal Society of Victoria, 77, 1–148.
Fakhr, M.S., (1975). Contribution a l'étude de la flore Rhéto–Liasique de la formation de Shemshak de l'Elbourz (Iran). These Uni, Pierre et Marie Curie Paris 6; Publication du Laboratoire de Paleobotanique de l’Universite Paris 6, no.2; 421pp.
Fakhr, M., & Marguerier, J. (1977). Prototaxoxylon feriziense nov. sp. bois fossile du Jurassique moyen de Lírán. Contribution alétude de la flore Rhèto-Liasique de la Formation Shemshak de LElbourz (Iran). Sciences naturelles, University of Paris.
Filatoff, J. (1975). Jurassic palynology of the Perth basin, Western Australia. Palaeontographica Abteilung B, 1–113.
Filatoff, J., & Price, P. L. (1988). A pteridacean spore lineage in the Australian Mesozoic. In P. A. Jell & G. Playford (Eds.), Palynological and palaeobotanical studies in honour of Basil E. Balme. Memoirs of the Association of Australasian Palaeontologists, 5, 89–124.
Frenguelli, J. (1941). Las Camptopterídeas del Lias de Piedra Pintada en el Neuquén (Patagonia). Not. Mus. La Plata, 7. Paléo, 27, 27–57.
Fürsich, F. T., Wilmsen, M., Seyed-Emami, K., Schairer, G., & Majidifard, M. R. (2003). Platform-basin transect of a Middle to Late Jurassic large-scale carbonate platform system (Shotori Mountains, Tabas area, east-Central Iran). Facies, 48(1), 171–198.
Fürsich, F. T., Hautmann, M., Senowbari- Daryan, B., & Seyed-Emami, K. (2005). The Upper Triassic Nayband and Darkuh Formations of east-central Iran: stratigraphy, facies patterns and biota of extensional basins on an accreted terrane. Beringeria, 35, 53–133.
Fürsich, F. T., Wilmsen, M., Seyed-Emami, K., & Majidifard, M. R. (2009). The Mid-Cimmerian tectonic event (Bajocian) in the Alborz Mountains, Northern Iran: evidence of the break-up unconformity of the South Caspian Basin. Geological Society, London, Special Publications, 312(1), 189–203.
Genkina, R. Z. (1963). Fossil flora of the Mesozoic coal-bearing deposits of the East Ural-field of Orsk Coal Basin. 115 pp., 51 pis. Inst. Geol. Exploit. Foss. Fuel, Academy of Sciences USSR, Nauka, Moscow. [in Russian].
Gomolitzky, N. P., & Khudayberdyev, R. H. (1976). About Middle Asian Jurassic flora. The Palaeobotanist, 25, 104–108.
Gothan, W. (1905). Zur Anatomie lebender und fossiler Gymnospermen-Hölzer. Abhandlungen der Königlich Preußischen Geologischen Landesanstalt und Bergakademie, 44, 1–108.
Hashemi-Yazdi, F. (2015). Palynology and palaeoecology of the Dalichai Formation in central - eastern Alborz Basin and the Hojedk Formation at the east -central Iran. PhD thesis. University of Tehran, Iran (unpublished).
Hashemi-Yazdi, F., Sajjadi, F., & Hashemi, H. (2014). Miospores – based palynostratigraphy of the Hojedk Formation, Eshkeli, north of Kerman. Paleontology, 2(1), 111–127.
Helby, R., Morgan, R., & Partridge, A. D. (1987). A palynological zonation of the Australian Mesozoic. In P. A. Jell (Ed.), Studies in Australian Mesozoic palynology. Association of Australasian Palaeontologists, Memoir, 4, 1–94.
Herbst, R. (1966). Revisión de la flora Liásica de Piedra Pintada, provincia de Neuquén, Argentina. Rev. Mus. La Plata (N.S.). Sección Paleontologia, 5, 27–53.
Ibrahim, M. I. A., Aboul Ela, N. M., & Kholeif, S. E. (2001). Palynostratigraphy of Jurassic to Lower Cretaceous sequences from the Eastern Desert of Egypt. Journal of African Earth Sciences, 32(2), 269–297.
Jacob, K., & Shukla, B. N. (1955). Jurassic plants from the Saighan series of northern Afghanistan and their paleoclimatic and paleogeographic significance. Memoirs of the Geological survey of India. Palaeontologia Indica, 33, 1–64.
Jiang, D.X., Wang, Y.D., Robbins, E.I., Wei, J. & Tian, N. (2008). Mesozoic non-marine petroleum source rocks determined by palynomorphs in the Tarim Basin, Xinjiang, Northwestern China. Geological Magazine, 145 (6), 868–885.
Kimyai, A. (1968). Jurassic plant microfossils from the Kerman region. Bulletin of the Iranian Petroleum Institute, 33, 91–111.
Kimyai, A. (1975). Jurassic palynological assemblages from the Shahrud region, Iran. Geoscience and Man, 11, 117–121.
Kimyai, A. (1977). Further information on the palynological stratigraphy of the Mesozoic coaly sediments from Kerman, Iran. Iranian petroleum institute, Proceeding of the 2nd Geological Symposium of Iran, Tehran: 191–217. (in Persian).
Mantle, D. J., & Riding, J. B. (2012). Palynology of the Middle Jurassic (Bajocian-Bathonian) Wanaea verrucosa dinoflagellate cyst zone of the North West Shelf of Australia. Review of Palaeobotany and Palynology, 180, 41–78.
McKellar, J. L. (1998). Late Early to Late Jurassic palynology, biostratigraphy and palaeogeography of the Roma Shelf area, northwestern Surat Basin, Queensland, Australia: including phytogeographic-palaeoclimatic implications of the Callialasporites dampieri and Microcachryidites microfloras in the Jurassic-Early Cretaceous of Australia, based on an overview assessed against a background of floral change and apparent true polar wander in the preceding late Palaeozoic-early Mesozoic. PhD thesis, School of Physical Sciences, The University of Queensland.
Nadjafi, A. A. (1982). Contribution à la connaissance de la flore ligneuse du Jurassique d'Iran (Doctoral dissertation). (unpublished).
Oboh-Ikuenobe, F. E. (1996). Correlating palynofacies assemblages with sequence stratigraphy in Upper Cretaceous (Campanian) sedimentary rocks of the Book Cliffs, east-Central Utah. Geological Society of America Bulletin, 108(10), 1275–1294.
Philippe, M., Puijalon, S., Suan, G., Mousset, S., Thévenard, F., & Mattioli, E. (2017). The palaeolatitudinal distribution of fossil wood genera as a proxy for European Jurassic terrestrial climate. Palaeogeography, Palaeoclimatology, Palaeoecology, 466, 373–381.
Phipps, D., & Playford, G. (1984). Laboratory techniques for extraction of palynomorphs from sediments. Papers of the Department of Geology, University of Queensland, 11(1), 1–23.
Poole, I., & Ataabadi, M. M. (2005). Conifer woods of the Middle Jurassic Hojedk Formation (Kerman basin) Central Iran. IAWA Journal, 26(4), 489–505.
Popa, M. E., Javidan, M., & Falahatgar, M. (2012). Klukia exilis (Philips 1829) Raciborski 1890 emend. Harris 1961 from Ahan Sar, Shemshak group, Iran. Acta Palaeontologica Romaniae, 8, 33–41.
Potonié, R. (1962). Synopsis der Sporae in situ. Die Sporen der fossilen Fruktifikationen (Tallophyta bis Gymnospermophyta) im natürlichen System und im Vergleich mit den Sporae dispersae. Geologisches Jahrbuch, Beihefte, 52, 1–204.
Potonié, R. (1967). New phylogenetic facts on fossil spores. Review of Palaeobotany and Palynology, 1(1–4), 75–82.
Quattrocchio, M. E., García, V., Martínez, M., & Zavala, C. (2001). A hypothetic scenario for the Middle Jurassic in the southern part of the Neuquén Basin, Argentina. 7° International symposium on Mesozoic terrestrial ecosystems, Asociación Paleontológica Argentina. Publicación Especial, 7, 163–166.
Reiser, R. F., & Williams, A. J. (1969). Palynology of the Lower Jurassic sediments of the northern Surat Basin, Queensland. SG Reid, Government Printer.
Regard, V., Faccenna, C., Bellier, O., & Martinod, J. (2008). Laboratory experiments of slab break-off and slab dip reversal: insight into the Alpine Oligocene reorganization. Terra Nova, 20(4), 267–273.
Rostovtseva, J. I. (2011). New palynological data about Middle Jurassic sediments in Northwest Moscow. Moscow University Geology Bulletin, 66(5), 348–353.
Sabbaghiyan H. (2009). Palynostratigraphy of the Dalichai Formation at Kuhe Rahband, south of Mahdishahr, Central Alborz, Iran. MSc. Thesis. University of Tehran, Iran (unpublished).
Sajjadi, F., & Playford, G. (2002). Systematic and stratigraphic palynology of Late Jurassic - earliest Cretaceous strata of the Eromanga Basin, Queensland, Australia, part 2. Palaeontographica, Abteilung A, 261(4–6), 1–99.
Sajjadi, F., Hashemi, H., & Dehbozorgi, A. (2007). Middle Jurassic palynomorphs of the Kashafrud formation, Koppeh Dagh Basin, Northeastern Iran. Micropaleontology, 53(5), 391–408.
Schiøler, P., Crampton, J. S., & Laird, M. G. (2002). Palynofacies and sea-level changes in the middle Coniacian–late Campanian (Late Cretaceous) of the East Coast Basin, New Zealand. Palaeogeography, Palaeoclimatology, Palaeoecology, 188(3–4), 101–125.
Schweitzer, H. J., & Kirchner, M. (1996). Die rhäto-jurassischen Floren des Iran und Afghanistans: 9. Coniferophyta. Palaeontographica, Abteilung B, 238(4–6), 77–139.
Schweitzer, H. J., & Kirchner, M. (1998). Die rhäto-jurassischen Floren des Iran und Afghanistans.11. Pteridospermophyta und Cycadophyta I. Cycadales. Palaeontographica, Abteilung B, 248(1–3), 1–85.
Seyed-Emami, K. (1971). The Jurassic Badamu Formation in the Kerman region, with some remarks on the Jurassic stratigraphy of Iran. Geological Survey of Iran, Report, 19, 1–80.
Seyed-Emami, K., Fürsich, F. T., & Wilmsen, M. (2004). Documentation and significance of tectonic events in the northern Tabas Block (east-central Iran) during the Middle and Late Jurassic. Rivista Italiana di Paleontologia e Stratigrafia, 110(1), 163–171.
Seyed-Emami, K., Fürisch, F. T., Wilmsen, M., Majidifard, M. R., & Skekarifard, A. (2008). Lower and Middle Jurassic ammonoids of the Shemshak Group in Alborz, Iran and their palaeobiogeographical and biostratigraphical importance. Acta Palaeontologica Polonica, 53(2), 237–260.
Sha, J., Olsen, P. E., Yanhong Pan, Y., Xu, D., Wang, Y., Zhang, X., & Vajda, V. (2015). Triassic-Jurassic climate in continental high-latitude Asia was dominated by obliquity-paced variations (Junggar Basin, Urumqi, China). PNAS, 112, 3624–3629.
Sha, J., Vajda, V., Pan, Y., Larsson, L., Wang, Y., Cheng, X. J., Deng, S., Yao, X., Chen, S., Zhang, X., & Peng, B. (2011). The stratigraphy of the Triassic−Jurassic boundary successions of the southern margin of Junggar Basin, northwestern China. Acta Geolologia Sinica, 85, 421–436.
Shevchuk, O., Slater, S. M., & Vajda, V. (2018). Palynology of Jurassic (Bathonian) sediments from Donbas, Northeast Ukraine. In S. M. Slater, E. Kustatscher & V. Vajda, (Eds.), Jurassic biodiversity and terrestrial environments. Palaeobiodiversity and Palaeoenvironments, 98(1), 153–164.
Slater, S. M., McKie, T., Vieira, M., Wellman, C. H., & Vajda, V. (2017). Episodic river flooding events revealed by palynological assemblages in Jurassic deposits of the Brent Group, North Sea. Palaeogeography, Palaeoclimatology, Palaeoecology, 485, 389–400.
Slater, S. M., Wellman, C. H., Romano, M. & Vajda, V. (2018). Dinosaur-plant interactions within a Middle Jurassic ecosystem palynology of the Burniston Bay dinosaur footprint locality, Yorkshire, UK. In S. M. Slater, E. Kustatscher & V. Vajda, (Eds.), Jurassic biodiversity and terrestrial environments. Palaeobiodiversity and Palaeoenvironments, 98(1), 139–151.
Stampfli, G. M., & Borel, G. D. (2002). A plate tectonic model for the Paleozoic and Mesozoic constrained by dynamic plate boundaries and restored synthetic oceanic isochrons. Earth and Planetary Science Letters, 196(1), 17–33.
Stefanowicz, S. (2008). Palynostratigraphy and palaeoclimatic analysis of Lower-Middle Jurassic (Pliensbachian-Bathonian) of the Inner Hebrides, NW Scotland (pp. 34). Dissertations in Geology at Lund University.
Stocklin, J. (1968). Structural history and tectonic of Iran: a review. American Association of Petroleum Geologists Bulletin, USA, 52, 1229–1258.
Stocklin, J., & Nabavi, M. H. (1971). Explanatory text of the Boshruyeh Quadrangle map 1: 250,000. Quadr. J, 7, 1–50.
Stukins, S., Jolley, D. W., McIlroy, D., & Hartley, A. J. (2013). Middle Jurassic vegetation dynamics from allochthonous palynological assemblages: an example from a marginal marine depositional setting; Lajas Formation, Neuquén Basin, Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology, 392, 117–127.
Takin, M. (1972). Iranian geology and continental drift in the Middle East. Nature, 235(5334), 147–150.
Thierry, J. (2000). Middle Callovian (157-155 ma). In J. Dercourt, M. Gaetani et al. (Eds.), Atlas Peri - Tethys, Palaeogeographical maps (pp. 1–97). Paris: CCGM/CGMW.
Tripathi, A. (2001). Permian, Jurassic and Early Cretaceous Palynofloral assemblages from subsurface sedimentary rocks in Chuperbhita Coalfield, Rajmahal Basin, India. Review of Palaeobotany and Palynology, 113, 237–259.
Tripathi, A. (2004). Palynology evidences of hitherto unrecognized Jurassic sedimentation in Rajmahal Basin, India. Rìvista Italiana di Paleontologia e Stratigrafia, 110(1), 35–42.
Tyson, R. V. (1987). The genesis and palynofacies characteristics of marine petroleum source rocks. Geological Society, London, Special Publications, 26(1), 47–67.
Tyson, R. V. (1993). Palynofacies analysis. In D. G. Jenkins (Ed.), Applied micropaleontology (pp. 153–172). Dordrecht: Kluwer Academic Publishers.
Tyson, R. V. (1995). Sedimentary organic matter; organic facies and palynofacies (p. 615). London: Chapman and Hall.
Vaez-Javadi, F., & Mirzaei-Ataabadi, M. (2006). Jurassic plant macrofossils from the Hojedk Formation, Kerman area, east-Central Iran. Alcheringa, 30(1), 63–96.
Vakhrameev, V. A. (1987). Climates and the distribution of some gymnosperms in Asia during the Jurassic and Cretaceous. Review of Palaeobotany and Palynology, 51, 205–212.
Vakhrameev, V. A. (1991). Jurassic and Cretaceous floras and climates of the earth (p. 318). Cambridge University Press.
Vajda, V. (2001). Aalenian to Cenomanian terrestrial palynofloras of SW Scania, Sweden. Acta Palaeontologica Polonica, 46(3), 403–426.
Van der Zwan, C. J. (1990). Palynostratigraphy and Palynofacies reconstruction of the Upper Jurassic to lowermost Cretaceous of the Draugen field, offshore mid Norway. Review of Palaeobotany and Palynology, 62, 157–186.
van Waveren, I., & Visscher, H. (1994). Analysis of the composition and selective preservation of organic matter in surficial deep-sea sediments from a high-productivity area (Banda Sea, Indonesia). Palaeogeography, Palaeoclimatology, Palaeoecology, 112(1–2), 85–111.
Vassiliev, Y. (1984). Mesozoic plant fossils from coal areas in Iran. Atlas of the Ministry of Mine and Metal 2(2), 97 pp., 47 pls.
Volkheimer, W., Rauhut, O. W. M., Quattrocchio, M. E., & Martinez, M. A. (2008). Jurassic paleoclimates in Argentina, a review. Revista de la Asociación Geológica Argentina, 63(4), 549–556.
Vozenin-Serra, & Taugourdeau-Lantz, J. (1985). La flore de la formation Shemshak (Rhetien a Bajocien; Iran); rapports avec les flores contemporaines; implications paleogeographiques. Bulletin de la Société Géologique de France, 1(5), 663–678.
Wilmsen, M., Fürsich, F. T., Seyed-Emami, K., & Majidifard, M. R. (2009a). An overview of the lithostratigraphy and facies development of the Jurassic system on the Tabas Block, eastcentral Iran. In M. F. Brunet, M. Wilmsen & J. Granath (Eds.), South Caspian to Central Iran basins. Geological Society, London, Special Publications, 312, 323–344.
Wilmsen, M., Fürsich, F. T., Seyed-Emami, K., Majidifard, M. R., & Taheri, J. (2009b). The Cimmerian orogeny in northern Iran: Tectono-stratigraphic evidence from the foreland. Terra Nova, 21(3), 211–218.
Wilmsen, M., Fürsich, F. T., & Taheri, J. (2009c). The Shemshak Group (Lower–Middle Jurassic) of the Binalud Mountains, NE Iran: stratigraphy, depositional environments and geodynamic implications. Geological Society, London, Special Publications, 312(1), 175–188.
Wilmsen, M., Fürsich, F. T., Seyed-Emami, K., Majidifard, M. R., & Zamani-Pedram, M. (2010). Facies analysis of a large-scale Jurassic shelf-lagoon: the Kamar-e-Mehdi Formation of east-Central Iran. Facies, 56(1), 59–87.
Wood, G. D., Gabriel, A. M., & Lawson, J. C. (1996). Chapter 3: Palynological techniques-processing and microscopy. In J. Jansonius & D. C. McGregor (Eds.), Palynology: Principles and applications. Dallas, American Association of Stratigraphic Palynologists Foundation, 1, 29–50.
Zavattieri, A. M., Rosenfeld, U., & Volkheimer, W. (2008). Palynofacies analysis and sedimentary environment of Early Jurassic coastal sediments at the southern border of the Neuquén Basin, Argentina. Journal of South American Earth Sciences, 25(2), 227–245.
Zonneveld, K. A., Versteegh, G. J., & de Lange, G. J. (1997). Preservation of organic-walled dinoflagellate cysts in different oxygen regimes: a 10,000 year natural experiment. Marine Micropaleontology, 29(3–4), 393–405.
Acknowledgements
We would like to express our thanks to Dr. M. S. Fakhr for his guidance and help in identification of our macroplant fossils. Also, it was an honour for us to have the guidance and consultancy of Dr. F. Hashemi Yazdi for this research. We would like to thank Dr. A. Dehbozorgi and National Iranian Steel Company for their cooperation in the field. This research was conducted under the public funds available to the University of Tehran. In addition, we thank Vivi Vajda and an anonymous reviewer for their comments that helped to improve the manuscript considerably.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Badihagh, M.T., Sajjadi, F., Farmani, T. et al. Middle Jurassic palaeoenvironment and palaeobiogeography of the Tabas Block, Central Iran: palynological and palaeobotanical investigations. Palaeobio Palaeoenv 99, 379–399 (2019). https://doi.org/10.1007/s12549-018-0361-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-018-0361-0